










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.13 no.2 Bogotá July/Dec. 2011
ARTÍCULO CORTO
Efectividad de cepas rizobianas de frijol bajo diferentes regímenes de fósforo
Effectiveness of cowpea rhizobial strains under different phosphorus regimes
Juliana Mayz Figueroa1
1Bióloga, MSc., Ph.D. Universidad de Oriente, Núcleo de Monagas, Campus "Juanico", Laboratorio de Rizobiología, Maturín, estado Monagas, Venezuela..julianamayz@cantv.net; julianamayz@gmail.com
Resumen
En Venezuela, el frijol representa una alternativa a la proteína animal, debido a su alto consumo y valor nutritivo, por ello se ha estimulado la implementación de programas para reactivar la economía de los pequeños y medianos productores, a fin de incrementar su producción y así tener mayor disponibilidad de proteína de alta calidad a bajo costo; de manera que, los estudios encaminados a mejorar su cultivo, son acertados. Se evaluó la efectividad de cepas rizobianas de crecimiento lento (cl) y rápido (cr) en frijol (Vigna unguiculata (L.) Walp.) cultivar TC9-6 en varios regímenes de fósforo (0, 20, 40 y 80 kgP2O5 ha-1 ), con un diseño experimental de bloques al azar con arreglo factorial. Las plantas se cultivaron en 4 kg de suelo de sabana 45 días y las cepas en caldo de levadura y manitol: 5 (cr: JV91) y 10 (cl: JV94) días. La inoculación (2 ml cada vez) fue aplicada a la siembra y 6 días más tarde. La utilización de fósforo (40-80 kgP2O5 ha-1 ) incrementó la nodulación (número, peso seco total e individual de nódulos) y favoreció la aparición de nódulos rojos; así mismo, acrecentó el peso de la materia seca, la altura, el número de hojas y la concentración de nitrógeno del vástago. Los valores fueron similares con ambos tipos de cepas (efectividad similar) y para las dos concentraciones (40-80 kgP2O5 ha-1 ), con las menores estimaciones para 0 y 20 kgP2O5 ha-1 . De acuerdo con los resultados las concentraciones de 40 y 80 kgP2O5 ha-1 fueron las más favorables para el crecimiento y la nodulación de frijol.
Palabras clave: Vigna unguiculata, Rhizobium, crecimiento, nodulación, fósforo.
Abstract
In Venezuela, cowpea is an alternative to animal protein due to its high consumption and nutritious value, so it has stimulated the implementation of programs to reactivate the small and medium producers economy, in order to increase its production and to have major high quality protein availability at low cost; so that, the studies carry on to improve its cultivation, are well-aimed. The effectiveness of slow (sg) and fast (fg) growing rhizobial strains was evaluated in cowpea (Vigna unguiculata (L.) Walp) cultivar TC9-6 at various phosphorus regimes (0, 20, 40 and 80 kgP2O5 ha-1 ): randomized block design with factorial arrangement. Plants were cultivated in 4 kg savannah soil: 45 days, and the strains in yeast and mannitol broth: 5 (fg: JV91) and 10 (sg: JV94) days. The inoculation (2 ml each time) was applied at sowing time and 6 days later. Phosphorus utilization (40-80 kgP2O5 ha-1 ) increa- sed nodulation (nodule number, total and individual dry weight) and favoured nodule red colour appearance; also, incremented shoot dry matter weight, height, leaves number and nitrogen concentration. Values were similar with both strain types (similar effectiveness) and to the two doses (40-80 kgP2O5 ha-1 ), with lower estimations to 0 and 20 kgP2O5 ha-1 . Accordingly with the results, the doses of 40 and 80 kgP2O5 ha-1 were the most favourable to cowpea growth and nodulation.
Key words: Vigna unguiculata, Rhizobium, growth, nodulation, phosphorus.
Introducción
De acuerdo con Sauer (1952) y Summerfield et al. (1974), el frijol tuvo su centro de origen en Etiopía, desde donde se extendió hacia Asia y el Mediterráneo a través de Egipto y Arabia. En la actualidad es cultivado ampliamente en países tropicales y subtropicales, donde se le dan diversos usos (consumo humano de las vainas, las hojas y de los granos verdes o secos, consumo animal en pastizales, y como heno y ensilaje) y se hacen estudios para conocer su potencialidad para otros.
En Venezuela, el frijol es cultivado principalmente en pequeñas parcelas en las sabanas orientales para consumo de sus granos frescos o secos; la mayoría de los suelos presentan deficiencias de Ca, N y P, pH ácido y toxicidad de aluminio y/o manganeso (Mayz et al., 2003). Para los años 1999-2000, este ocupó el segundo lugar de producción y consumo humano después de la caraota (MAC, 2000; García et al., 2009), condición que hoy permanece. Debido a su alto valor nutritivo (alrededor de 25% de proteína cruda), el frijol representa una alternativa a la proteína animal, razón que ha estimulado la implementación de programas sociales para reactivar la economía de los pequeños y medianos productores, con el fin de incrementar su producción y tener una mayor disponibilidad de proteína de alta calidad a un menor costo que la proteína animal (PESA, 2002, Defang et al., 2008). Además, el frijol es una planta con una alta heredabilidad que ha permitido, a través de estrategias de cruzamiento tradicional, mejorar su rendimiento y calidad (Muchero et al., 2009; Xu et al., 2009).
En Venezuela, la fuente de proteína en la formulación de raciones para becerros y aves proviene, en buena parte, de la soya, pero esta es un producto básicamente importado y para el cual se predice una gran escasez debido a la expansión de la economía china y a la emergencia de los países asiáticos. El uso de fuentes alternativas de proteína con cultivos que pueden ser producidos en la finca ganadera, coloca al frijol como una leguminosa de gran potencial tanto alimenticio como económico. Esta es una leguminosa ampliamente conocida entre productores, precoz, con una producción alrededor de 1.200 kg de MS/ ha, cosechada como planta completa (Canelones and Castejón, 2006; Defang et al., 2008).
En este contexto, los estudios conducentes a establecer las mejores condiciones para el cultivo de frijol, a fin de incrementar su producción y superficie de siembra, son pertinentes. Así, este estudio contribuye al establecimiento de la mejor combinación de dosis de fósforo y de cepa rizobiana, para maximizar el crecimiento de frijol y la fijación de nitrógeno, con la finalidad de obtener un mejor rendimiento. Se evaluó el efecto de varias dosis de fósforo en combinación con cepas rizobianas de crecimiento lento y rápido sobre el crecimiento y nodulación de frijol.
Materiales y métodos
El experimento se llevó a cabo en invernadero con luz natural y temperatura diurna de 28 a 33 °C, en un suelo Ultisol virgen no estéril de la Estación Experimental de Sabana de la Universidad de Oriente en Jusepín, con clase textural areno francosa, bajo contenido de materia orgánica, nitrógeno, calcio, fósforo, magnesio y potasio y baja capacidad de intercambio catiónico (tabla 1 ). Se usaron cuatro tratamientos de fósforo (0, 20, 40 y 80 kgP2O5 ha-1 ) en combinación con cepas rizobianas de crecimiento lento (JV94) y rápido (JV91) bajo un diseño de bloques al azar con arreglo factorial. La selección de estas cepas obedece a su conducta simbiótica efectiva (Mayz et al., 2003) en el cultivar de frijol TC9-6, utilizado en este ensayo. Como fuente de fósforo se usó superfosfato triple (46% de P2O5 ), del cual se utilizaron 57,96; 115,93 y 231,88 mg por cada 4 kg de suelo de sabana, correspondientes respectivamente a las dosis de 20, 40 y 80 kgP2O5 ha-1 ; para estos cálculos se consideró el peso de una hectárea (3 x 106 kg) a una profundidad de 20 cm y de densidad 1,5g/cm3 .
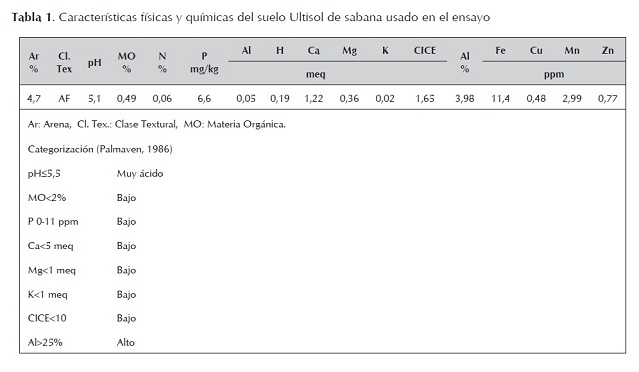
Las plantas de frijol (Vigna unguiculata (L.) Walp) cv. TC9-6 (Valladares, 1990) se cultivaron en materas plásticas (4 kg) durante 45 días y las cepas en caldo de extracto de levadura y manitol en un agitador orbital (60 rpm) por 5 (JV91) y 10 (JV94) días respectivamente. El número de células viables se calculó por relación de la absorbancia medida a 540 nm de los cultivos de las cepas con la curva de crecimiento o poblacional establecida en el Laboratorio de Rizobiología (UDO, Monagas, Venezuela), de acuerdo al procedimiento descrito por Somasegaran and Hoben (1994). Las semillas, seleccionadas por uniformidad de tamaño, fueron superficialmente desinfectadas con solución de cloruro de mercurio (0,2%), luego lavadas varias veces con agua destilada estéril y finalmente germinadas en cajas de Petri con papel toalla húmedo a temperatura ambiente (28-30 °C). Se sembraron cuatro semillas pregerminadas con radículas de 1,5-2 cm de largo, raleándose a dos por matera cuando las plantas tuvieron 4,5 cm de altura. La inoculación (2ml planta-1 ) se realizó con la suspen- sión bacteriana (1,09-1,18 x 109 células activas ml -1 ) de las cepas individuales al momento de la siembra y seis días más tarde.
En la cosecha, en las dos plantas por matera, se evaluaron: el peso seco, la altura, el número de hojas y la concentración de nitrógeno del vástago y la nodulación (número y peso seco total, peso individual y color de los nódulos: rojos o rosados, considerados como efectivos, y blancos, calificados como inefectivos). Los pesos secos se obtuvieron después del secado en estufa a 65 °C por 72 h y el nitrógeno se determinó por el método de Kjeldahl, después de la digestión ácida de la materia seca, para lo cual se usó un equipo Tecator, Kjeltec system. Los datos fueron analizados de acuerdo al diseño empleado y las medias comparadas por la diferencia mínima significativa (p≤ 0,05) usando el programa estadístico Statistix 8.0 (Analytical Software, 2003).
Resultados y discusión
El crecimiento de frijol fue afectado por las concentraciones de fósforo, pero no por las cepas aplicadas (figura 1). Los menores pesos secos del vástago se presentaron en las plantas cultivadas en ausencia de fósforo o con 20 kgP2O5 ha-1 (peso promedio: 1,13 g planta-1 ), incrementándose estos con las concentraciones de 40 y 80 kgP2O5 ha-1 (peso promedio: 1,87 g planta-1 ); los valores obtenidos en estos dos niveles son similares entre sí. La reducción en ausencia de fósforo o con la concentración de 20 kg P2O5 ha-1 fue de un 40%. Similarmente, el número de hojas y la altura de la parte aérea fueron mayores y estadísticamente diferentes a los valores obtenidos en ausencia de fósforo o con 20 kg P2O5 ha-1 (figuras 2s y 3). Se podría asumir que la reducción del crecimiento se debe en parte al menor número de hojas y altura de la planta. Los pa-rámetros de crecimiento medidos, peso seco, altura y número de hojas del vástago se correlacionaron significativamente con las concentraciones de fósforo, respectivamente 0,86; 0,86 y 0,88.
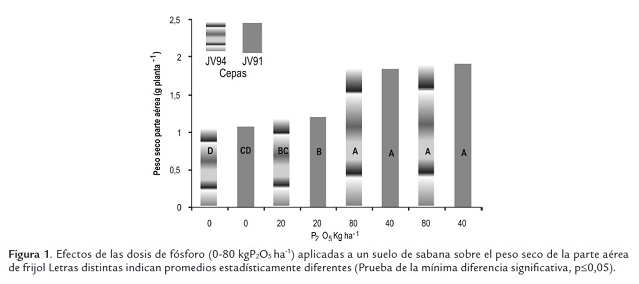
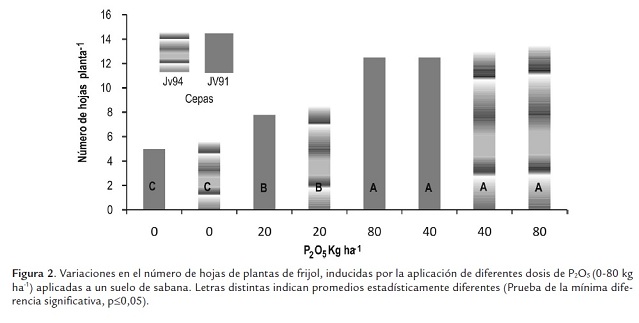
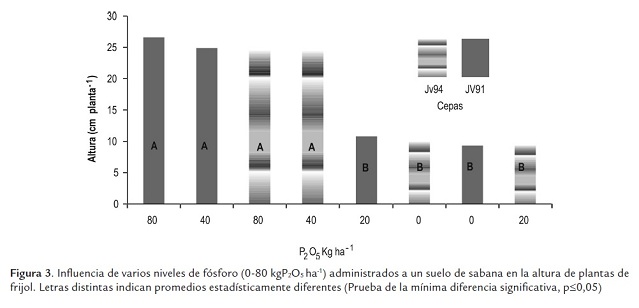
Después del nitrógeno, el fósforo es el elemento más importante asociado con la fijación de nitrógeno, resulta fundamental en diferentes etapas del desarrollo y crecimiento de las plantas, estimula el crecimiento a través de su participación en estructuras esenciales y en procesos metabólicos, por ejemplo forma parte del ADN, ATP y ADP, compuestos responsables del almacenamiento de energía, que permiten el transporte de nutrimentos a través de la pared celular y la síntesis de ácidos nucleícos y proteínas (Thapliyal et al., 2009). De manera específica se ha encontrado que la arquitectura radicular puede sufrir varios cambios primarios en caraota a consecuencia de la deficiencia de fósforo, tales como un incremento del crecimiento de las raíces laterales y de la ramificación de las raíces secundarias a expensas de la elongación de la raíz principal (Lynch and Brown, 2001).
Fujita et al. (2003) han señalado algunas consecuencias de la deficiencia de fósforo, entre estas están: reducción del pool de fósforo residual y de la tasa fotosintética, restricción de la división, elongación y expansión celular en los ápices meristemáticos y alteración de las relaciones hídricas y de la conductancia estomática. De tal manera que una deficiencia del elemento se manifiesta finalmente en una reducción del crecimiento, como se observa en este ensayo con los niveles de 0 y 20 kg ha-1 . Además se ha encontrado en Vigna aconitifolia cv. Mash-88, Vigna radiata cv. Moong-6601 y Glycine max cv. Tamahomare, retardo del inicio de la formación de las vainas (Chaudhary et al., 2008), en Vicia faba cv. Minor, Medicago polymorph y Arachis hypogaea, reducción del número y peso de las vainas por planta, y de la producción y peso de las semillas (Muir et al., 2001; Munir and Abdel-Rahman, 2002; Hossain et al., 2007). La adición de fósforo a suelos deficientes estimula el crecimiento radicular y fortalece el vástago, lo cual avala lo observado con las concentraciones de 40 y 80 kgP2O5 ha-1 : mayores valores de la materia seca, altura y número de hojas del vástago.
Los síntomas de deficiencia de fósforo se observaron en las plantas a las cuales no se les aplicó P2O5 y en esas tratadas con 20 kgP2O5 ha-1 , estos incluyen: plantas pequeñas con tallos y pecíolos delgados, transición de color en las hojas desde verde oscuro hacia verde oliva y luego amarillamiento, puntos violeta a lo largo o entre las venas, desarrollándose en áreas necróticas.
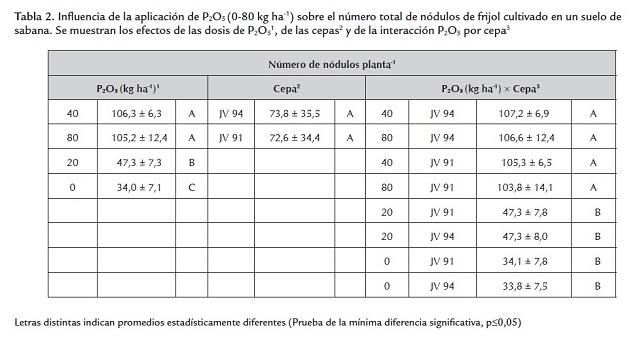
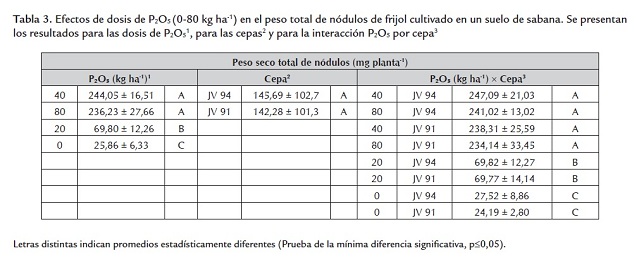
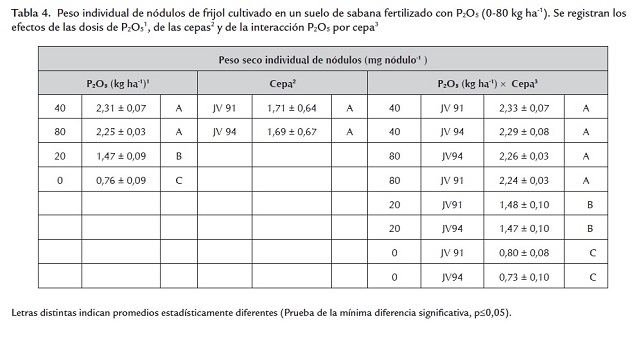
Al igual que el crecimiento, la nodulación (número, peso seco total e individual y color de los nódulos) fue influenciada por las dosis de P2O5 pero no por las cepas aplicadas. El menor número de nódulos se presentó en las plantas no fertilizadas con P o en esas a las cuales se les aplicaron 20 kg ha-1 , obteniéndose valores similares entre sí; mientras que el número fue máximo con 40 y 80 kg ha-1 (sin diferencias significativas entre ambas dosis) (tabla 2 ). La biomasa nodular total e individual fue mayor en las plantas fertilizadas con 40 y 80 Kg ha-1 , presentándose una severa disminución en los otros tratamientos (0 y 20 KgP2O5 ha-1 ) (tablas 3 y 4 ). Las dosis de fósforo y los parámetros de nodulación (número, peso total e individual de nódulos) presentaron correlaciones de 0,86; 0,86 y 0,85, respectivamente. El color de los nódulos presentes en los niveles de 40 y 80 kg ha-1 varió de rosado a rojo; en contraste, en las plantas no fertilizadas o abonadas con 20 kg ha-1 fueron de color blanco, lo cual aunado al menor número y peso total e individual de los nódulos en esos tratamientos podría indicar un efecto adverso de la deficiencia de fósforo en la iniciación, el crecimiento y el desarrollo nodular. Estos efectos se han notado en otras especies de leguminosas. Almeida et al. (2000) observaron en Trifolium repens L. cv. Milkanova que una severa deficiencia de fósforo inhibe en mayor extensión la nodulación y la fijación de nitrógeno que el crecimiento de la planta; Qiao et al. (2007) sugieren que la deficiencia de fósforo deteriora la fijación simbiótica del N2 a través del retraso en el desarrollo del nódulo y del inicio de la función nodular e Islam y Ali (2009) encontraron en Cicer arietinum un aumento del nitrógeno fijado con la aplicación de 80 kgP ha-1 .
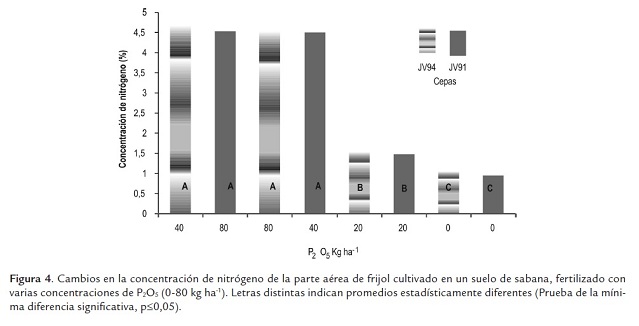
La concentración de nitrógeno varió con las concen- traciones de fósforo, pero no con las cepas aplicadas, incrementándose hasta 80 kgP2O5 ha-1 , sin diferencias significativas entre los valores obtenidos con 40 y 80 kgP2O5 ha-1 . Las menores concentraciones se presentaron con la dosis de 20 kgP2O5 ha-1 y en las plantas no fertilizadas; en ambos tratamientos fueron significativamente diferentes (figura 4). Estos menores valores son secuela de la deficiencia de nitrógeno por la falta del suministro nitrogenado a través de la fijación simbiótica como lo denotan las estimaciones de la nodulación para ambos tratamientos.
Conclusiones
Los resultados muestran que un enriquecimiento del suelo de sabana con 40 ó 80 kgP2O5 ha-1 mejora la conducta simbiótica de las cepas JV91 y JV94, que se traduce en un mayor número y peso total de los nódulos, mayor peso por nódulo y presencia de nódulos rojos o rosados, indicativos de actividad fijadora de nitrógeno. Este efecto favorecedor sobre la fijación de nitrógeno se tradujo en un mejor crecimiento de las plantas cultivadas con estas concentraciones, mostrado por un mayor peso seco de la parte aérea, número de hojas y altura de las plantas; de tal manera que se revela un efecto benéfico del fósforo tanto en el crecimiento como en la nodulación de frijol.
Referencias bibliográficas
1 Almeida, J., Hartwig, U., Frehner, M., Nösberger, J. and Lüscher A. 2000. Evidence that P deficiency induces N feedback regulation of symbiotic N2 fixation in white clover (Trifolium repens L.) Journal of Experimental Botany 51 (348): 1289-1297. [ Links ]
2 Analytical Software. 2003. Statistix 8.0 for Windows. Tallahassee, United States. [ Links ]
3 Canelones, C. and Castejón, M. 2006. Flours of whole cowpea (Vigna unguiculata) and cob maize (Zea mays) as supplements for suckling calves. Zootecnia Tropical. 24 (3): 361-378. [ Links ]
4 Chaudhary, M. I., Adu-Gyamfi, J. J., Saneoka, H., Nguyen, N. T., Suwa, R., Kanai, S., El-Shemy, H. A., Lightfoot, D. A. and Fujita, K. 2008 The effect of phosphorus deficiency on nutrient uptake, nitrogen fixation and photosynthetic rate in mashbean, mungbean and soybean. Acta Physiologiae Plantarum 30 (4): 537-544. [ Links ]
5 Defang, H., Teguia, A., Awah-Ndukum, J., Kenfack, A., Ngoula, F. and Metuge, F. 2008. Performance and carcass characteristics of broilers fed boiled cowpea (Vigna unguiculata L. Walp.) and or black common bean (Phaseolus vulgaris) meal diets. African Journal of Biotechnology 7 (9): 1351-1356. [ Links ]
6 Fujita, K., Okada, M., Lei, K., Ito, J., Ohkura, K., Adu-Gyamfi, J. J. and Mohapatra, P. K. 2003. Effect of P-deficiency on photoassimilate partitioning and rhythmic changes in fruit and stem diameter of tomato (Lycopersicon esculentum) during fruit growth. Journal of Experimental Botany 54 (392): 2519-2528. [ Links ]
7 García, O., Infante, R. y Rivera, C. 2009 Las leguminosas, una fuente importante de fibra alimentaria: Una visión en Venezuela. Instituto Nacional de Higiene Rafael Rangel Aldao. 40 (1): 57-63. [ Links ]
8 Hossain, M. A., Hamid, A. and Nasreen, S. 2007. Effect of nitrogen and phosphorus fertilizer on N/P uptake and yield performance of groundnut (Arachis hypogaea L.). Journal of Agricultural Research (Lahore) 45 (2): 119-127. [ Links ]
9 Islam, M. y Ali, M. S. 2009. Efecto de la aplicación de azufre-fósforo sobre la fijación de nitrógeno y la captación de nutrientes en garbanzo (Cicer arietinum L.). Agrociencia 43: 815-826. [ Links ]
10 Lynch, J. P. and Brown, K. M. 2001. Topsoil foraging: an architectural adaptation of plants to low phosphorus availability. Plant and Soil 237: 225-237. [ Links ]
11 Mayz, J., Campos, F. and Valladares, N. 2003. Evaluation of rhizobial (Rhizobium sp.) isolates for their effects on cowpea [Vigna unguiculata (L.) Walp.] growth and yield. Tropical Agriculture 80: 36-40. [ Links ]
12 Ministerio de Agricultura y Cría (MAC). 2000. Rendimiento de productos vegetales. Dirección de Estadísticas e Información. Ministerio de Agricultura y Cría. Caracas, Venezuela. [ Links ]
13 < Muchero, W., Diop, N. N., Bhat, P. R., Fenton, R. D., Wanamaker, S., Pottorff, M., Hearne, S., Cisse, N., Fatokun, C., Ehlers, J. D., Roberts, P. A. and Close T. J. 2009. Consensus genetic map of cowpea [Vigna unguiculata (L) Walp.] and synteny based on EST-derived SNPs. Proceedings of the National Academy of Sciences of the United States of America 106 (43): 18159-18164. /p> [ Links ]
14 Muir, J. P., Pitman, W. D., Coombs, D. F. 2001. Seeding rate, phosphorus fertilization, and location effects on ´Armadillo´ Burr Medic. Agronomy Journal 93: 1269-1275. [ Links ]
15 Munir, T. and Abdel-Rahman, T. 2002. Impact of seeding rate, seeding date, rate and method of phosphorus application in faba bean (i<>Vicia faba L. minor) in the absence of moisture stress. Biotechnologie, Agronomie Société et Environnement 6 (3): 171-178. [ Links ]
16 Palmaven S. A. 1986. Análisis de suelo y su interpretación. Serie B. Información Técnica 40 p. [ Links ]
17 Programa Estratégico de Seguridad Alimentaria (PESA). 2002. Informe de comienzo proyecto UTF/Ven/008/Ven julio-octubre 2002. Caracas, Venezuela. 25 p. [ Links ]
18 Qiao, Y., Tang, C., Han, X. and Miao, S. 2007. Phosphorus deficiency delays the onset of nodule function in soybean. Journal of Plant Nutrition 30 (9): 1341-1353. [ Links ]
19Sauer, C. O. 1952. Agricultural origins and dispersals. Bowman Memorial Lectures, Series 2. New York: American Geographical Society. 110 p. [ Links ]
20 Somasegaran, P. and Hoben, H. J. 1994. Handbook for rhizobia. Methods in legume Rhizobium technology. Berlín, Germany: Springer-Verlag. 456 p. [ Links ]
21 Summerfield, R.J., Huxley, P.A. and Steele, W. M. 1974. cowpea (Vigna unguiculata (L.) Walp.). Field Crop Abstracts. 27: 301- 312. [ Links ]
22 Thapliyal, R., Chaukiyal, S. P. and Pokhriyal, T. C. 2009. Effect of nitrogen and phosphorus doses on the nitrogen fixation activity of Albizia lebbeck plants in the nursery. NFT News 11 (1): 1-4. [ Links ]
23 Valladares, N. 1990. Nuevas líneas experimentales de frijol (Vigna unguiculata (L.) Walp) para la ecología de sabana, desarrolladas por selección individual de la población original "Tejero Criollo". Premio Agropecuario Banco Consolidado. Caracas, Venezuela. 115 p. [ Links ]
24 Xu, N. W., Xu, S. and Ehlers, J. 2009. Estimating the broad-sense heritability of early growth of cowpea. International Journal of Plant Genomics 2009: 1-4. [ Links ]

