










Services on Demand
Journal
Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.13 no.1 Bogotá Jan./June 2011
ARTÍCULO CORTO
Adaptación de una cepa compatible con Acidithiobacillus ferrooxidans sobre concentrados de calcopirita (CuFeS2), esfalerita (ZnS) y galena (PbS)
Adaptation of a strain Acidithiobacillus ferrooxidans compatible on concentrates of chalcopyrite (CuFeS2), sphalerite (ZnS) and galena (PbS)
E. Mejía 1 , J. D. Ospina 2 , B. L. Osorno 3 , M. A. Márquez 4 , A. L. Morales 5.
1 Escuela de Ingeniería de Materiales, Grupo de Mineralogía Aplicada y Bioprocesos, Universidad Nacional de Colombia, Sede Medellín. ermejiare@unal.edu.co
2 Escuela de Ingeniería de Materiales, Grupo de Mineralogía Aplicada y Bioprocesos, Universidad Nacional de Colombia, Sede Medellín. judospinaco@unal.edu.co
3 Escuela de Ingeniería de Materiales, Grupo de Mineralogía Aplicada y Bioprocesos, Universidad Nacional de Colombia, Sede Medellín. losorno@unal.edu.co
4 Escuela de Ingeniería de Materiales, Grupo de Mineralogía Aplicada y Bioprocesos, Universidad Nacional de Colombia, Sede Medellín. mmarquez@unal.edu.co
5 Grupo de Estado Sólido, Sede de Investigación Universitaria, Universidad de Antioquia, Sede Medellín. amoral@fisica.udea.edu.co
Recibido: julio 30 de 2010 Aprobado: mayo 25 de 2011
Resumen
En este estudio se evaluó la adaptación de una cepa compatible con Acidithiobacillus ferrooxidans a altas densidades de pulpa de calcopirita, esfalerita y galena, con dos distribuciones de tamaño de partícula, -200 y -325 serie Tyler de tamices. Los microorganismos fueron adaptados por la disminución gradual de la fuente principal de energía, sulfato ferroso, y el aumento en el contenido de mineral, para finalmente realizar un subcultivo sin la adición de fuente de energía externa. La realización de subcultivos en serie resultó ser una estrategia eficaz para la adaptación a altas densidades de pulpa de esfalerita, calcopirita y galena indicando que el protocolo empleado es adecuado. Los resultados muestran que la cepa compatible con Acidithiobacillus ferrooxidans es más resistente a altas concentraciones de esfalerita, seguido por calcopirita y finalmente por galena. El tamaño de partícula juega un papel fundamental en la adaptación de los microorganismos al mineral.
Palabras clave: esfalerita, calcopirita, galena, adaptación, Acidithiobacillus ferrooxidans, biolixiviación.
Abstract
In this study the adaptation of Acidithiobacillus ferrooxidans-like to high concentrations of chalcopyrite, sphalerite and galena were evaluated with two mineral-particle sizes: 200 and 325 Tyler mesh. The strain was adapted using two simultaneous processes. The first one consisted in a gradual decreasing of the main energy source, ferrous sulphate. The second one consisted in a gradual increasing of the mineral content. Finally, a test was made without ferrous sulphate. The serial subculturing was found to be an efficient strategy to adapt Acidithiobacillus ferrooxidans-like to higher concentrations of chalcopyrite, sphalerite and galena. This indicates that a suitable protocol was employed. The results showed that Acidithiobacillus ferrooxidans-like is more resistant to high concentration of sphalerite, chalcopyrite and galena in descendant order. The particle size played an important role in the adaption of microorganism to the mineral.
Key words: Sphalerite, chalcopyrite, galena, adapting, Acidithiobacillus ferrooxidans, bioleaching.
Introducción
La biolixiviación de sulfuros metálicos ha despertado gran interés en las últimas décadas debido al agotamiento de menas de alto grado, lo cual hace necesario encontrar nuevas tecnologías "ambientalmente más amigables" que permitan el beneficio de menas de bajo tenor (Brierley y Luinstra, 1993; Marsden y House, 1992; Watling, 2006).
Los sulfuros utilizados en este trabajo presentan gran importancia industrial. La esfalerita (ZnxFe1_xS) es la fuente primaria de zinc en el mundo, presentando gran interés en aplicaciones industriales como la galvanización de aceros, la obtención de latón y en fabricación de pinturas (Harmer et al., 2006; Haghshenas et al., 2009).
La calcopirita (CuFeS2) es la fuente primaria de cobre en el mundo, representando el 70% de las reservas. Su beneficio se hace generalmente por pirometalurgía; sin embargo, esta tecnología genera contaminaciones de SO2, principal causante de las lluvias ácidas. La investigación se ha centrado, por tanto, en la hidrometalurgia donde la extracción de cobre se ve limitada por su carácter refractario (Dutrizac, 1981; Rivadeneira, 2006). La galena (PbS) es un mineral de gran importancia desde dos puntos de vista, 1) como fuente primaria de plomo, y 2) como material semiconductor, con un bandgap aproximado de 0,4 eV (Muscat y Gale, 2003). Además, este mineral es importante desde el punto de vista ambiental ya que a menudo es responsable de aumentar la acidificación de las aguas luego de la explotación minera (Muscat y Gale, 2003).
La adaptación de animales, plantas y microorganismos al ambiente es un fenómeno natural. Los organismos adaptados a ambientes específicos por largos períodos de tiempo presentan mejores resultados (Elzeky y Attia, 1995). En los procesos de lixiviación se realizan generalmente varias etapas de adaptación del microorganismo con el fin de obtener una mejor velocidad de disolución del metal de interés (Shahverdi et al., 2001; Shi y Fang, 2004; Xia et al., 2008). En los procesos biohidrometalúrgicos, la adaptación a iones metálicos realizada en cultivos selectivos es el método más popular para lograr una buena actividad de las cepas (Haghshenas et al., 2009).
Los microorganismos comúnmente usados en la lixiviación de sulfuros, Acidithiobacillus ferrooxidans y Acidithiobacillus thiooxidans metálicos son mesófilos, se presentan en intervalos de temperatura entre los 20 y 45 ºC; los acidófilos viven en ambientes cuya acidez puede bajar hasta pH < 1, y los quimiolitótrofos, que derivan la energía para llevar a cabo sus procesos metabólicos de compuestos inorgánicos (Bozdemir et al., 1996; Rawling et al., 1999; Daoud y Karamanev, 2005), los cuales obtienen la energía necesaria de la oxidación de compuestos reducidos del azufre y del Fe2+ a Fe3+. La diferencia entre estos dos microorganismos es que Acidithiobacillus thiooxidans no es capaz de utilizar la oxidación del ion ferroso como fuente de energía (Bozdemir et al., 1996; Rawling et al., 1999; Cabrera et al., 2005; Daoud y Karamanev, 2006).
La exposición de cultivos mesófilos a concentrados de sulfuros metálicos genera inhibición o disminución en su actividad, especialmente a altas densidades de pulpa. Este fenómeno se debe a varios factores: 1) menor cantidad de oxígeno disuelto, 2) daño a la pared celular como resultado de la fricción entre las partículas minerales y los microorganismos, y 3) efecto nocivo de los iones metálicos como Pb2+, Cu2+ y Zn2+ (Cabrera et al., 2004; Akcil et al., 2007). Sin embargo, estas bacterias pueden ser adaptadas para tolerar estos factores inhibitorios (Cabrera et al., 2004; Xia et al., 2008). El método generalmente usado para adaptar los microorganismos al mineral, en estos proceso, consiste en una serie de cultivos sucesivos en los cuales se incrementa la densidad de pulpa y se disminuye la fuente de energía principal, obligando a los microorganismos a crecer a expensas de la disolución del mineral (Haghshenas et al., 2009).
En la biolixiviación de concentrados de esfalerita, calcopirita y galena, un factor que retarda e inhibe la actividad bacteriana es la toxicidad de altas concentraciones de Zn2+, Cu2+ y Pb2+ que se encuentran en la solución (Rawling et al., 1999; Dopson et al., 2003; Xia et al., 2008). Gentina y Acevedo (2005) encontraron niveles inhibitorios para Acidithiobacillus ferrooxidans. Para el caso del Cu2+ y Zn2+, una concentración mayor a 10000 mg/L generó efectos inhibitorios. Aston et al. (2010) observaron que, para el caso del plomo, solo 0,036 mg/L generaban inhibición. Xia et al. (2008) y Haghshenas et al. (2009 a y b) encontraron que se daba una mejor lixiviación del Cu2+ en las cepas adaptadas de Acidithiobacillus ferrooxidans. Li y Jun-Ke (2000) encontraron resultados similares para la disolución de Zn2+. Por tanto, una buena adaptación de los microorganismos al mineral aumenta la tasa de disolución de los metales de interés.
El propósito de este trabajo fue comparar la adaptación de una cepa compatible con Acidithiobacillus ferrooxidans a altas densidades de pulpa de calcopirita, esfalerita y galena, aumentando gradualmente el porcentaje de pulpa y disminuyendo la fuente de energía —sulfato ferroso—, y trabajando con dos distribuciones de tamaño de partícula.
Materiales y métodos
Obtención y caracterización inicial del mineral
Los experimentos se realizaron usando muestras naturales de mineral tomadas en varias minas del departamento de Antioquia. La esfalerita se tomó de la mina El Centeno (Buriticá), la galena fue provista por la empresa minera Frontino Gold Mine (Segovia), y la calcopirita fue tomada de la mina La Chorrera (Cisneros). Los minerales fueron sometidos, individualmente, a un proceso de conminución y concentración gravimétrica en mesa Wilfley, con el fin de concentrar la fase de interés. Para el caso de la esfalerita, fue necesario realizar un proceso de flotación espumante. Los concentrados fueron molidos en mortero de ágata y tamizado con el fin de asegurar dos distribuciones de tamaños de partícula: ?200 y -325 de acuerdo con la serie Tyler de tamices. El mineral se esterilizó en autoclave a 18 psi y 120 ºC por 20 min. Mediante el uso del microscopio óptico de luz plana polarizada, modo luz reflejada, marca Leitz Laborolux 11POL, con objetivos de aumento de 10X y 20X en aire y 32X y 50X de inmersión en aceite, se estimó por conteo de puntos, según norma ASTM D 2799 de 2009, la composición mineralógica resultante.
Microorganismos empleados
Se utilizó una cepa compatible con Acidithiobacillus ferrooxidans, cedida por el laboratorio de biomineralogía de la Universidad Nacional de Colombia, sede Medellín. La cepa es proveniente de la mina de carbón "La Angostura" manto "El Vampiro II" ubicada en el municipio de Morales, Cauca. En el estudio hecho por Cardona (2007) se verificó la compatibilidad de dichos microorganismos con Acidithiobacillus ferrooxidans por medio de pruebas bioquímicas. Los microorganismos fueron crecidos previamente en medio T&K con una composición química de 0,5% de (NH4)SO4; 0,5% de MgSO4.7H2O; 0,5% de K2HPO4 y 33,33 g/L de sulfato ferroso. Durante este proceso se logró aumentar la concentración celular y obtener una población idónea para inocular los ensayos ~ 108 células/ml, estimada a partir de conteo en cámara de Newbauer.
Ensayos de adaptación
Los microorganismos fueron adaptados independientemente a concentrados de calcopirita, esfalerita y galena, en medio T&K, con reemplazo sucesivo de sulfato ferroso por concentrado de mineral. La primera etapa consistió en 2,5% (p/v) de mineral y 10% (v/v) de sulfato ferroso, el segundo subcultivo 5% (p/v) de mineral y 7,5% (v/v) de sulfato ferroso, el tercer subcultivo 7,5% (p/v) de mineral y 5% (v/v) de sulfato ferroso, el penúltimo subcultivo constó de 10% (p/v) de mineral y 2,5% (v/v) de sulfato ferroso, y el último periodo de adaptación se hizo con 10% (p/v) de mineral y fue inoculado sin adición externa de sulfato ferroso. Los medios fueron acidificados a pH 1,8 con ácido sulfúrico (H2SO4) al 98%; esta solución fue esterilizada en autoclave a 120 ºC por 20 min. Los experimentos se inocularon con 10% (v/v) y una población de 108 células/ml, la cual fue determinada por conteo en cámara de Newbauer. Los ensayos de biolixiviación se llevaron a cabo por 14 días en erlenmeyers de 250 ml, utilizando un volumen de trabajo de 100 ml. Se hizo control de evaporación, diariamente, por pérdida de peso. Los ensayos fueron incubados con agitación de 180 rpm ± 2 rpm y 30 ºC ± 1 ºC. Todas las condiciones fueron replicadas y se incluyó el respectivo control abiótico.
Medidas de Eh y pH
Cada dos días se hizo un seguimiento de la cinética del proceso mediante medidas de pH (HACH HQ40d multi PHC30103) con electrolito KCl y potenciales de óxido reducción (Shot Handylab 1 Pt 6880) in situ (electrodo de referencia Ag0/AgCl). Las medidas se hicieron en cámara de flujo laminar horizontal para garantizar las condiciones de esterilidad en los electrodos empleados y en el sistema. En los ensayos con concentrado de galena (tanto para los ensayos inoculados como los controles abióticos), fue necesario acidificar el sistema con ácido sulfúrico (H2SO4) con el fin de evitar efectos nocivos en los microorganismos ya que la galena consume protones, aumentando el pH hasta valores superiores a 5,0 como ha sido reportado por Bhatti et al. (1993) y García et al. (1995).
Resultados
Caracterización inicial de los minerales
Los análisis al óptico mostraron que para la esfalerita la fase mineral principal fue esfalerita (ZnS) 80% con cantidades menores de pirita (FeS2) 4%, calcopirita (CuFeS2) 10% y cuarzo (SiO2) 6%; la composición de la calcopirita fue 88% de calcopirita (CuFeS2), 9% de ganga (SiO2), 1% de covelita (CuS) y 2% de molibdenita (MoS2), para el caso de la galena la concentración fue 93,3% de galena (PbS), 6,2% de esfalerita (ZnS) y 0,5% de calcopirita (CuFeS2).
Calcopirita (CuFeS2). Potencial de óxido-reducción (Eh) y pH
En general, todos los subcultivos presentaron una fase de adaptación seguida de una fase estacionaria para finalmente estabilizarse hacia el final de proceso (figura 1 a, c, e y g). Sin embargo, se presentaron pequeñas variaciones en los subcultivos. En la primera etapa la fase de adaptación o fase lag duró 6 días para el tamaño de partícula más grueso, mientras que para el tamaño de partícula más fino duró solo 2 días (figura 1a). En la segunda etapa el tamaño de partícula más fino presentó un comportamiento similar que en la etapa anterior, siendo un poco más larga la fase lag (alrededor de 8 días), mientras que la malla más gruesa mostró un crecimiento exponencial desde el inicio del proceso (figura 1 c). En el tercer subcultivo, los dos tamaños de partícula presentaron fase lag hasta el día 4, seguido de una fase exponencial para finalmente estabilizarse. El ensayo realizado con tamaño de partícula más fino presentó un mayor Eh (620 mV) (figura 1 e).
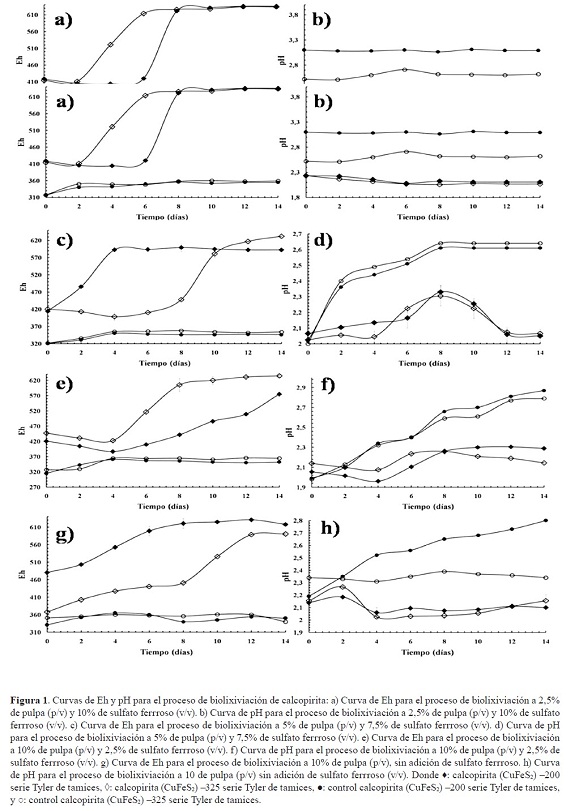
En el último periodo de adaptación el comportamiento fue similar. Sin embargo, la fase de adaptación fue mayor para el tamaño de partícula más fino, mientras que el tamaño más grueso duró 2 días (figura 1g). Es importante anotar que los ensayos no inoculados no presentan cambios aparentes en ninguno de los subcultivos (figura 1 a, c, e y g). En todos los subcultivos se observó que al aumentar el contendido de pulpa se presentó un leve incremento en el pH (figura 1b, d, f y h). En el caso del segundo y cuarto subcultivo el pH disminuye y se estabiliza al final de proceso, siendo más rápido este descenso para el mayor porcentaje de pulpa (figura 1 d y h). El pH en los ensayos no inoculados fue estable solo en el primer subcultivo (figura 1 b). El segundo y tercer subcultivo exhibieron un incremento progresivo del pH en el tiempo (alrededor de 2,8) (figura 1 d y f). En el último subcultivo el tamaño de partícula más grueso presentó un incremento a lo largo del proceso, mientras que el tamaño más fino no mostró cambios aparentes (figura 1 h).
Esfalerita (ZnxFe1xS). Potencial de óxido-reducción (Eh) y pH
En general, todos los subcultivos presentaron un comportamiento similar al concentrado de calcopirita, mostrando una fase de adaptación seguida de una fase de crecimiento exponencial y finalmente una fase estacionaria (figura 2 a, c, e y g). Sin embargo, al igual que para el concentrado de calcopirita, se presentaron pequeñas variaciones en los subcultivos. En la primera etapa, la fase lag duro 4 días para ambos tamaños de partícula, seguido de un incremento exponencial hasta el final de proceso (figura 2 a). En el segundo subcultivo se observó un crecimiento exponencial desde el inicio del proceso para ambos tamaños de partícula (figura 2 c). El tercer subcultivo presentó una fase de adaptación de cuatro días para ambos tamaños de partícula, seguida de un crecimiento exponencial para finalmente estabilizarse en el tiempo (figura 2 e). El cuarto subcultivo evidenció una fase lag mayor para el tamaño de partícula más fino (6 días), seguida de un crecimiento exponencial hasta el final de proceso. Mientras que el tamaño de partícula más grueso mostró una fase lag de dos días seguida de un crecimiento exponencial para finalmente estabilizarse (figura 2 g). Los ensayos no inoculados no presentaron cambios aparentes en ninguno de los subcultivos de adaptación (figura 2 a, b, e y g). El comportamiento del pH en general para todos los subcultivos presentó un incremento en los primeros días del proceso para finalmente descender y estabilizarse (figura 2 b, d, f y h). Este aumento inicial en el pH fue más marcado en el último subcultivo (figura 2 g). Los ensayos no inoculados mostraron un aumento progresivo del pH hasta estabilizase 199 en todos los 200 subcultivos, alcanzando mayor acidez a mayor porcentaje de pulpa (figura 2 b, d, f y h).

Galena (PbS). Potencial de óxido-reducción (Eh) y pH
Al igual que para el concentrado de calcopirita y esfalerita, se observó la presencia de tres fases características en la curvas de Eh: fase de adaptación o fase lag, fase de crecimiento exponencial y fase estacionaria (figura 3 a, c, e y g). Para los dos primeros subcultivos el comportamiento es muy similar, presentando una fase de adaptación de 4 días para el primero y de 2 días para el segundo, seguido de una fase de crecimiento exponencial hasta el final de proceso (alcanzando valores alrededor de 600 mV) (figura 3 a y c). Los dos últimos subcultivos presentaron fases de adaptación más largas (6 y 8 días respectivamente), seguidas de una fase de crecimiento exponencial. Los valores de Eh alcanzados son menores, alrededor de 550 mV (figura 3 e y g). En general, se observó un aumento generalizado del pH en todas las etapas de adaptación, a pesar de la acidificación del sistema con ácido sulfúrico (hasta el día 6), pero a partir del día sexto el pH decae y se estabiliza en el tiempo (figura 3 b, d, f y h). Los ensayos no inoculados presentaron un comportamiento creciente para todos los subcultivos (figura 3 b, d, f y h). Sin embargo, en el tercer y el cuarto subcultivos se observa un descenso a partir del día 8 (figura 3 f y h). Cabe anotar que en todos los subcultivos los valores de Eh y pH son similares durante todo el proceso para los dos tamaños de partícula.

Discusión
La importancia del pH
A continuación se discuten los comportamientos obtenidos para los concentrados de esfalerita, calcopirita y galena durante el período de adaptación. El pH es un factor determinante en el proceso de adaptación de los microorganismos a los diferentes minerales e influye de forma significativa en la velocidad de crecimiento de los mismos, debido a que afecta a los grupos ionizables presentes en las enzimas, situadas en el citoplasma y periplasma de la célula (Das et al., 1999; Gómez y Cantero, 2005).
El aumento inicial en el pH puede deberse a varios factores: 1) La presencia de carbonatos y fases silicatadas en contacto con la solución lixiviante genera consumo de ácido (Sampson et al., 2005; Arroyave, 2007); 2) el consumo de protones generado principalmente en los monosulfuros como calcopirita, esfalerita y galena siendo mayor en este último (García et al., 1995; Da Silva, 2004; Ballester, 2005), y 3) a la oxidación del Fe2+ (Bevilaqua et al., 2002; Meruane y Vargas, 2003).
El aumento inicial de pH en todos los subcultivos de adaptación para el caso de la esfalerita se debe aparentemente a la presencia de carbonatos en el concentrado, ya que este contenía gran cantidad de ganga (alrededor del 6%). El aumento pudo inhibir el adecuado crecimiento de los microorganismos y por tanto retardar la adaptación (Das et al., 1999). Sin embargo, en todos los subcultivos se observa estabilidad en el pH hacia el final de proceso (figura 2 b, d, f y h), lo cual sugiere una neutralización de los carbonatos, ya que este mineral en presencia de ácido sulfúrico se neutraliza formando yeso y otro tipo de sulfatos dependiendo de la composición química del carbonato (Márquez, 1999). En general, el incremento en el pH fue mayor para las muestras más finas, lo cual pudo deberse a que un menor tamaño de partícula se traduce en mayor área superficial y, por tanto, una mayor reactividad, generando un consumo de protones más elevado. Además, a menor tamaño de partícula se aumenta el estrés hidrodinámico para los microorganismos, lo cual pudo evitar que estos favorecieran la neutralización de carbonatos más rápidamente como ocurrió para el mayor tamaño de partícula (figura 2 b, d, f y h). Estos resultados están de acuerdo con lo observado por Silverman (1961), Deveci (2004) y Haghshenas et al. (2009 a y b). Por otra parte, Deveci (2002) encontró que a menor tamaño de partícula hay mayor posibilidad de colisiones, las cuales son las responsables del daño celular o incluso de la muerte de los microorganismos. El consumo de ácido también puede ser atribuido a la oxidación del Fe2+; no obstante, este proceso genera un aumento menor en el pH (~3), según lo reportado por Bevilaqua et al. (2002) y Meruane y Vargas (2003).
El aumento generalizado del pH inicial en todas las etapas de adaptación para el caso de la galena es interpretado como consumo de protones debido a su carácter de monosulfuro. El consumo de protones para este mineral es mucho mayor que para la calcopirita y esfalerita como ha sido reportado por varios autores (García et al., 1995; Da Silva, 2004; Mejía et al., 2007), lo que hizo necesario acidificar el sistema hasta agotar la demanda de protones, la cual se dio aproximadamente el día sexto, a partir del cual se observó una paulatina disminución en los valores de pH. Esto es debido a que la disolución de galena en ambientes ácidos ocurre como resultado de la protonación en la superficie del mineral, donde el único mecanismo de protonación en solución acuosa energéticamente favorable se genera por la unión de tres H+ en los tres átomos superficiales de azufre, los cuales rodean un átomo de plomo sustituido por un cuarto ion H+ (Gerson y O’Dea, 2003; Acero et al., 2007). Estas teorías explican el decrecimiento en el valor del pH alrededor del sexto día para estabilizarse alrededor de 2,0 y más bajos en la última etapa de adaptación, con 1,8 (figura 3 b, d, f y h), indicando de este modo un incremento en la concentración de H+ producido por la actividad bacteriana. En este caso, al igual que en la esfalerita, el aumento del pH fue mayor para el tamaño de partícula -325 serie Tyler de tamices confirmando el efecto adverso del menor tamaño de partícula sobre los microorganismos, debido posiblemente a mayores colisiones entre las partículas.
Para el caso de la calcopirita, el comportamiento en el pH fue muy estable en todo el proceso de adaptación, presentando un ligero incremento al inicio de todas las etapas, explicado como la adaptación de los microorganismo al mineral, el cual se estabiliza rápidamente (Bevilaqua et al., 2002; Meruane y Vargas 2003). Este comportamiento da indicios de una buena adaptación y actividad bacteriana (Cox et al., 1977; Tekeuchi y Susuki, 1994).
La importancia del potencial de óxido-reducción (Eh)
El potencial de óxido-reducción puede relacionarse con el crecimiento y la actividad bacteriana. Un rápido incremento en este se puede deber a una buena adaptación de los microorganismos al mineral, indicando indirectamente que los iones solubilizados aparentemente no inhiben la capacidad bacteriana de oxidar el ion ferroso (Xia et al., 2008). Además, todos los concentrados de minerales empleados presentaron tres fases características en las curvas de crecimiento bacteriano: fase de adaptación o fase lag (los microorganismos adaptan su metabolismo a las nuevas condiciones ambientales), fase de crecimiento exponencial (las bacterias consumen los nutrientes del medio a velocidad máxima, y su velocidad de crecimiento es la máxima) y fase estacionaria (no se incrementa el número de bacterias, los microorganismos entran en fase estacionaria bien porque se agota algún nutriente esencial del medio o porque los productos de desecho que han liberado durante la fase de crecimiento exponencial hacen que el medio sea inhóspito) (Madigan et al., 2003). Cabe mencionar que aunque las concentraciones de los iones metálicos no fueron medidas al inicio y final del proceso, fue posible observar turbidez, cambio de color y olor en los medios de cultivo a medida que avanzaba el desarrollo para todos los ensayos, lo que da indicios de cambios en el sistema; además, al final del proceso fue posible observar una buena población bacteriana indicando que los iones solubilizados no generaron alta toxicidad en los medios de cultivo.
A continuación se discuten los comportamientos obtenidos por la esfalerita, calcopirita y galena en todos los periodos de adaptación.
El tamaño de partícula más fino presentó un mejor desempeño del Eh para todos los subcultivos debido a que a menor tamaño de partícula el mineral posee un área superficial mayor y su reactividad aumenta. Al haber una mayor fuente de Fe2+ para los microorganismos, se da una rápida transformación de Fe2+ a Fe3+ obteniéndose de este modo altos valores de potencial. Además, a bajas concentraciones de pulpa los daños debidos a las colisiones en las partículas son menores (Deveci, 2002). Sin embargo, para el caso del concentrado de esfalerita, aunque el ion Zn2+ es menos tóxico que el Cu2+ en solución, la presencia de carbonatos aparentemente afectó el buen desarrollo de los microorganismos, siendo menos evidente para el tamaño de partícula -200 serie Tyler de tamices. Además, esto también podría estar relacionado con la reactividad del mineral en función del tamaño de partícula. Por otra parte, el comportamiento del Eh para el concentrado de galena se debe a la gran toxicidad del plomo para los microorganismos, donde el tamaño de partícula juega un papel fundamental —a menor tamaño de partícula se tiene mayor concentración de Pb2+ en solución—.
En el subcultivo realizado para 5% (p/v) de mineral y 7,5% (v/v) de sulfato ferroso, la fase lag fue menor, alrededor de 2 días, mostrando de forma indirecta una buena adaptación de los microorganismos al mineral. El crecimiento del potencial de óxido-reducción es mejor, al igual que en la etapa anterior, para el tamaño de partícula más grueso en el caso de la esfalerita y la galena. En esta etapa la calcopirita mostró un mejor comportamiento para el tamaño de partícula más grueso, lo cual da indicios que a mayor concentración de Cu2+ los microorganismos son afectados por la toxicidad de este ion, lo cual dificulta su etapa de adaptación (Rawling et al., 1999; Rawling, 2005). Para el ensayo de adaptación realizado con 7,5% (p/v) y 5% (v/v) de sulfato ferroso —datos no mostrados—, el comportamiento fue muy similar al obtenido en la segunda etapa de adaptación.
Para el caso de 10% (p/v) de pulpa y 2,5% (v/v) de sulfato ferroso (figura 1e, 2e y 3e), el crecimiento del potencial redox fue muy similar para todos los minerales. No se observó fase estacionaria. Sin embargo el crecimiento, aunque constante, es lento hasta aproximadamente el día sexto del proceso. Este comportamiento indica que los microorganismos se van adaptando a las condiciones del sistema paulatinamente, usando su energía para tolerar las condiciones adversas y no para generar una rápida oxidación del Fe2+ a Fe3+; además, pueden toman la fuente de energía de las trazas de Fe2+ provenientes del inóculo bacteriano, la cual es de más fácil acceso. Finalmente, se logró una buena adaptación luego del día sexto, donde se presenta un fuerte incremento en el Eh, indicando que los iones dejados en solución no afectaron aparentemente la actividad oxidativa de los microorganismos, lo que está de acuerdo con lo observado por Xia et al. (2008).
Al disminuir totalmente la fuente de energía (figura 1g, 2g y 3g) el crecimiento del Eh fue constante pero más lento que en la etapa anterior, indicando que aunque los microorganismos se adaptaron al sistema, les costó más tiempo tolerar las condiciones adversas. Además, como los microorganismos no contaban con fuente de energía extra, y como se agotan las trazas de Fe2+ provenientes del inóculo, se vieron obligados a tomar la fuente de energía de los iones de Fe2+ provenientes de la disolución en medio ácido de los minerales, proceso que tiene una cinética más lenta (Zapata et al., 2007). Alrededor del día 8 se dio un marcado incremento indicando, en principio, una buena actividad bacteriana para la oxidación del ion ferroso.
De acuerdo con Touvinen et al. (1971), la tolerancia del microorganismo al mineral es adquirida durante la fase lag, lo cual puede ser observado en la fase estacionaria de crecimiento del Eh y en el aumento inicial del pH.
La evolución del potencial de óxido-reducción se relaciona con la tasa de crecimiento bacteriano y su capacidad de oxidar el Fe2+ (Gentina y Acevedo, 2005). Esta capacidad oxidativa aumentó a través del tiempo en las cuatro etapas de adaptación, donde las bacterias alcanzaron valores de Eh alrededor de 600 mV en corto tiempo y los mantuvieron hasta el final de proceso, especialmente en las primeras etapas de adaptación, en las que aún tenían fuente de energía adicional. En la última fase de adaptación, sin adición de fuente de energía, el crecimiento de la curva de Eh para los sulfuros es más lento, aunque el valor aumenta paulatinamente, lo que sugiere que los microorganismos toman la fuente de energía necesaria para su crecimiento de la oxidación del Fe2+ proveniente del mineral (Prayuenyong, 2002). Esto da indicios de una buena actividad oxidativa y de que las bacterias respondieron favorablemente a las etapas de adaptación, llevándose a cabo el mecanismos indirecto (Prayuenyong, 2002; Ballester, 2005). El aumento del Eh sugiere que en presencia de bacterias hay un transporte constante de electrones en el sistema, como la trasformación de Fe2+ a Fe3+, función principal de A. ferrooxidans (Rossi, 1990; Kodali et al., 2004).
Además de los ensayos de adaptación para una cepa compatible con A. ferrooxidans se realizaron ensayos alternos de adaptación para una cepa compatible con A. thiooxidans (datos no mostrados). Cuando se adicionó azufre elemental como fuente de energía extra, los ensayos mostraron producción de ácido, evidenciado por la disminución del pH (0,6), y los valores de potencial aumentaron hasta aproximadamente 460 mV para todos los ensayos. Sin embargo, al ir disminuyendo la fuente externa de energía, este microorganismo fue perdiendo la habilidad de producir ácido. Al suprimir totalmente la fuente de energía los valores de pH obtenidos fueron similares a los controles abióticos lo cual da indicios de que este microorganismo no fue capaz de obtener energía del mineral (Bevilaqua et al., 2002).
Conclusiones
La adaptación de una cepa compatible con Acidithiobacillus ferrooxidans en etapas sucesivas mediante la disminución gradual de fuente de energía y aumento en el contenido del mineral (calcopirita, esfalerita y galena respectivamente), resultó ser un método eficaz para este tipo de procesos. Esta metodología provee una adecuada adaptación de los microorganismos al mineral.
La metodología empleada permitió observar que la presencia de iones metálicos afecta la evolución del crecimiento bacteriano y la oxidación del ion ferroso. La cepa compatible con A. ferrooxidans mostró más tolerancia a los iones de zinc, cobre y plomo para el tamaño de partícula -200 serie Tyler de tamices. Para el tamaño de partícula -325 serie Tyler de tamices el microorganismo presentó más tolerancia al Cu2+, seguido por el Zn2+ y finalmente al Pb2 +, esto debido al efecto del tamaño de la partícula y a la presencia de carbonatos en el concentrado de esfalerita. Como recomendación, es necesario complementar este estudio con técnicas moleculares que permitan verificar la presencia de A. ferrooxidans y su viabilidad en este tipo de procesos. Además, hacer medidas de hierro, sulfatos, cobre, plomo y zinc que permitan verificar el aumento de dichos iones a través del proceso.
Agradecimientos
Los autores agradecen al programa de biotecnología de Colciencias; a los laboratorios de biomineralogía, Cimex, preparación de rocas y carbones de la Universidad Nacional de Colombia, sede Medellín: al laboratorio de Ingeominas, Medellín; a I. C. Cardona de la Universidad de West Virgina, USA, por sus comentarios. ALM agradece al CODI de la Universidad de Antioquia.
Referencias bibliográficas
1 Acero, P., Cama, J., Ayora, C. 2007. Rate law for galena dissolution in acidic environment. Chemical Geology, 245 (007): 219-229. [ Links ]
2 Akcil, A., Ciftci, H., Deveci, H. 2007. Role and contribution of pure and mixed cultures of mesophiles in bioleaching of a pyritic chalcopyrite concentrate. Mineral Engineering, 20: 310-318. [ Links ]
3 Arroyave, D. 2007. Evaluación del proceso de biooxidación a escala de laboratorio del mineral de la mina El Zancudo, Titiribí, Antioquia. Tesis de Maestría. Universidad Nacional de Colombia Sede Medellín. [ Links ]
4 Aston, E., Peyton, M., Lee, B., Apel, A. 2010. Effects of ferrous sulfate, inoculum history, and anionic form on lead, zinc, and copper toxicity to Acidithiobacillus caldus strain BC13. Environmental Toxicology and Chemistry, 29 (12): 2669-2675. [ Links ]
5 Ballester, A. 2005. Mecanismos de la biolixiviación, capítulo 1. En: Fundamentos y perspectivas de las tecnologías biomineras, 9-24. [ Links ]
6 Bevilaqua, D., Leite, A., García, Jr O., Touvinen, O. 2002. Oxidation of Chalcopyrite by Acidihiobacillus ferrooxidans and Acidithiobacillus thiooxidans in shake flasks. Process Biochemistry, 38: 587-592. [ Links ]
7 Bhatti, T., Bigham, J., Vuorinen, A., Tuovinen, O. 1993. Biological leaching of sulfides with emphasis on pyrrhotite and pyrite. In: Proceedings of International Symposium on Biotechnology for Sustainable Development, Kauser A, Malik, Anwar Nasim & Ahmad M. Khalid, NIBGE, Faisalabad, Pakistan, 299-308. [ Links ]
8 Bozdemir, T., Durusoy, T., Erincin, E., Yürüm, Y. 1996. Biodesulfurization of Turkish lignites. Fuel, 75 (13): 1596-1600. [ Links ]
9 Brierley, J., Luinstra, L. 1993. Biooxidation-heap concept for pretreatment of refractory gold ore. In: Biohydrometallurgical Biohydrometallurgical Technologies, A. E. [ Links ]
10 Cabrera, G., Gómez, J., Cantero, D. 2005. Kinetic study of ferrous sulphate oxidation of Acidithiobacillus ferrooxidans in the presence of heavy metal ions. Encyme and Microbial Technology, 36: 301-306. [ Links ]
11 Cardona, I. 2008. Mineralogía del proceso de biodesulfurización de carbones provenientes de la zona río Guachinte-río Asnazú (Valle del Cauca y Cauca). Tesis de Maestría, Materiales y Procesos. Universidad Nacional de Colombia, Sede Medellín. [ Links ]
12 Cox, J., Nicholas, D., Inglew, W. 1977. Transmembrane electrical potencial and transmembrane pH gradient in the acidophilic Thiobacillus ferrooxidans. Biocem, J, 178: 195-200. [ Links ]
13 Da Silva G. 2004. Kinetics and mechanism of the bacterial and ferric sulphate oxidation of galena. Hydromatallurgy, 75: 99-110. [ Links ]
14 Daoud J., Karamanev, D. 2006. Formation of coal jarosite during Fe2+ oxidation by Acidithiobacillus ferrooxidans. Mineral Engineering, 19: 960-967. [ Links ]
15 Das, T., Ayyappan, S., Chaudhury, G. R. 1999. Factors Affecting Bioleaching Kinetics of Sulfide Ores Using Acidophilic Microorganisms. BioMetals, 12: 1-10. [ Links ]
16 Deveci H. 2002. Effect of solids on viability of acidophilic bacteria. Minerals Engineering, 15: 1181-1189. [ Links ]
17 Deveci, H., 2004. Effect of particle size and shape of solids on the viability of acidophilic bacteria during mixing in stirred tank reactors. Hydrometallurgy, 71: 385-396. [ Links ]
18 Dutrizac, J. E. 1981. The dissolution of chalcopyrite in ferric sulfate and ferric chloride media. Met. Trans. B. 12B: 371-378. [ Links ]
19 Elzety, M., Attia, Y. 1995. Efect of bacterial adaptation on kinetics and mechanisms of bioleaching ferrous sulfides. The Chemical Engineering Journal, 56: 115-124. [ Links ]
20 García, Jr., O., Bigham, J., Tuovinen, O. 1995. Oxidation of galena by Thiobacillus ferrooxidans and Thiobacillus thiooxidans. Canadian Journal Microbiology, 41: 508-514. [ Links ]
21 Gentina, J., Acevedo, F. 2005. Biolixiviación de minerales de oro. In: Fernando Acevedo y Juan Carlos Gentina (eds.). Fundamentos y perspectivas de las tecnologías biomineras. 79-91. [ Links ]
22 Gerson, A., O'Dea, A. 2003. A quantum chemical investigation of the oxidation and dissolution mechanisms of Galena. Geochim. Cosmochim. Acta 67 (5): 813-822. [ Links ]
23 Gómez, J., Cantero, D. 2005. Biooxidación del ión ferroso. In: Fernando Acevedo y Juan Carlos Gentina (eds.). Fundamentos y perspectivas de las tecnologías biomineras. 25-43. [ Links ]
24 Haghshena, D. F., Keshavarz, A. E., Amouei, T., Bonakdarpour, B., Nasernejad, B. 2009a. Adaptation of Acidithiobacillus ferrooxidans to high grade sphalerite concentrate. Minerals Engineering, 22: 1299-1306. [ Links ]
25 Haghshenas, D., Keshavarz, A., Bonakdarpour, B., Darvishi, D., Nasernejad, B. 2009b. Hydrometallurgy. Article in press doi:10.1016/j.hydromet.2009.08.007. [ Links ]
26 Harmer, S., Thomas, J., Fornasiero, D., Gerson, A. 2006. The evolution of surface layers formed during chalcopyrite leaching. Geochimica et Cosmochimica Acta, 70: 4392-4402. [ Links ]
27 Hong-Mei, L., Jia-Jun, K. 2001. Technical note influence of Cu 2+ and Mg 2+ on the growth and activity of Ni 2+ adapted Thiobacillus ferrooxidans. Minerals Engineering, 14: 113-116. [ Links ]
28 Kodali, B., Rao, B., Narasu, L., Pogaku, R. 2004. Effect of biochemical reactions in enhancement of rate of leaching. Chemical Engineering Science, 59: 5069-5073. [ Links ]
29 Madigan, M., Martinko, J., Parker, J. 2003. Biología de los microorganismos. 10 ed. Madrid: Pearson Educación. [ Links ]
30 Márquez, M. 1999. Mineralogia dos processos de oxidação sobre pressão e bacteriana do minerio de ouro da mina São Bento, MG. Tese de doutorado. Universidad de Brasilia. [ Links ]
31 Marsden, J., House, I. 1992. The chemistry of gold extraction. England: Ellis Horwood Limited. [ Links ]
32 Mejía, R., Ospina, C., Márquez, M. 2007. Mineralogía del proceso de oxidación bacteriana de la galena (PbS) por Acidithiobacillus ferrooxidans, mediante análisis de microscopía electrónica de barrido, FTIR, y DRX. Scientia et Technica, 36: 549-554. [ Links ]
33 Meruane, G., Vargas, T. 2003. Bacterial oxidation of ferrous iron by Acidithiobacillus ferrooxidans in the pH range 2,5-7,0. Hydrometallurgy, 71 (1-2): 149-158. [ Links ]
34 Muscat, J., Gale, J. 2003. First principles of the surface of galena PbS. Geochemica at Cosmochimica Acta, 67 (5): 799-805. [ Links ]
35 Prayouenyong, P. 2002. Coal biodesulphurization. Journal Science Technology, 24: 493-507. [ Links ]
36 Rawlings, D. E. 2005. Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microbial Cell Factories, 4: 13. [ Links ]
37 Rawlings, D. E., Tributsch, H., Hansford, G. 1999. Reasons why "Leptospirillum "-like species rather than Thiobacillus ferrooxidans are the dominant iron-oxidizing bacteria in many commercial processes for the biooxidation of pyrite and related ores. Microbiology, 145, 5-13. [ Links ]
38 Rivadeneira, J. 2006. Introduction. Mining innovation in Latin America Report. Publication via on-line (http://www.mininginnovation.cl/content.htm). Santiago, Chile. 6-7. [ Links ]
39 Rossi, G. 1990. Biohydrometallurgy. Hamburg: McGraw-Hill Book Company GmbH. [ Links ]
40 Sampson, M., van der Merwe, J., Harvey, T., Bath, M. 2005. Testing the ability of a low grade sphalerite concentrate to achieve autothermality during biooxidation heap leaching Minerals Engineering. 18: 427-437. [ Links ]
41 Shahverdi, A., Yazdi, M., Oliazadeh, M., Darebidi, M. 2001. Biooxidation of mouteh refractory gold-bearing concentrate by an adapted Thiobacillus ferrooxidans. Journal of Sciences, Islamic Republic of Iran, 12 (3): 209-212. [ Links ]
42 Shi, S., Fang, Z. 2005. Bioleaching of marmatite flotation concentrate by adapted mixed mesoacidophilic cultures in an air-lift reactor. International Journal of Mineral Processing, 76: 3-12. [ Links ]
43 Silverman, M., Rogoff, M., Wender, I. 1961. Bacterial oxidation of pyritic materials in coal. Appl. Microbiol., 9 (6): 491-496. [ Links ]
44 Tang, K., Baskaran, V., Nemati, M. 2009. Bacteria of sulphur cycle: an overview of microbiology, biokinetics and their role in petroleum and minig industries. Biochemical Engineering Journal, 44: 73-94. [ Links ]
45 Tekeuchi, T., Susuki, I. 1994. Effect of pH sulfite oxidation by Thiobacillus cell with sulfurous acid or sulfur dioxide as posible sustrate. J. Bacteriol., 176 (3): 913-916. [ Links ]
46 Tuovinen, H., Niemela, I., Gyllenberg, G. 1971. Tolerance of Thiobacillus ferrooxidans to some metals. Antonie van Leewenhoek, 37: 489-496. [ Links ]
47 Waltling, H. R. 2006. The bioleaching of sulphide minerals with emphasis on copper sulphides-a review. Hydrometallurgy, 84 (1-2): 81-108. [ Links ]
48 Xia, L., Liu, J., Xiao, L., Zeng, J., Li, B., Geng, M., Qiu, G. 2008. Single and cooperative bioleaching of sphalerite by two kinds of bacteria-Acidithiobacillus ferriooxidans and Acidithiobacillus thiooxidans. Trans. Nonferrous Met. Soc. China, 12: 190-195. [ Links ]
49 Zapata, M., Márquez, G., Ossa, M. 2007. Sulphur product layer in sphalerite biooxidation: Evidences for a mechanism of formation. Advances Materials Research, 20 (21): 134-138. [ Links ]

