










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.13 no.2 Bogotá July/Dec. 2011
ARTÍCULO CORTO
Organogénesis directa in vitro a partir de hojas de la planta Antiplasmodial Solanum nudum Dunal
Direct organogenesis in vitro from leaves of Solanum nudum Dunal - Antiplasmodial plant
Oscar Manuel Suárez Cardoso1 , Esther Julia Naranjo Gómez2 , Lucía Atehortúa Garcés3 , Silvia Blair Trujillo4
1Biólogo. Grupo de biotecnología Vegetal. Universidad de Antioquia. Calle 70 No. 52 - 21, A.A. 1226. Medellín, Colombia. oscarsuarez16@ gmail.com
2M.Sc. Grupo de biotecnología Vegetal. Universidad de Antioquia. Calle 70 No. 52 - 21, A.A. 1226. Medellín, Colombia. estherjulia@gmail.com
3PhD. Grupo de biotecnología Vegetal. Universidad de Antioquia. Calle 70 No. 52 - 21, A.A. 1226. Medellín, Colombia. latehor@gmail.com
4M.Sc. Grupo de Malaria. Sede de Investigación Universitaria. Universidad de Antioquia. Calle 62 No. 52-59. Torre 1. Laboratorio 610, Sede de Investigación Universitaria, Telefax: 219 64 87Medellín, Antioquia. silviablair@gmail.com
Recibido: agosto 09 de 2011 Aprobado: noviembre 15 de 2011
Resumen
Solanum nudum Dunal (Solanaceae), es una especie vegetal con potencial para desarrollar un tratamiento quimioterapéutico contra la malaria. Este es el primer reporte de un protocolo rápido, eficiente y reproducible de organogénesis directa a partir de segmentos de hoja de plántulas in vitro de esta especie.
Los segmentos de hojas de plántulas de 5 meses de germinadas fueron cultivados sobre medio Murashige y Skoog (MS) a mitad de sales y vitaminas, con diferentes concentraciones de Bencilaminopurina (BAP), en combinación con Acido Indolacético (AIA). Se evaluó también el efecto de la iluminación en periodos 0/45, 15/30 y 30/15 días oscuridad/ luz, sobre la inducción de brotes. Se registró un promedio alto de formación de brotes (4,83) en explantes cultivados en medio suplementado con 2,0 mg/L de BAP y 0,1 mg/L de AIA, bajo condición de iluminación por un periodo de 30/15 días oscuridad/luz. Luego de la inducción, los brotes obtenidos fueron transferidos a medio MS suplementado con 0,3 mg/L de Giberelina (GA3), y mantenidos en condiciones de luz donde también enraizaron. Las plántulas regeneradas se llevaron a condiciones de invernadero y fueron morfológicamente similares a las plantas madres.
Palabras clave: Solanaceae, Solanum nudum, Planta antimalárica, micropropagación, inducción de brotes.
Abstract
Solanum nudum Dunal (Solanaceae) is a plant with a potential for developing chemotherapeutic treatments against malaria. This is the first report of a fast, efficient, and reproducible direct organogenesis protocol from leave segments from in vitro seed-grown plantlets.
Leaves segments from 5 months old germinated plantlets were placed on half concentration Murashige and Skoog medium (MS), supplemented with several concentrations of Bencilaminopurin (BAP) combinated with Indolacetic Acid (IAA). Dark/ light incubation effect in periods 0/45, 15/30 and 30/15 dark/light days were evaluated on the buds induction. High frequency buds formation was shown (4,83) in explants cultured on MS supplemented with BAP 2,0 mg/L and AIA 0,1 mg/L under a period of 30 days of dark condition incubation. After induction, buds obtained were transferred to MS medium supplemented with gibberellic acid (GA3) 0,3 mg/L and maintained under artificial cool light, there the plantlets rooted. Regenerated plantlets were placed under greenhouse conditions and these were morphologically similar to donor plants.
Key words: Solanaceae, Solanum nudum, Antimalarial plant, micropropagation, buds induction.
Introducción
La planta Solanum nudum Dunal, pertenece a la familia Solanaceae y se encuentra distribuida en Centroamérica y Suramérica, desde México hasta Ecuador y Venezuela, y en las Antillas Mayores (Tropicos.org, 2011). En Colombia, la especie es conocida por las comunidades del Pacífico como “Saúco y Zapata”. Esta especie es de hábito arbóreo con una altura aproximada de 3 metros, hojas alternas, elípticas, margen entera, base atenuada, ápice acuminado y envés con pelos en las bifurcaciones de la vena principal. Su fruto es una baya globosa, con muchas semillas planas. Las hojas de esta planta son utilizadas por las comunidades del Pacífico colombiano en la medicina tradicional para el tratamiento de fiebres, dolores de cabeza, escalofríos, inflamaciones del hígado, entre otros (Blair y Madrigal, 2005).
En condiciones de laboratorio, se han determinado distintos compuestos esteroidales presentes en los extractos de sus hojas, que han mostrado actividad in vitro sobre cepas de Plasmodium falciparum resistentes a Cloroquina (Sáez et al., 1998), y un alto efecto terapéutico en el tratamiento in vivo de ratones infectados con P. berghei, (Echeverri et al., 2001). Adicionalmente, se determinó que ninguno de sus extractos tiene propiedades mutagénicas (Pabón et al., 2003) y causan inhibición del crecimiento de cepas de P. falciparum debido a la presencia de compuestos esteroidales obtenidos de sus hojas (Pabón et al., 2002) y se ha comprobado la actividad antiplasmódica de los extractos en trofozoitos hepáticos de P. vivax (Londoño et al., 2006).
Las poblaciones naturales, de especies de plantas medicinales, históricamente han afrontado problemas que las han llevado a diferentes grados de peligro de extinción, como la sobreexplotación, debido a la dificultad de la síntesis química de los metabolitos secundarios y las consecuencias del cambio climático, entre otras (Olliaro y Yuthavong, 1999; Caldentey y Barz, 2003; Nalawade y Tsay, 2004; Bourdy et al., 2008). Las herramientas biotecnológicas ofrecen una alternativa para superar algunos de estos problemas mediante el desarrollo de sistemas de micropropagación in vitro que garanticen la conservación y la producción de materia prima básica para la industria (Rout, 2005).
Las plantas medicinales propagadas in vitro conforman una fuente inmediata de material vegetal uniforme y libre de contaminación microbiana y química para la caracterización bioquímica y la identificación de compuestos activos (Paniego y Giulietti, 1998). La micropropagación vegetal ha sido utilizada exitosamente para la conservación de diferentes especies medicinales, cuyas poblaciones han llegado a estar en peligro de extinción (Martin et al., 2003; Dalal y Ravishankar, 2004; Sudha et al., 2005), además para la extracción de metabolitos secundarios para el desarrollo de medicamentos antimaláricos, entre otros (Liu et al., 2003).
Considerando que la población de S. nudum de Tumaco (Nariño, Colombia) es reportada por los curanderos como antimalárica (Blair y Madrigal, 2005) y ha sido comprobada su promisoria actividad antiplasmodial (Sáez et al., 1998; Echeverri et al., 2001; Pabón et al., 2002; Pabón et al., 2003; Londoño et al., 2006), es importante encontrar vías para la multiplicación de estos genotipos con fines de extracción de sus metabolitos de interés y conservación ex situ, como herramienta para evitar la pérdida cultural del conocimiento etnomedicinal y la sobreexplotación del recurso natural. Por lo anterior, el presente trabajo tuvo como objetivo el desarrollo de un protocolo de micropropagación para la especie S. nudum.
Materiales y métodos
Fuente de material vegetal y protocolo de desinfección
Se colectaron frutos de Solanum nudum Dunal, en el corregimiento de Robles, vereda Inguapí del Guadual (Latitud: 1,7° 42’7’’; Longitud: -78,6° 41’46’’), municipio de Tumaco, departamento de Nariño, Colombia. La colecta fue realizada en el mes de octubre de 2008 y el material vegetal fue llevado al Laboratorio de Biotecnología Vegetal de la Universidad de Antioquia. Las semillas fueron extraídas y desinfectadas mediante lavado con jabón quirúrgico (Quirucidal®), seguido por una inmersión en fungicida (Colimyl®) 2g/L por 2 horas y luego en hipoclorito de sodio (NaOCl 0,25% del volumen en agua destilada). Después de cada paso las semillas fueron enjuagadas 3 veces en agua destilada estéril. Las semillas desinfectadas fueron finalmente llevadas a medio de cultivo. La manipulación del material vegetal fue realizada en cámara de flujo laminar horizontal.
Condiciones de cultivo
Todos los medios de cultivo, empleados para el desarrollo del experimento, fueron preparados con agua destilada y homogenizados en una plancha de calentamiento con agitación magnética. El pH inicial fue ajustado a 5.75 en un pH-metro (Metrohm), con soluciones de NaOH y HCl 1N y gelificado con 2,0 g/L de Gelrite (Phytotech). La esterilización de los medios de cultivo se realizó en autoclave a 121 °C y 15 psi durante 20 minutos. Las condiciones lumínicas del laboratorio fueron de aproximadamente 800 lux.
Germinación in vitro
Las semillas fueron puestas a germinar en cajas de Petri que contenían el medio sugerido por Sumita y Timir (1989): sales y vitaminas de Murashige y Skoog (MS) (Murashige y Skoog, 1962) a mitad de su concentración, suplementado con la hormona Giberelina (GA3), a una concentración de 10mg/L y sucrosa 20 g/L. Las cajas Petri fueron ubicadas en oscuridad por un periodo de 7 días, una vez iniciada la germinación fueron llevadas a condiciones de luz.
Las plántulas obtenidas fueron subcultivadas en recipientes de vidrio con una capacidad de 100 ml y 56 mm de diámetro, que contenían 20 ml de medio MS a la mitad de su concentración en sales y vitaminas, suplementado con 1,0 g/L de carbón activado, y mantenidas bajo condiciones de iluminación, con el fin de asegurar material totalmente aséptico para el proceso de micropropagación clonal masiva.
Inducción de brotes adventicios
Se tomaron como explantes segmentos de hojas de plántulas de 150 días de germinadas (figura 2A). Los explantes fueron ubicados con su superficie adaxial sobre medio sólido MS suplementado con las hormonas ácido indolacético (AIA) 0,1mg/L y en diferentes concentraciones de Bencil aminopurina (BAP) (0,5; 1,0; 2,0 mg/L); un medio libre de hormonas fue tomado como control.
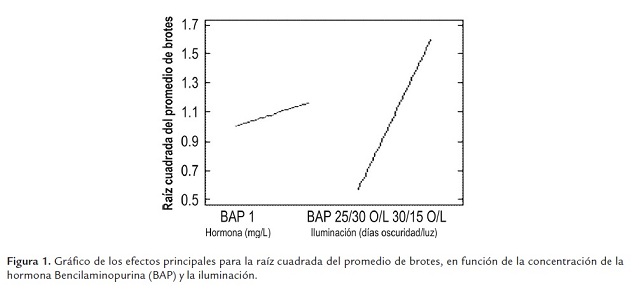

Para evaluar el proceso de inducción de brotes, además del factor de la concentración hormonal, se incluyó el efecto de las condiciones de iluminación en periodos (días) de oscuridad/luz en tres niveles (15/30; 30/15 y 0/45= control). Los brotes fueron transferidos a medio MS suplementado con 0,3 mg/L de GA3 para su desarrollo. Cuando las plántulas superaron los 4 cm de altura y formaron raíces bien desarrolladas fueron transferidas a macetas plásticas cubiertas con polipropileno, bajo iluminación artificial de 24 h, humedad relativa de 80% y mantenidas con riego cada 3 días.
Diseño estadístico
Se empleó el programa Statgraphics Centurion para la aleatorización de los tratamientos y el análisis de los datos, se verificaron los supuestos básicos del ANOVA a un nivel de confianza del 95%, valores p<0.05 fueron considerados estadísticamente significativos.
Se utilizó un diseño factorial 4x3, es decir, con 4 niveles del factor hormona BAP mg/L (0,5; 1,0; 2,0 y control) y 3 niveles del factor iluminación días oscuridad/ luz (15/30, 30/15 y control). Se hicieron 4 réplicas por tratamiento con 5 repeticiones cada una. Con el fin de cumplir los supuestos de normalidad y varianza constante (Gutiérrez y de la Vara, 2004), se usó como variable independiente la raíz cuadrada del promedio de los brotes por tratamiento.
Resultados y discusión
Proceso de desinfección y germinación in vitro
El proceso de desinfección aplicado a las semillas fue eficiente y ningún tipo de contaminación fue observada. Las semillas germinaron entre los días 7 y 9 después de la siembra en el medio de cultivo suplementado con GA3, con un porcentaje de germinación del 100%; las semillas en medios libres de hormonas y en campo presentaron tasas bajas de germinación. El alto porcentaje de germinación obtenido puede ser atribuido al efecto de la adición de Giberelinas (GA3) en el medio de cultivo. Estas hormonas son reconocidas por Taiz y Zeiger (2006) como inductoras de la germinación, que al ser aplicadas a semillas de plantas silvestres pueden complementar los requerimientos (luz y temperatura) para superar la posible dormancia que estas puedan llegar a tener.
Inducción de brotes adventicios
Una observación preliminar de los datos condujo a la eliminación de los controles y del nivel 0,5 del factor hormona del diseño estadístico propuesto, pues el número de brotes obtenidos con esos niveles siempre fue cero (Gutiérrez y de la Vara, 2004). Por lo tanto, el diseño final fue un factorial 2x2, completamente aleatorizado. Dado que en el análisis de varianza (ANOVA) no se evidenció un efecto significativo de la interacción entre los factores, se eliminó del análisis para evitar el error que esta pudiera aportar al diseño.
La hormona BAP en las concentraciones evaluadas no mostró un efecto estadísticamente significativo para la inducción de brotes adventicios (valor p=0,7422); sin embargo, se observó una mejor respuesta proporcional a la concentración de fitohormona: a mayor concentración de hormona (2,0 mg/L) se encuentra una mayor respuesta inductora de brotes organogénicos adventicios (figura 1), bajo las mismas condiciones de iluminación. Tivarekar y Eapen (2001), reportaron un comportamiento similar en la inducción de brotes sobre cotiledones inmaduros de fríjol mungo (Vigna radiata): a mayor concentración de la citoquinina encontraron una mejor respuesta inductora de brotes, con los mismos reguladores de crecimiento usados en este estudio. Se ha reportado que la formación de estos brotes se debe a cambios en células del parénquima que son distribuidas hacia los alrededores de los haces vasculares, las cuales son las señales que dirigen el destino de las células hacia la formación de agregados celulares o meristemáticos inducidos por la adición de diferentes concentraciones de la hormona BAP al medio (Salazar et al, 2005).
El factor iluminación en días oscuridad/luz mostró un efecto estadísticamente significativo (valor p=0,0413) dentro de los niveles evaluados. Los tratamientos de incubación en oscuridad, seguidos de un periodo de incubación en luz, han sido reportados como benéficos en la inducción de brotes masivos en diferentes especies y tipos de explantes, entre los que se incluyen: segmentos de hojas de Maracuyá (Passiflora edulis Sims) (Appezzato et al, 1999), dos variedades de la especie Diospyros kaki (Choi et al, 2001), Alstroemeria cv. ‘Yellow King’ (Pedraza et al, 2006), Violeta Africana (Saintpaulia ionantha Wendl.) (Al- Hussein et al, 2006), Durazno (Prunus domestica L.) (Petri y Scorza, 2010). Aunque no se conoce el mecanismo de cómo la incubación en la oscuridad estimula la respuesta organogénica, se cree que puede preservar y facilitar el movimiento de reguladores de crecimiento a sitios de regeneración (Compton, 1999), además de influenciar los niveles endógenos de la hormona AIA (López - Carbonell et al, 2000, citados por Petri y Scorza, 2010). En esta investigación se comprobó que un periodo de incubación oscuridad/luz es necesario para la inducción de organogénesis directa en la especie Solanum nudum Dunal.
La formación de brotes adventicios fue observada en los extremos de corte y en la vena media, siendo más numerosos en los primeros (figura 2B y C). Aunque se observó la formación de callo en pequeñas zonas localizadas en los extremos de los explantes, estas no se encontraban relacionadas con los brotes formados, la superficie de callo no superaba el 3% del total del explante y no se evidenció crecimiento, ni respuesta organogénica en dichas zonas, las cuales finalmente se necrosaron.
Los tratamientos control, establecidos para cada factor, no mostraron ninguna respuesta en la inducción de brotes. La mayoría de los explantes murieron antes de finalizar el periodo de experimentación. Lo anterior sugiere que la aplicación al medio de cultivo de una combinación citoquinina/auxina exógenas es necesaria para la inducción de brotes adventicios en segmentos de hoja de la especie Solanum nudum. Kaminek et al. citados por Khanam et al. (2000), reportaron que los niveles de citoquininas en células vegetales y su efecto inductor de respuesta organogénica dependen de la biosíntesis y de la absorción a partir de fuentes extracelulares, además de una posible influencia de las auxinas.
Luego de inducidos, los brotes fueron llevados a medio suplementado con Giberelina 0,3 mg/L (GA3) donde se observó un desarrollo normal, las plántulas alcanzaron hasta 4,0 cm de altura y raíces bien desarrolladas, que permitieron su fácil transferencia a condiciones de invernadero, donde alcanzaron su crecimiento y desarrollo en forma normal. Estas plantas presentaron características morfológicas similares con el material de campo (figura 2D), y análisis químicos cualitativos mostraron presencia de algunos de los metabolitos de interés (resultados no mostrados).
Previa a esta investigación, el grupo de Biotecnología Vegetal de la Universidad de Antioquia realizó un trabajo para el establecimiento a partir de la siembra de yemas apicales y axilares, en el cual no se logró la propagación de la especie, debido a la intensa formación de callo (Molina, 2006). De acuerdo con la literatura consultada, este es el primer reporte de un protocolo de propagación masiva in vitro de la especie Solanum nudum Dunal.
Conclusiones
Este trabajo describe un protocolo de organogénesis directa de Solanum nudum, a partir de segmentos de hoja, para la inducción de brotes organogénicos. Este es importante porque garantiza la estabilidad fenotípica y fitoquímica (datos no mostrados) del material reproducido, establece una fuente constante de material para la conservación ex situ que puede apoyar una estrategia de domesticación para la explotación sostenible de esta especie con potencial farmacéutico, lo que a su vez minimiza la presión sobre el recurso natural y contribuye a la conservación de la diversidad de especies vegetales de la región del Pacífico colombiano y la preservación de la cultura, especialmente la etnomedicina de las poblaciones que habitan esta región. Adicionalmente, se puede convertir en un modelo para estudios de tipo fisiológico y molecular, además de constituir una fuente de nuevas características agronómicamente deseadas para los cultivos comerciales de Solanáceas o puede constituir un complemento a herramientas biotecnológicas de mejoramiento como la transformación genética.
Agradecimientos
Los autores agradecen al Dr. Carlos Fernández, por la asesoría en el diseño estadístico; al Biólogo Felipe Cardona, del Herbario de la Universidad de Antioquia, por la recolección del material vegetal. La financiación de este proyecto se hizo con dineros del Ministerio de Agricultura de Colombia, Proyecto 2007V755-29, aprobado al Grupo malaria de la Universidad de Antioquia, y con dineros del Programa de Estrategia de Sostenibilidad 2010, Universidad de Antioquia.
Referencias bibliográficas
1 Al-Hussein, S., Shibl, R. and Karam, N. S. 2006. Regeneration in African Violet (Saintpaulia ionantha Wendl.) using Different Leaf Explants, Cytokinins Sources, and Light Regimes. Jordan Journal of Agricultural Sciences 2 (4): 361-370. [ Links ]
2 Appezzato, B., Carnero, M. L. and Carnier, M. 1999. Anatomical studies of in vitro organogenesis induced in leaf-derived explants of passionfruit. Pesquisa Agropecuaria Brasileira 34 (11): 2007-2013. [ Links ]
3 Blair, S. y Madrigal, B. 2005. Plantas antimaláricas de Tumaco, Costa Pacífica colombiana. Medellín, Colombia: Ed. Universidad de Antioquia pp. 266-270. [ Links ]
4 Bourdy, G., Willcox, M., Ginsburg, H., Rasoanaivo, P., Graz, B. and Deharo, E. 2008. Ethnopharmacology and malaria: New hypothetical leads or old efficient antimalarials? International Journal for Parasitology 38 (1): 33-41. [ Links ]
5 Caldentey, K. and Barz, W. 2003. Plant Biotechnology and Transgenic Plants: Plant derived drugs and extracts. New York, Estados unidos : Ed. Marcel Dekker Inc. p. 720. [ Links ]
6 Choi, J. Y., Kim, H. J., Lee, C. H., Bae J. M., Chung, S., Shin, J. S., and Hyung N. I. 2001. Efficient and simple plant regeneration via organogenesis from leaf segment cultures of persimmon (Diospyros kaki Thunb.). In vitro cellular and developmental biology- Plant 37 (2): 274-279. [ Links ]
7 Compton, M. 1999. Dark treatment improves adventitious shoot organogenesis from cotyledons of diploid watermelon. Plant Cell Tissue and Organ Culture 58 (3): 185-188. [ Links ]
8 Dalal, N. and Ravishankar, R. 2004. In vitro propagation of Oroxylum indicum Vent. a medicinally important forest tree. Japanese Forestry Society 9 (1): 61-65. [ Links ]
9 Echeverri, M., Blair, S., Carmona, J. and Pérez, P. 2001. Effect of Solanum nudum extracts on the liver infected with Plasmodium berghei. American Journal of Chinese Medicine 29 (3-4): 477-484. [ Links ]
10 Gutiérrez, P., y De la Vara S., R. 2004. Análisis y diseño de experimentos. México, México: Ed. McGraw-Hill Interamericana, pp. 238-239. [ Links ]
11 Khanam, N., Khoo, C. and Khan, A. 2000. Effects of cytokinin/ auxin combinations on organogenesis, shoot regeneration and tropane alkaloid production in Duboisia myoporoides. Plan Cell Tissue and Organ Culture 62 (2): 125-133. [ Links ]
12 Liu, Z., Guo, C., Wang, Y. and Ouyang, F. 2003. Comparison of various bioreactors on growth and artemisinin biosynthesis of Artemisia annua L. shoot cultures. Process Biochemistry 39 (1): 45-49. [ Links ]
13 Londoño, B., Arango, E., Zapata, C., Herrera, S., Sáez, J., Blair, S. and Carmona, J. 2006. Effect of Solanum nudum Dunal. (Solanaceae) steroids on hepatic trophozoites of Plasmodium vivax. Phytotherapy Research 20 (4): 267-273. [ Links ]
14 Martin, G., Geetha, S., Raja, S., Raghu, A., Balachandran, I. and Ravindran, P. 2003. An efficient micropropagation system for Celastrus paniculatus Wild.: a vulnerable medicinal plant. Japanese Forestry Society 11 (6): 267-273. [ Links ]
15 Molina, Y. 2006. Establecimiento del cultivo in vitro de la planta antimalárica Solanum nudum Dunal. Ibagué, Colombia: Universidad del Tolima, p. 103. [ Links ]
16 Murashige, T. and Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Plant Physiology 15 (3): 473-479. [ Links ]
17 Nalawade, S. and Tsay, H. 2004. In vitro propagation of some important chinese medicinal plants and their sustainable usage. In Vitro Cellular and Developmental Biology 40 (1): 143-154. [ Links ]
18 Olliaro, P. and Yuthavong, Y. 1999. An overview of chemotherapeutic targets for antimalarial drugs discovery. Pharmacology Therapy 81 (2): 91-110. [ Links ]
19 Pabón, A., Blair, S., Carmona, J., Maestre, A. and Camargo, M. 2002. Inhibition of Plasmodium falciparum by steroids isolated from Solanum nudum. Phytotherapy Research 16 (1): 59- 62. [ Links ]
20 Pabón, A., Blair, S., Carmona, J., Zuleta, M. and Sáez, J. 2003. Evaluation of mutagenicity of antimalarial products isolated from Solanum nudum (Solanaceae). Pharmazie 58 (4): 263-267. [ Links ]
21 Paniego, N. and Giulietti, A. 1998. Artemisinin production by Artemisia annua L- transformed organs cultures. Enzyme and Microbial Technology 18 (7): 526-530. [ Links ]
22 Pedraza, M., López, M., González, V., Engleman, E. and Sánchez, P. 2006. In vitro regeneration of Alstroemeria cv. ´Yellow King´ by direct organogenesis. Plan Cell Tissue and Organ Culture 84 (2): 189-198. [ Links ]
23 Petri, C., and Scorza, R. 2010. Factors affecting adventitious regeneration from in vitro leaf explants of ´Improved French´ plum, the most important dried plum cultivar in the USA. Annals of Applied Biology 156 (1): 79-89. [ Links ]

