










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.14 no.1 Bogotá Jan./June 2012
ARTÍCULO DE INVESTIGACIÓN
Potencial probiótico de cepas nativas para uso como aditivos en la alimentación avícola
Probiotic potential of native strains, for as feed additives for poultry
Cecilia Lara Mantilla1 y Ángela Burgos Portacio 2.
1Química; MSc; Ph D. Línea de Investigación en Biotecnología. Directora e investigadora GRUBIODEQ. Investigadora principal proyecto. Directora Tesis Maestría. Universidad de Córdoba. lara_mantilla_cecilia@hotmail.com; clara@sinu.unicordoba.edu.co
2 Microbiologa Industrial. Magister en Biotecnología. Investigadora GRUBIODEQ. Universidad de Córdoba.
Recibido: marzo 14 de 2011 Aprobado: junio 04 de 2012
Resumen
Se evaluó in vitro el potencial probiótico de cepas nativas aisladas de las heces de pollos asilvestrados (Gallus gallus) pertenecientes a los géneros Lactobacillus sp, Bacillus sp y levaduras tipo saccharomyces sp; se determinó la actividad probiótica mediante pruebas de resistencia al ácido (pH 3, 4, 5, 6, 7), sales de bilis (0,05, 0,1, 0,15, 0.3 %), tolerancia al NaCl (2, 4, 7, 10 %), actividad antagónica (Salmonella sp, E. coli), determinación del tipo de fermentación, crecimiento a temperaturas (28,37,43°C) y capacidad de crecimiento. Las cepas con mayor tolerancia se identificaron a través de pruebas bioquímicas y fermentación de carbohidratos. Como resultado se observó que tres microorganismos: Saccharomyces sp. (3), Bacillus sp. (7) y Lactobacillus sp. (14) poseen propiedades probióticas.
Palabras clave: Gallus gallus, Saccharomyces sp., Bacillus sp., Lactobacillus sp., aditivos microbianos.
Abstract
Was evaluated in vitro the probiotic potential of native strains isolated from feces of wild chickens (Gallus gallus) belonging to the genera Lactobacillus sp., Bacillus sp. and Saccharomyces ; probiotic activity was determined by testing acid resistance (pH 3, 4, 5.6, 7), bile salts (0,05, 0,1, 0,15, 0,3%), tolerance to NaCl (2, 4, 7,10%), antagonistic activity (Salmonella spp, E. coli), production gas (glucose), growth temperatures (28, 37, 43 ° C) and growth capacity. The most tolerant strains were identified by biochemical tests and carbohydrate fermentation. As a result it was found that three microorganisms: Saccharomyces sp. (3), Bacillus sp. (7) and Lactobacillus sp. (14) have probiotic properties.
Key words: Gallus gallus, Saccharomyces sp., Bacillus sp., Lactobacillus sp., microbial additives.
Introducción
En condiciones silvestres las aves como el Gallus gallus, poseen un tracto gastrointestinal en el que reside una gran variedad de microorganismos; de tal forma que los pollitos eclosionados obtienen las primeras bacterias de la boca, buche, excrementos de la madre y del medio ambiente; sin embargo en las instalaciones avícolas los pollitos eclosionan en un medio casi estéril y son ubicados en sus primeras semanas de vida en galpones con condiciones higiénicas estables, que imposibilitan la colonización de una microbiota intestinal beneficiosa; además de esto, los pollos están sometidos a situaciones que les producen estrés como son: la alta densidad de población, vacunación, altas o bajas temperaturas, humedad inadecuada, incidencia de gases tóxicos, inmunodepresión; que ocasionan desbalances de la microbiota intestinal en caso de que esta exista (Rondón et al., 2008; Lu et al., 2003). De esta manera las aves se encuentran expuestas a la incidencia de microorganismos patógenos que pueden causar enfermedades gastrointestinales como la salmonelosis y la colibacilosis (Barnes et al., 2003; La Ragione et al., 2002). Para contrarrestar los efectos de dichas enfermedades se utilizan los antibióticos de amplio espectro; pero estos compuestos han originado graves problemas de resistencia microbiana y efectos residuales colocando en riesgo la inocuidad alimentaria. Por otro lado se aumentan los costos del modelo de producción y una notable disminución en los índices productivos de la industria avícola.
Por lo anteriormente expuesto se plantea la utilización de probióticos como coadyuvantes dietéticos de origen microbiano que benefician la fisiología del hospedante al modular la inmunidad de la mucosa y la inmunidad sistémica, así como mejorar el balance nutricional y microbiano en el tracto gastrointestinal (Tannock, 1999); en la producción avícola, se reportan resultados sobre el uso de estos aditivos microbianos que como en la mayoría de los casos, suelen ser muy satisfactorios por los buenos rendimientos en la producción (Ramírez et al., 2005). Entre los productos probióticos con propósito aviar, se destacan aquellos que contienen microorganismos autóctonos del tracto gastrointestinal de aves en estado saludable (Miroslava et al., 2004) constituidas principalmente por diferentes especies de los géneros bacterianos: Lactobacillus, bacillus, estreptococos, enterococos y levaduras saccharomyces; los cuales han demostrado una alta eficiencia en la reducción de patógenos intestinales y en el aumento de los indicadores bioproductivos (Corrier et al., 1998).
La especificidad de especies probióticas en el animal, es un factor importante que interfiere en la colonización y en la adhesión in vivo de los microorganismos, (Frizzo et al., 2006); es necesario realizar una adecuada evaluación de cepas de acuerdo con diferentes criterios de selección, (resistencia al ácido, sales, temperatura, entre otras), de forma tal que los microorganismos colonizadores lleguen en estado viable y en cantidades suficientes una vez que han superado las barreras ácida y biliar en el tracto digestivo (Tuomola et al., 2001); los resultados de estas pruebas también pueden predecir la capacidad in vivo de las cepas; de ahí que, el éxito de un probiótico depende en gran medida de realizar una buena selección in vitro (Rondón et al., 2008).
El objetivo de la presente investigación fue evaluar cepas nativas aisladas de heces de aves silvestres (Gallus gallus), ante las principales barreras químicas del tránsito gastrointestinal, para seleccionar aquellas que presenten mayor potencial probiótico y que puedan ser utilizadas para elaborar aditivos microbianos regionales destinados a la dieta de polluelos.
Materiales y métodos
Aislamiento de los microorganismos.
A partir de un pool de heces de 30 aves (Gallus gallus) sanas, de 1 a 4 meses de edad, alimentadas a base de maíz, restos de cocina, vegetales y sin antibióticos, se aislaron bacterias del genero Bacillus sp., Lactobacillus sp. y levaduras tipo Saccharomyces sp. Fueron utilizados medios selectivos, Agar De Man, Rugosa, Sharpe (MRS) (Lactobacillus sp.), Nutritivo (Bacillus sp) y Rosa Bengala (RB) (Saccharomyces sp.) teniendo en cuenta las condiciones de crecimiento (Rondón et al., 2008).
Cada cepa aislada fue sometida a cuatro pruebas preliminares: tinción de Gram, tinción de endosporas (método de Wirtz), pruebas catalasa y oxidasa.
Lactobacillus sp.: se seleccionaron aquellas cepas catalasa negativa, oxidasa negativa y bacilos Gram positivos en cadenas no esporulados (Kandler y Weiss, 1986).
Bacillus sp.: se seleccionaron aquellas cepas catalasa positiva, oxidasa positiva y bacilos Gram positivos endosporados (Granados et al., 1997).
Saccharomyces sp.: se le realizó tinción de Gram y montaje con azul de lactofenol (Kreger, 1984; Madigan et al., 2004).
Evaluación de las propiedades Probioticas in vitro.
A una concentración conocida inoculo (10 6 UFC/mL) de cada uno de los microorganismos se le realizaron las siguientes pruebas:
a) Tolerancia a cambio de pH: se evaluaron las cepas a diferentes valores de pH, 3, 4, 5.6, 7 (incubación a 37 °C durante 24 hrs); la sobrevivencia y resistencia se comprobó al comparar el conteo de microorganismos viables del inoculo, con las células sobrevivientes después de la incubación a diferentes valores de pH (Zavaglia et al., 1998); el porcentaje de resistencia fue calculado por la siguiente ecuación (Kociubinski et al., 1999):
% R pH = [(UFC/mL) MRS pH x 100] / (UFC/mL) MRS (inoculo)
Se estableció como criterio de selección escoger aquellas cepas que resistieron el pH por encima del 50 % y se les realizó las siguientes pruebas probióticas.
b) Tolerancia a sales biliares: el ensayo fue realizado a diferentes concentraciones de sales 0,05, 0,1, 0,15 y 0,3 % p/v ajustado el pH =7 con HCI 5% (incubación a 37 °C durante 24 hrs); al cabo de este tiempo la sobrevivencia y resistencia a sales biliares se comprobó mediante la determinación del número de células viables (UFC) (Brizuela, 2003; Rubio et al., 2008, Rondón et al., 2008; Avila et al., 2010).
c) Tolerancia a cambios de temperatura: fueron probadas diferentes temperaturas, 28, 37 y 43 °C, durante 24 hrs; la sobrevivencia y resistencia a estas temperaturas se comprobó mediante la determinación del número de células viables (UFC) (Rubio et al., 2008, Rondón et al., 2008; Ávila et al., 2010).
d) Tolerancia a altas concentraciones de NaCl: la prueba se realizó utilizando caldos MRS, Malta, Nutritivo y diferentes concentraciones de NaCl, 2, 4, 7 y 10 % p/v, (incubación a 37 °C durante 24 hrs); finalizado el tiempo se determino el crecimiento a altas concentraciones de NaCl mediante la medición de la densidad óptica (DO) a 600 nm a 0 y 24 hrs. (Rondón et al., 2008).
e) Fermentación de la glucosa: se utilizaron los caldos MRS, Malta, Nutritivo, que contenían 0,2 % v/v de una solución de purpura de bromocresol (0.5 %) y campanas de Durham; la producción de gas se evidencio mediante la presencia de gas en las campanas y luego de incubación a 37 °C durante 48 hrs (Rubio et al., 2008, Rondón et al., 2008).
f) Prueba de antagonismo: esta prueba fue realizada contra Salmonella sp y E. coli pertenecientes al banco de cepas del laboratorio GRUBIODEQ, las cuales se sembraron en forma masiva en agar Mueller hinton; en la superficie de estos se colocaron tres discos impregnados con los microorganismos de ensayo; se introdujeron en la nevera durante 30 minutos (T= 15 0C) y después se incubaron a 37 °C durante 48 hrs. La acción antagónica se evidencio por la presencia de halos de inhibición y crecimiento alrededor de los discos. (Leiva et al., 2004; Mejía et al., 2007).
g) Capacidad de crecimiento: las cepas que resistieron la presencia de sales biliares, acidez se cultivaron en 30 ml de caldo MRS, Malta, Nutritivo y fueron incubadas a 37°C durante 24 y 48 hrs; el crecimiento se comprobó mediante la determinación del número de células viables (UFC) y se determinó el porcentaje (Laurencio et al., 2002; Rondón et al., 2008).
Identificación de los microorganismos con potencial Probiótico.
Los tres microorganismos que presentaron mejores propiedades probióticas (Lactobacillus sp, Bacillus sp., Saccharomyces sp.), se identificaron mediante pruebas bioquímicas: TSI, SIM, UREA, CITRATO, LIA, NITRATO; y los carbohidratos: glucosa, sacarosa, lactosa, maltosa, manosa, galactosa, xilosa, fructosa; Se incubaron a 37 °C durante 24 hrs (Bergey'S. 1994; Kandler y Weiss, 1986). Todos los ensayos por triplicado.
Análisis Estadístico.
Para el análisis de los datos se utilizó el programa Statistix for Windows®; el análisis de varianza se realizó para verificar diferencias significativas entre las medias, con un nivel de significancia con α = 0,05 y la prueba de Tukey fue usada para realizar las comparaciones múltiples entre las medias.
Resultados y discusión
Se aislaron un total de 20 cepas y de acuerdo a sus características macroscópicas y microscópicas, solo 11 correspondieron a los géneros de interés (Bacillus, Lactobacillus y Saccharomyces): 4 Lactobacillus sp, 5 Bacillus sp y 2 Saccharomyces sp.
a) Tolerancia a cambio de pH. De las 11 cepas evaluadas solo 3: Saccharomyces sp (3), Bacillus sp (7) y Lactobacillus sp (14) resistieron el pH= 3 durante 24 hrs, aunque se evidencio una disminución porcentual del crecimiento en los tres géneros. Sin embargo a pH= 4, se presentó el mayor porcentaje de crecimiento de las tres cepas lo cual indica la supervivencia en condiciones de acidez; mientras que a pH= 5.6, 7 disminuyó un poco pero también se mantuvo estable el crecimiento (tabla 1); por lo tanto solo estas tres cepas fueron seleccionadas para realizar las pruebas de resistencia a sales biliares.
Similares resultados se observaron con Lactobacillus salivarius (Rondón et al. (2008) y L. rhamnosus (Brizuela, (2003) en los cuales se evidenció poco crecimiento a pH de 3 y un incremento significativo de la población a pH de 5.
Es interesante que las cepas evaluadas en la presente investigación toleren pH= 3 durante 24 hrs, ya que existen hallazgos en donde la cepa Lactobacillus acidophilus M92 solo pudo sobrevivir durante 3 hrs a este pH, produciéndose luego la lisis del 60 % de la población inicial (Suskovic et al., 1997). Introducir como criterio de selección la supervivencia a pH de 3, permitió detectar los microorganismos resistentes a estas condiciones extremas, sobre todo si se parte del hecho que el jugo gástrico de los pollos puede llegar a valores de pH de 0,5 - 2,0 (Ehrmann et al., 2002). En el caso de la levadura, la tolerancia al ácido puede deberse a la presencia de proteínas especializadas que catalizan el intercambio de cationes monovalentes (Na+ o K+) e H+ a través de las membranas, regulando las concentraciones de cationes y el pH a nivel citoplasmático y de organelos (Mitsui et al., 2005; Ohgaki et al., 2005). Otro de los posibles mecanismos de regulación es a través de la ATPasa localizada en la membrana citoplasmática. Esta puede crear un gradiente electroquímico de protones que conduce al transporte secundario de solutos y que está implicado en el mantenimiento del pH cercano a la neutralidad (Viegas et al., 1998; Sychrovae et al., 1999). En el género Bacillus, la tolerancia a la acides gástrica ha sido asociada a la presencia de endosporas (Jiraphocakul et al., 1990), cuyas estructuras contienen el complejo calcio - acido dipicolinico -peptidoglicano que confieren la resistencia a situaciones adversas del medio ambiente como: pH, T°, radiaciones, desecación, déficit nutricional, altas presiones y agentes químicos (Madigan et al., 2004).
Los Lactobacillus son generalmente más resistentes a las condiciones acidas que otras bacterias lácticas, siendo capaces de crecer hasta un pH de 4, esta resistencia les permite seguir creciendo durante las fermentaciones lácticas naturales, aún cuando el pH haya caído tanto, que otras bacterias lácticas ya no puedan crecer (Madigan et al., 2004).
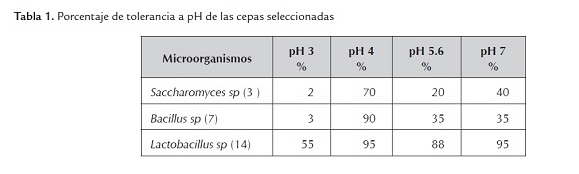
b) Tolerancia a sales biliares. Se observó que las 3 cepas son capaces de tolerar el rango de las concentraciones de sales biliares experimentadas (figura 1); estadísticamente en los tres microorganismos se presenta un efecto significativo en el crecimiento a medida que aumenta la concentración de sales biliares. De acuerdo a los resultados obtenidos se puede decir, que las tres cepas nativas son microorganismos capaces de sobrevivir a concentraciones de sales biliares desde 0,05% (p/v) hasta 0,3% (p/v), pudiendo desarrollar sus actividades metabólicas sin verse completamente inhibidas, típico en microorganismos aislados de aves de corral.
Se ha descubierto que las levadura tipo S. cerevisiae soportan concentraciones de sales biliares debido a la presencia de proteínas integrales de membrana unidas a ATP (proteínas ABC), responsables de la translocación de las sales biliares pudiendo transportar eficientemente ácidos biliares conjugados. Otro posible mecanismo es la acumulación de polioles y glicerol, para regular la presión osmótica de la célula. (Ortiz et al., 1997).
Las bacterias ácido lácticas del género Lactobacillus, son capaces de producir la enzima conocida como sal biliar hidrolasa (SBH), que cataliza la hidrólisis de las sales biliares conjugadas con glicina y taurina. Esta desconjugación pudiera ocurrir en la fase estacionaria del crecimiento bacteriano, ya que la actividad de la SBH se incrementa al disminuir el pH por producción de gran cantidad de ácidos orgánicos (Corzo y Gilliland, 1999). Las diferencias en la tolerancia al tránsito gastrointestinal también pueden deberse a las diferencias existentes en la estructura de la pared celular de las distintas especies y géneros bacterianos (Prasad et al., 1998).
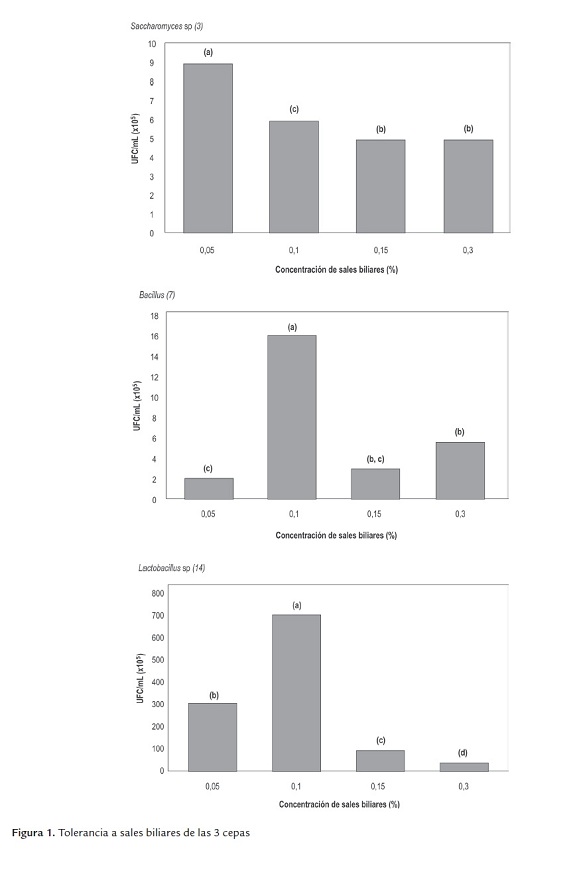
c) Tolerancia a cambio de temperatura. Las tres cepas toleraron las temperaturas de 28, 37 y 43°C durante 24 hrs; estadísticamente hay diferencia significativa, lo cual evidencia el efecto de la temperatura sobre el crecimiento de cada microorganismo; la temperatura de 37 °C fue la que mostró mejores resultados para Saccharomyces sp (3) y Bacillus sp (7), mientras que para Lactobacillus sp (14) fue de 43 °C (figura 2).
La temperatura es uno de los factores más importantes que afectan al crecimiento y supervivencia de los microorganismos, varía entre los diferentes géneros y reflejan el rango de temperatura optima de su habitad natural; a medida que se eleva la temperatura, las reacciones químicas y enzimáticas son más rápidas y el crecimiento se acelera, hasta el punto en que tienen lugar las reacciones de inactivación (Madigan et al., 2004).
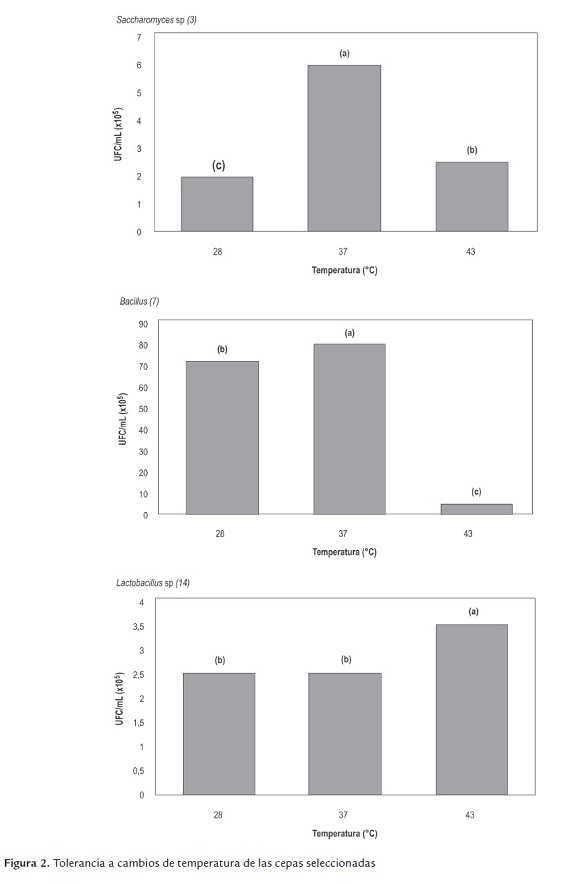
d) Tolerancia a altas concentraciones de NaCl. Se observó un efecto significativo en el crecimiento de los tres microorganismos a medida que aumentó la concentración de NaCl, siendo notable en 2 y 4% y muy bajo al 7 % y 10% p/v de NaCl (figura 3).
La mayoría de las bacterias son relativamente insensibles a las variaciones de la presión osmótica, y se adaptan a cambios fuertes en la concentración de solutos del medio, porque poseen una pared celular mecánicamente rígida. Basados en lo anterior, se deduce que las cepas nativas tienen cierta capacidad de acondicionarse ante determinadas concentraciones de sales, siendo 7% la máxima condición de salinidad tolerada hipertónicamente.
e) Produccion de gas a partir de la glucosa. Saccharomyces sp (3), Bacillus sp. (7) y Lactobacillus sp. (14) fermentaron la glucosa sin producción de gas, durante las 24 hrs de incubación, demostrando una asimilación del carbohidrato; también se evidencia la ruta metabólica homofermentativa utilizada por el género Lactobacillus sp., indicando la producción de acido láctico.
f) Prueba de antagonismo. La prueba de antagonismo frente a patógenos, demostró que Saccharomyces sp. (3) y Bacillus sp. (7) no producen sustancias antimicrobianas que puedan difundirse al medio y sean capaces de contrarrestar in vitro el crecimiento de los patógenos evaluados. Si se observó que estos dos géneros realizan exclusión competitiva haciéndolos resistentes al ambiente gastrointestinal siendo su colonización y multiplicación más fructífera en comparación con los patógenos.
Lactobacillus sp. (14) si produjo halos de inhibición de 1mm frente a los 2 patógenos, lo cual puede deberse a la producción de sustancias inhibitorias; se ha reportado que este género se caracteriza por la producción de bacteriocinas además del acido láctico (Domitille et al; 2005).
g) Capacidad de crecimiento. Los tres microorganismos de estudio mostraron altos valores de crecimiento luego de resistir las condiciones expuestas (tabla 2).
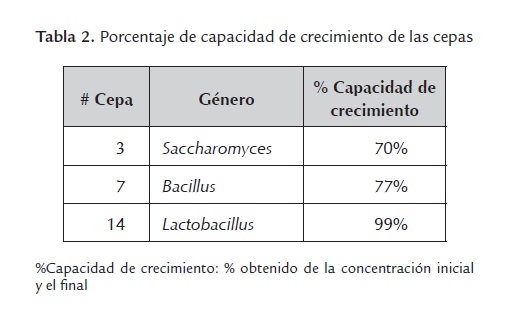
La capacidad de crecimiento es una propiedad que debe caracterizar a las cepas probióticas, ya que deben presentarse en cantidades suficientes para llegar al tracto gastrointestinal TGI, resistir los impedimentos químicos que aquí se presentan y ser capaces de establecerse para lograr una buena colonización de la mucosa y el contenido intestinal (Salminen et al; 1996).
En los productos con mayor actividad probiótica se emplean los microorganismos autóctonos del tracto digestivo de los animales; se destacan por su efectividad en la mejora de los indicadores de salud, al producir la exclusión de microorganismos potencialmente patógenos, así como por el estimulo de la respuesta inmune; además de intervenir en la prevención de enfermedades infecciosas en los animales y en el hombre.
Conclusiones
Se obtuvieron 3 cepas nativas con potencial probiótico pertenecientes a los géneros: Lactobacillus sp., Bacillus sp. y Saccharomyces sp., las cuales mostraron la mayor tolerancia a las condiciones del tracto gastrointestinal como: crecimiento a pH 3, concentración de sales biliares y de NaCl de 0.3 %, y 7 % respectivamente, T = 43°C, exclusión competitiva a patógenos y alta Capacidad de crecimiento.
Los resultados obtenidos in vitro demuestran que las tres cepas nativas poseen propiedades probióticas y pueden ser utilizadas como aditivos microbianos destinados a la alimentación de pollos recién eclosionados, para controlar su microbiota intestinal benéfica, estimular su sistema inmune, inhibir el crecimiento de patógenos oportunistas e incrementar los índices bioproductivos del sector avícola del departamento de Córdoba. Sin embargo se requieren estudios in vivo para validar sus efectos benéficos.
Agradecimientos
Los autores agradecemos a la Universidad de Córdoba y a la Gobernación de Córdoba por la financiación de parte del presente trabajo de investigación.
Referencias bibliográficas
1 Ávila, J., Ávila, M., Tovar, B., Brizuela, M., Perazzo, y., Hernández, H. 2010. capacidad probiótica de cepas del género Lactobacillus extraídas del tracto intestinal de animales de granja. Revista Científica Universidad de Zulia-Venezuela, 20(2):161-169. [ Links ]
2 Barnes, H. J., Vaillancourt, J. P., Gross, W. B. 2003. "Colibacillosis" Diseases of Poultry, 11th Edition, Section II, Chapter 18. [ Links ]
3 Bergey's. 1994. Manual of determinative Bacteriology.9 edition. Edited by john G.Holt Copyright Williams y Wilkins, Baltimore. New York; EE.UU; ISBN 0-683-00603-7. [ Links ]
4 Brizuela, M. 2003. Selección de cepas de bacterias ácido lácticas para la obtención de un preparado con propiedades probióticas y su evaluación en cerdos. Tesis Doctoral. ICIDCA, Cuba. p.101. [ Links ]
5 Corrier, D.E., Byrdii, J.A., Hume, M.E., Nisbet, D.J., Stanker, L.H. 1998. Effect of simultaneous or delayed competitive exclusion treatment on the spread of salmonella in chicks. J.Appl.poultry Res: pp.132-137. [ Links ]
6 Corzo, G., Gilliland, S. E. 1999. Bile salt hydrolase activity of three strains of Lactobacillus acidophilus. Journal of Dairy Science 82: 472-480. [ Links ]
7 Domitille, F., Ce'dric, N., Berger, M., Coconnier, V., Moal, L., Servin, A. 2005.pH-, Lactic Acid-, and Non-Lactic Acid- Dependent Activities of Probiotic Lactobacilli against Salmonella enteric serovar typhimurium. Appl Env Microbiol; 71(10): 6008-6013. [ Links ]
8 Ehrmann, M. A., Kurzak, P., Bauer, J., Vogel, R. F. 2002. Characterization of lactobacilli towards their use as probiotic adjuncts in poultry. Journal of Applied Microbiology, 92: 966-975. [ Links ]
9 Frizzo, L., Soto, L., Bertozzi, E., Sequiera, E., Marti, L., Rosmini, M. 2006. Evaluación in vitro de las capacidades probióticas orientadas al diseño de inóculos probióticos multiespecie para ser utilizados en la crianza de terneros. Rev FAVE. 5: 1-2. [ Links ]
10 Granados, R., Villaverde, M. 1997. Microbiologia.primera Edicion.Thomson paraninfo, S.A. Madrid, España. p. 8, 13, 323. ISBN 84-9732-123-5. [ Links ]
11 Jiraphocakul, S., Sullivan, W.T., Shahani, M, K.1990. Influence of dried B.subtilis culture and antibiotics as performance and intestinal microflora in turkey. Puoltry Science (9):1966-1973. [ Links ]
12 Kanlder, O., Weiss, N. 1986. Regular nonsporing Gram positive rods. Section 14 In: Garrity, G. (Ed) Bergey's Manual of Systematic Bacteriology. Springer, New York. 2:1208-1260. [ Links ]
13 Kociubinskik, G., Pérez, P., De Antoni, G. 1999. Screening of bile resistance and bile of precipitation in lactic acid bacteria and bifidobacteria. Journal of Food Protection 62, 905- 912. [ Links ]
14 Kreger, V., Rij, M.J.W. 1984. The yeasts. A taxonomy study. Elsevier, Ámsterdam. [ Links ]
15 La Ragione, R. M., and woodward, M. J. 2002. "Virulence factors of Escherichia coli serotypes associated with avian colisepticemia" Research in Veterinary Science, 73: 27-35. [ Links ]
16 Laurencio, M., Pérez, M., Piad, R., Milián, G., Rondón, A. J., Amigo, S., Bocourt, R., Savón, L. & Díaz, M. 2002. Determinación del contenido de bacterias ácido lácticas y Bacillus del tracto gastrointestinal de pollos de ceba y obtención de una mezcla de Exclusión Competitiva. Rev. Cubana Ciencia Avícola 26: 17-22. [ Links ]
17 Leiva, S., M. Yañez, L. Zaror, H. Rodríguez & H. García-Quintana. 2004. Actividad antimicrobiana de Actinomycetales aislados desde ambientes acuáticos de sur de Chile. Rev. Med. Chile; 132; 151-159. [ Links ]
18 Lu, J., Idris, U., Harman, B., Ofacre C., Murer, J.Y., Lee, M. 2003. Diversity and succession of the intestinal bacterial community of the maturiting Broilers chicken. Applied and environmental microbiology 69(11):6816-6824. [ Links ]
19 Madigan, M.T., Martinko, J.M., Parker, J. Brock. 2004. Biología de los Microorganismos. 10a ed. Pearson Education, Madrid. [ Links ]
20 Mejia, J., Chacon, Z., Guerrero, B., Otoniel, J., López, G. 2007. Obtencion de cepas de Lactobacillus: caracterización in vitro como potenciales probióticas. Revista científica.Universidad de Zulia Maracaibo, Venezuela. 17(2) 178-185. [ Links ]
21 Miroslava, M., Viola, S., Klaudia, B., Andrea, L., Sona, G. 2004. Effect of probiotic Activity of Enterococcus faecium EE3 strain against Salmonella infection in Japanese quails. Bull veterinary institute pulawy 48(3)387-390. [ Links ]
22 Mitsui, K., Yasui, H., Nakamura, N., Kanazawa, H. 2005. Oligomerization of the Saccharomyces cerevisiae Na+/H+ antiporter Nha1p: Implications for its antiporter activity. Biochim Biophys Acta; 1720: 125-136. [ Links ]
23 Ohgaki, R., Nakamura, N., Mitsui, K., Kanazawa, H. 2005. Characterization of the ion transport activity of the budding yeast Na+/H+ antiporter, Nha1p, using isolated secretory vesicles. Biochim Biophys Acta; 1712: 185-196. [ Links ]
24 Ortiz, D., Stpierre M., Abdulmessih, A., Arias I. 1997. A yeast ATP-binding cassette type protein medianting ATP-dependent bile acid transport. J Biol Chem; 272: 15358-15365. [ Links ]
25 Prasad, J., Gill, H., Smart, J., Gopal, P. K. 1998. Selection and characterization of Lactobacillus and Bifidobacterium strains for use as probiotic. International Dairy Joumal 8: 993-1002. [ Links ]
26 Ramírez, B. O., Zambrano, S, Y., Ramírez P, Y., Rodríguez V, Y., Morales, M. 2005. Evaluación del efecto probiótico del Lactobacillus sp. origen aviar en pollitas de inicio reemplazo de la ponedora comercial en los primeros 42 días de edad. Revista Electrónica de Veterinaria REDVET. p.8 [ Links ]
27 Rondón, A. J., Samaniego, L. M., Bocourt, R., Rodríguez, S., Milián, G., Ranilla, M. J., Laurencio, M., Pérez, M. 2008. Aislamiento, identificación y caracterización parcial de las propiedades probióticas de cepas de Lactobacillus sp. Procedentes del tracto gastrointestinal de pollos de ceba. Rev.Somenta (México) 6(1):56-63. [ Links ]
28 Rubio M, A., Hernández E, M., Aguirre R, A., Poutou P.R. 2008. Identificación preliminar in vitro de propiedades probióticas en cepas de S. cerevisiae .Revista MVZ Córdoba, Vol. 13 (1): 1157-1169. [ Links ]
29 Salminen, S., Laine, M., Von Wrigh, A., Vuopio-Varkila, L., Korhonen, T., Mattila-Sandholm, T. 1996. Development of selection criteria for probiotic strains to assess their potential in functional foods: a Nordic and European approach. Bioscience Microflora 15: 61-70. [ Links ]
30 Suskovic, J., Brkic, B., Matisic, S., Maric, V. 1997. Lactobacillus acidophilus M92 as potencial probiotic strain, Milchwissenschaft. 52:430-435. [ Links ]
31 Sychrovae, H., Ramírez, J., Peña, A. 1999. Involvement of Nha1 antiporter in regulation of intracellular pH in Saccharomyces cerevisiae. Microbiol Let; 171: 167-172. [ Links ]
32 Tannock, G. W. 1999. A fresh look at the intestinal microflora.Chapter 1. In: Probiotics. A Critical Review. Ed. by G. W. Tannock. Horizon Scientific Press. England. [ Links ]
33 Tuomola, E. M., Crittenden, R., Playne, M., Isolauri, E., Salminen, S. J. 2001. Quality assurance criteria for probiotic bacteria. American Journal of Clinical Nutrition 73:393-398. [ Links ]
34 Viegas, C., Almeida, P., Cavaco M, I. C. 1998. The H1-ATPase in the plasma membrane of Saccharomyces cerevisiae is activated during growth latency in Octanoic Acid-supplemented medium accompanying the decrease in intracellular pH and cell viability. Appl Env Microbiol; 64: 779-783. [ Links ]
35 Zavaglia, A. G., Kociubinski, G., Pérez, P., De Antoni, G. 1998. Isolation and characterization of Bifidobacterium strains for probiotic formulation. Journal Food Protection 61:865-873. [ Links ]

