










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.14 no.1 Bogotá Jan./June 2012
ARTÍCULO DE INVESTIGACIÓN
Búsqueda y selección de una proteasa fúngica con potencial aplicación en la restauración de documentos históricos en el Archivo de Bogotá
Screening for a fungal protease with potential in the biorestoration of historical valuable documents in Bogota Archive
Carlos Alberto Cruz Ramírez1, Luz Stella Villalba Corredor2, José Fernando Mikán Venegas3, Sonia Amparo Ospina Sánchez4.
1Biólogo, cM.Sc en Microbiología. Centro Internacional de Física (CIF) & Laboratorio de Tecnología en Enzimas, Instituto de Biotecnología, Universidad Nacional de Colombia.
2Bacterióloga, M.Sc. Laboratorio de Física, Química, y Biología, Archivo de Bogotá.
3Biólogo, M.Sc., Ph.D. Laboratorio de Investigaciones, Facultad de Medicina, Universidad Militar Nueva Granada.
4Química farmacéutica, M.Sc., Ph.D. Laboratorio de Tecnología en Enzimas, Instituto de Biotecnología, Universidad Nacional de Colombia-Sede Bogotá.
Recibido: julio 1 de 2011 Aprobado: junio 22 de 2012
Resumen
Se ha buscado y seleccionado sistemáticamente una proteasa que pudiese ser usada en la eliminación "limpia" de encolantes sobre soportes documentales con valor de patrimonio histórico de forma eficiente y económica, a partir de la colección de hongos filamentosos del Archivo de Bogotá. De 74 morfotipos viables evaluados sobre placas selectivas, 32 morfotipos presentaron formación de halos de hidrólisis evidentes sobre placas diferenciales. De ellos, se evaluó el perfil isoenzimático de 8 morfotipos provenientes de muestreos documentales directos y de 2 morfotipos proteolíticos promisorios provenientes de un trabajo previo. Los 10 morfotipos seleccionados fueron representativos de los géneros Penicillium, Stachybotrys, Chaetomium, y Eladia. Luego de inducir la producción de proteasas extracelulares en medios líquidos diferenciales bajo tres fases de fermentación, se realizaron isoelectroénfoques analíticos tendientes a la observación de isoformas en el gradiente de pH establecido (3.0-10.0). Solo los morfotipos 8D (Chaetomium sp.) y 21D (Eladia saccula) presentaron una isoforma alcalina extrema, de puntos isoeléctricos 8.5 y 8.8, respectivamente, susceptible de selección con miras a su purificación y caracterización parcial de forma económica y eficiente. Los demás morfotipos, representativos de los géneros Penicillium sp., y Stachybotrys sp., presentaron unicamente isoformas proteolíticas en el rango acido de pH con puntos isoeléctricos que oscilan entre 4.0 y 5.0.
Palabras clave: biodeterioro, hongo filamentoso, proteasa, halo de hidrólisis, punto isoeléctrico.
Abstract
Studies on a protease as an efficient, environmental friendly and relatively economical remover of residual proteins for historical valuable documents were performed and was selected for this work, from the filamentous fungi collection of the Bogota Archive. 32 morphotypes of 74 evaluated show hydrolytic activities over differential solid media. From them, 8 morphotypes obtained directly from documental samples and representative of the genera Penicillium and Stachybotrys were selected and their isoenzyme profile were tested. Also 2 previous morphotypes with promisorius proteolytic activities and representative of the genera Chaetomium and Eladia were analysed. Extracelullar proteases production was induced in differential liquid media on three fermentation steps and analitycal isoelectrofocusing were performed over pH 3.0-10.0 ranges. Only the morphotypes 8D (Chaetomium sp.), and 21D (Eladia sp.), showed an alkaline isoform with pIs 8.5 and 8.8, respectly, suceptible of selection for its purification and characterization through efficient and economical way. The others morphotypes only showed acid isoforms with pIs in the range of 4.0 and 5.0.
Key words: biodeterioration, filamentous fungi, protease, hydrolytic halo, isoelectric point.
Introducción
La historia de la humanidad ha estado ligada a la transmisión de información mediante el empleo de diversos tipos de materiales de origen natural, sintético y semisíntetico, los cuales han estado expuestos a deterioro tanto a nivel biológico como ambiental a lo largo del tiempo (Hueck 2001). Algunos fondos documentales con carácter histórico custodiados por el Archivo de Bogotá, entidad encargada de la protección de los recursos documentales de la ciudad mediante su acopio, conservación y organización, han presentado biodeterioro como consecuencia de la colonización puntual de hongos filamentosos que utilizan los componentes de las obras (principalmente proteínas y polisacáridos) como fuentes de carbono y nitrógeno mediante el empleo de sus complejos sistemas enzimáticos ocasionando deterioro sobre los soportes a nivel físico y químico dañando la estética y la técnica de registro (Villalba y Mikán 2005; Villalba et al., 2004; Mateus et al., 2001).
El conocimiento que se tiene de este proceso sobre soportes documentales es escaso y conducente a la aplicación de procedimientos tradicionales, de base química, contaminantes, tóxicos e inespecíficos empleados durante las prácticas de restauración. En consecuencia, el Archivo de Bogotá ha venido conformando, desde el año 2005, el cepario institucional resultado del aislamiento riguroso de morfotipos implicados en el proceso de biodeterioro, con el fin de conocer y aprovechar la diversidad biológica existente en nuestros archivos, una importante fuente de información para el entendimiento del proceso, y la base para dar soluciones integrales y prácticas entorno a la problemática del microdeterioro documental.
Bajo este contexto, la entidad estatal ha venido focalizando sus esfuerzos en la búsqueda de tratamientos alternativos para la remoción "limpia" de compuestos orgánicos residuales sobre los documentos, tales como proteínas encolantes y almidones, que no solo se convierten en blanco biológico detrimental, sino que también se modifican ampliamente con el tiempo volviéndose resistentes al uso común de solventes orgánicos, carbonatos y fosfatos (contaminantes y nocivos) empleados durante las prácticas de restauración. En ese sentido, se han venido aunando esfuerzos en la búsqueda y selección preliminar de perfiles proteolíticos idóneos para la eliminación de proteínas residuales, predominantemente en ambientes alcalinos (a manera de "detergente" en términos de limpieza y de "inhibición" de crecimiento microbiano ulterior, sin generación de condiciones de pH extremas que perjudiquen la estabilidad de los documentos) a partir de su cepario institucional (Rojas et al., 2009; Cruz 2008; Rojas 2007).
Este trabajo busca actualizar la información disponible sobre la selección de hongos filamentosos con perfiles proteolíticos promisorios a partir del cepario del Archivo de Bogotá, con miras a la elección acertada de una isoforma proteolítica de interés con base en su punto isoeléctrico para ser purificada en el menor número de pasos metodológicos posibles, caracterizada y empleada en la remoción económica, especifica, mínimamente intrusiva, y con baja toxicidad ambiental y humana, de proteínas residuales sobre fondos documentales seleccionados. Adicionalmente, se busca brindar localmente una alternativa al empleo de proteasas comerciales importadas como la tripsina y la proteinasa K que usualmente vuelven económicamente inviables los procesos de biorestauración debido a sus altos costos.
Materiales y métodos
Recuperación de hongos filamentosos
Los aislamientos de hongos filamentosos mantenidos en el cepario del Archivo de Bogotá, provenientes de muestreos ambientales y documentales realizados mediante asistencia técnica a los diferentes Archivos de Gestión Distrital, representativos de procesos de biodeterioro, y conservados en estado puro en agua estéril a 4°C mediante la técnica de cultivos monospóricos (Ho y Ko 1997), fueron recuperados en caldos Sabouraud suplementados con jugo V8 por un término de 24 horas a 28°C con una agitación constante de 150rpm. Los hongos fueron transferidos a placas Sabouraud pH 5.5 incubadas por 5-7 días a 28°C para la obtención de biomasa y la consecución de inóculos microbianos (Rojas et al., 2009).
Selección de hongos proteolíticos promisorios sobre placas selectivas
Se evaluó la actividad proteolítica de cada hongo filamentoso viable mediante la inoculación de placas de 3%(p/v) agar Caseinato de Calcio pH 7.2 con una suspensión de 106propágulos/mL (obtenidos a partir de cultivos en agar Sabouraud de 8 días de incubación) en pozo central teniendo en cuenta la metodología propuesta por Cowan y Daniel (1982). Los cultivos fueron realizados por triplicado e incubados por 72 horas a 28°C con el fin de observar directamente halos de hidrólisis que reflejaran las actividades hidrolíticas en cada placa. Los halos de degradación y el diámetro de las cepas fueron medidos con un calibrador con el fin de calcular el tamaño de los halos de hidrólisis manejando la metodología descrita por Pedroza et al. (2001). Se definió el factor de hidrólisis como la proporción existente entre el diámetro promedio del halo y el diámetro promedio de la cepa.
Producción de proteasas extracelulares en medios de cultivo líquidos
Se determino la producción de proteasas extracelulares en morfotipos seleccionados mediante la inoculación de medios líquidos bajo tres fases de fermentación: obtención de biomasa, inducción a estrés nutricional e inducción de proteasas (Cooper y Wood 1980 citado por Rojas et al., 2009). La producción de biomasa se realizó mediante la inoculación de 30mL de caldo Sabouraud con 100µL de una suspensión de 105-106 propágulos/mL incubando a 28ºC por 5 días con una agitación constante de 150 rpm. El micelio resultante fue recuperado asépticamente mediante filtración en papel y transferido a un medio mínimo de sales pH 7.0 (30 mL) [0.1% (p/v) KH2PO4, 0.2% (p/v) NaNO3, 0.05% (p/v) MgSO4, 1%(v/v) solución elementos traza: 0.1%(p/v) K2HPO4, 0.002%(p/v) NaMoO4, 0.002%(p/v) MnCl2, 0.002%(p/v) CuSO4, 0.001%(p/v) FeSO4, 0.0003%(p/v) ZnSO4, 0.052%(p/v) acido bórico, 0.052%(p/v) acido cítrico; ajustar pH a 7.0 con 1M NaOH], incubando a 28°C por 24 horas y 150 rpm. Finalmente, la biomasa fue recuperada y transferida a 30mL de Medio Mínimo de Sales suplementado con Caseína al 0.1%(p/v) incubando a 28°C por 3-4 días y 150 rpm. La biomasa resultante fue separada del extracto crudo mediante filtración en papel y conservada en congelación a -70ºC. El extracto crudo fue clarificado mediante centrifugación a 4500rpm por 10 minutos a 4°C, y conservado en congelación a -20°C para análisis posterior.
Determinación de la concentración total de proteínas
Se determino el contenido de proteínas totales en los extractos crudos empleando el método de Bradford modificado (Stoscheck 1990). En todos los casos se mezclaron: 10µL de la solución de proteína, 20µL de 1M NaOH y 200µL del colorante en placas de 96 pozos fondo plano. La mezcla fue equilibrada a temperatura ambiente por 5 minutos, y se determinó su absorbancia a 630 nm en un lector ELISA pioway biomedical microplate reader M3000® contra los blancos apropiados (generalmente agua desionizada o tampón de baja fuerza iónica). Se construyó una curva de calibración utilizando albúmina de suero bovino y lisozima como estándares manejando un rango de concentración de proteínas entre 0.06 - 2.5µg/µL.
Determinación de la actividad proteolítica
Se cuantifico la actividad proteolítica en los extractos crudos mediante la metodología descrita por Charney y Tomarelli (1947), empleando Azocaseína como sustrato de reacción. Se preparó una solución de 1.5%(p/v) azocaseína en 0.1M bicarbonato de sodio pH 8.3 para efectuar las digestiones enzimáticas. La reacción se desarrolló pipeteando 20µL de la muestra de proteínas en 200µL de la solución sustrato, permitiendo la incubación de la mezcla por 60 minutos a 45ºC en baño María. La reacción enzimática se detuvo mediante la adición de 200µL de una solución de 5%(p/v) acido tricloroacetico (TCA). Los tubos fueron equilibrados a 4ºC por 30 minutos, y centrifugados a 8000rpm por 20 minutos a la misma temperatura. Finalmente, se dispusieron 200µL de cada sobrenadante en una placa de 96 pozos fondo plano, y se efectuaron las lecturas de absorbancia en un lector ELISA pioway biomedical microplate reader M3000® a 405nm. Para efectos de este trabajo se definió una unidad equivalente de proteinasa K (Ueq) como la cantidad de enzima que libera 1µmol de tirosina positiva para Folin & Ciocalteus por minuto de reacción, bajo las condiciones experimentales del ensayo, siguiendo la metodología propuesta por Cruz (2011).
Revisión de perfiles isoenzimáticos para proteasas alcalinas
Se analizó el perfil isoenzimático de proteasas mediante isoelectroénfoque (IEF) analítico en geles de poliacrilamida para los extractos crudos que presentaron actividades hidrolíticas evidentes sobre azocaseína, siguiendo la metodología propuesta por Cruz (2008) y Mikán (2001). Los geles de poliacrilamida (125x105x0.4mm) fueron preparados empleando las siguientes soluciones (concentración final (v/v)): 15% acrilamida-bisacrilamida PlusOne ReadySol IEF T40 C3 (Pharmacia-Biotech®), 10% glicerol, 4% anfolitos (Pharmalyte pH 3 a 10, Pharmacia-Biotech®), 0,075% TEMED al 10%(p/v), 0,35% APS al 10%(p/v). Las corridas se realizaron en una cámara de electroforesis LKB Bromma 2117 Multiphor (Pharmacia®), a una temperatura regulada de 10ºC con un circulador termostático MultiTem III (Pharmacia®), empleando 1M NaOH (tira correspondiente al cátodo) y 1M H3PO4 (tira correspondiente al ánodo) como electrolitos. Las condiciones de corrido fueron: Precorrido 1000V, 8mA, 8Va por 30 minutos; Siembra y Corrido 2200V, 8mA, 8Va por 90 minutos; Focalización 3000V, 8mA, 8Va por 30 minutos.
Determinación del punto isoeléctrico (pI) de las proteínas
La sección del gel que contiene 10µL del marcador de proteínas (Broad range pI markers, Pharmacia®) fue cortada y fijada por 60 minutos en una solución 10%(p/v) TCA, 5%(p/v) 5-ácido sulfosalicílico, y 40%(v/v) metanol. Se realizó un lavado por 20 minutos en una solución etanol al 40%(v/v), y ácido acético al 10%(v/v), y se equilibró el gel por 10 minutos en la solución decolorante (30%(v/v) etanol, y 8%(v/v) ácido acético). Los marcadores fueron teñidos por 15 minutos en 0.5% (p/v) azul Coomassie R-250 disuelto en la solución decolorante y precalentado a 60°C. Finalmente, el gel se sometió a lavados sucesivos en la solución de decoloración hasta obtener un fondo claro. Para la estimación del isoeléctrico se determinó la distancia de cada marcador pI al cátodo y se graficó contra su correspondiente pI teórico. Una vez obtenida la gráfica, se realizó una conversión de la distancia al cátodo de las proteínas de interés, reveladas mediante zimogramas, a punto isoeléctrico (pI) mediante el empleo de la ecuación de la curva.
Zimogramas
La actividad proteolítica en gel se detectó mediante el método de Peyronel y Cantera (1995), incubando el gel de proteínas post-isoelectroénfoque en una solución de 1.5%(p/v) azocaseína disuelta en 0.1M bicarbonato de sodio pH 8.3 a 45ºC por 60 minutos en baño María. Posterior a la incubación se realizó un lavado con agua desionizada por 1 minuto, y se fijo el gel en una solución de 12.5%(p/v) TCA a 45ºC por 60 minutos. Se efectuó un lavado con agua desionizada por 1 minuto para revelar las zonas claras de hidrólisis en un fondo naranja oscuro sumergiendo el gel en una solución 1N NaOH.
Se realizaron algunas modificaciones al método anterior sobreponiendo el gel de proteínas post-isoelectroénfoque con un gel sustrato compuesto de gelatina comercial o albumina de suero bovino disuelta en 0.1M bicarbonato de sodio pH 8.3 y 1.5%(p/v) agarosa. El gel sustrato fue superpuesto evitando la formación de burbujas y asegurando un contacto estrecho con el gel de interés. Luego de incubar a 45ºC por 30 minutos, el gel sustrato fue retirado y fijado en una solución 40%(v/v) etanol, 10%(v/v) ácido acético, disueltos en agua destilada por 60 minutos. El gel fue teñido empleando la coloración azul Coomassie enunciada anteriormente, hasta que se visualizaran zonas claras de hidrólisis en el gel.
Resultados
Selección de hongos proteolíticos promisorios sobre medios de cultivo sólidos
Se evaluaron las actividades proteolíticas de 74 morfotipos viables mantenidos en el cepario del Archivo de Bogotá, provenientes de muestreos ambientales y documentales realizados mediante asistencia técnica a los diferentes Archivos de Gestión Distrital. De ellos, el 43.2% (32 morfotipos) presentaron actividades proteolíticas evidentes mediante la formación de halos de hidrólisis sobre placas de agar Caseinato de Calcio (figura 1-1). Se seleccionaron los morfotipos 22A, 23D y 55D (Penicillium spp.), 55A y 60A (NN), y 34A (Stachybotrys sp.) cuyo factor de hidrólisis igualaba o superaba 0.90 (umbral) denotando mayor producción enzimática en relación al crecimiento de la cepa. Esta relación facilitaba los procesos de filtrado y reducía la secreción de proteínas "contaminantes" propias del desarrollo microbiano que pudiesen interferir en procesos de purificación ulteriores.
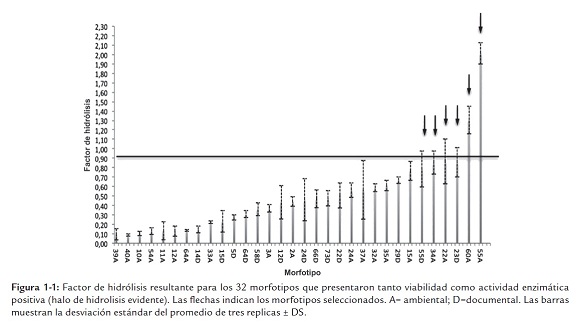
A pesar de la prevalencia de hongos proteolíticos promisorios provenientes de muestreos ambientales, se decidió seguir con los morfotipos obtenidos directamente de documentos con indicadores de biodeterioro debido a su estrecha relación con los componentes de los soportes objeto de estudio (principalmente proteínas y carbohidratos).
Producción de proteasas extracelulares en caldos de cultivo y revisión de perfiles isoenzimáticos para proteasas alcalinas en hongos filamentosos promisorios.
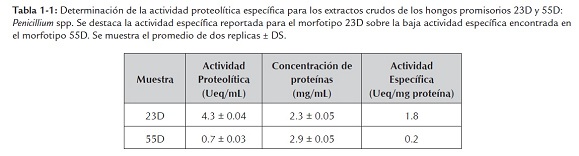
Los morfotipos 23D y 55D, seleccionados en el screening en placa, fueron cultivados en medios líquidos en tres fases de fermentación: obtención de biomasa, inducción a estrés nutricional e inducción de proteasas. Los extractos crudos resultantes fueron evaluados en cuanto al contenido de proteínas totales por el método de Bradford modificado y a su actividad proteolítica sobre azocaseína soluble. En la tabla 1-1 se reportan las actividades específicas para los extractos crudos, destacándose la actividad proteolítica del morfotipo 23D sobre la del morfotipo 55D.
En consecuencia, se determinó del perfil isoenzimático para proteasas en el extracto crudo del morfotipo 23D con el objeto de observar el número y distribución de isoformas a lo largo del gradiente de pH establecido. Del isoelectroénfoque se observo una única isoforma proteolítica evidente hacia el rango acido de pH (pI= 4.81) (figura 1-2) que hacia factible su purificación en el menor número de pasos metodológicos posibles. Sin embargo, se decidió realizar la búsqueda de proteasas de pI alcalino de fácil purificación en los seis hongos filamentosos "documentales" subsecuentes con mayores factores de hidrolisis durante el screening en placa descrito en la figura 1-1: Morfotipos 12D (Stachibotrys sp.) y 22D, 29D, 64D, 66D y 73D (Penicillium spp.). Estos hongos podían presentar proteasas de mayor estabilidad en procesos de biolimpieza sobre superficies alcalinas teniendo en cuenta que varios de los procesos de restauración documental que actualmente se emplean en el Archivo de Bogotá se llevan a cabo bajo dichas condiciones de pH. Igualmente, se decidió corroborar la secreción de dichas proteasas en dos hongos filamentosos seleccionados en un trabajo previo desarrollado en el Archivo de Bogotá: Morfotipos 8D (Chaetomium sp.) y 21D (Eladia saccula) (Rojas et al., 2009; Cruz 2008; Rojas 2007).

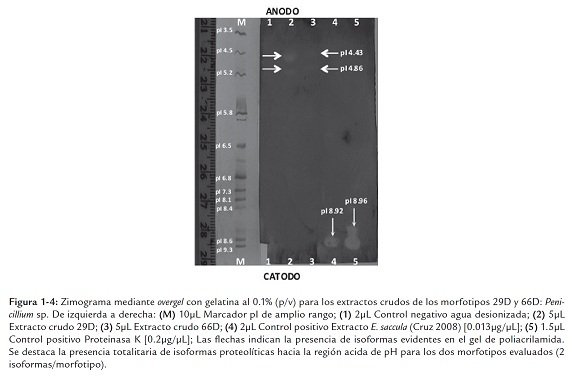
La figura 1-3 muestra las isoformas resultantes para seis de los ocho morfotipos evaluados: 12D, 22D, 73D, 64D, 8D, y 21D, manteniéndose la presencia de isoformas proteolíticas en el rango acido de pH, a excepción del morfotipo 8D que mostró una isoforma tenue hacia el extremo alcalino de pH. Se destaca la ausencia de isoformas para el morfotipo 21D en contraste con lo reportado por Rojas et al. (2009). Paralelamente, se desarrollaron nuevos ensayos de zimografía para los extractos crudos de los dos morfotipos restantes (29D y 66D: Penicillium sp.) mediante la superposición de overgeles sobre los geles de IEF luego del corrido electroforético (Cruz 2008). Este cambio metodológico busco mayor visibilidad de las isoformas que se observaban tenues en los ensayos de zimografía anteriores. En la figura 1-4 se presentan las isoformas proteolíticas resultantes para los morfotipos 29D y 66D en donde se observo claramente la "tendencia" microbiana hacia la secreción de isoformas acidas. Se destacan la resolución y claridad de las isoformas obtenidas en el registro fotográfico.

Como resultado, se iniciaron nuevas fermentaciones tendientes a reactivar las rutas metabólicas involucradas en la síntesis de proteasas extracelulares (principalmente de pI alcalino) que por razones como represión catabólica o estrés fisiológico por el método de conservación al que estaban sometidos los hongos en el cepario, se habían dejado de sintetizar. Los hongos seleccionados fueron los morfotipos 8D Chaetomium sp., 12D Stachybotrys sp., y 21D E. Saccula , en razón de: (i) el morfotipo 8D mostró una isoforma proteolítica alcalina concordante con los reportes previos susceptible de "amplificación" en términos de producción de proteínas; (ii) el morfotipo 12D servía como control del ensayo debido a su perfil productor de una única isoforma proteolítica "acida"; (iii) el morfotipo 21D no solo había presentado un alto potencial para la producción de una proteasa extracelular pI alcalino susceptible de purificación en el menor número de pasos metodológicos posibles en un trabajo previo (Rojas et al., 2009), sino que además era novedoso en el campo del biodeterioro documental.
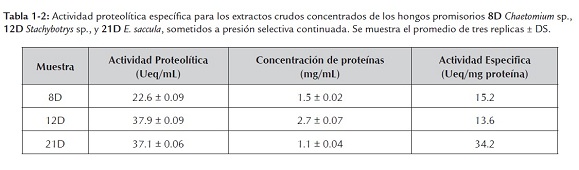
Se realizaron cultivos líquidos bajo presión diferencial continua (3 sub-cultivos/morfotipo) empleando extracto de carne al 0.1% (p/v) como única fuente de carbono y nitrógeno, bajo condiciones alcalinas (pH 8.0). Cada sub-cultivo fue desarrollado a 28°C por 8 días con una agitación constante de 150rpm. Luego del tercer sub-cultivo se inició el esquema de producción de proteasas convencional, y se estimó la actividad proteolítica especifica en los extractos crudos resultantes (tabla 1-2). Se destaca la alta actividad proteolítica obtenida en los tres morfotipos objeto de estudio y la recuperación de la actividad enzimática en el morfotipo 21D.
Los análisis de isoelectroénfoque permitieron dilucidar la recuperación y/o "amplificación" de la expresión de proteasas hacia la región alcalina de pH en dos de los géneros evaluados: Chaetomiun sp., y Eladia sp. Las figuras 1-5 y 1-6 revelan el perfil isoenzimático obtenido para las muestras en mención. En la Figura 1-5 se observan las isoformas resultantes para el morfotipo 21D por triplicado, resaltándose la presencia de una solo isoforma proteolítica de pI=8.8. En contraste, en la figura 1-6 se observa el perfil isoenzimático resultante para los morfotipos 8D y 12D, con 4 y 3 replicas, respectivamente. Se destaca la presencia de 4 isoformas evidentes para el género Chaetomiun sp. (pIs: 3.4, 4.5, 5.8 y 8.6), y dos isoformas acidas evidentes para el género Stachybotrys sp. (pIs: 3.7 y 5.6).


Con miras a la purificación y caracterización de una proteasa estable y activa bajo condiciones alcalinas con potencial para efectuar procesos de biolimpieza documental, se selecciono el perfil isoenzimático alcalino del morfotipo 21D (E. Saccula ) sobre el perfil del morfotipo 8D (Chaetomiun sp.) debido a que: (i) La presencia de una única isoforma "alcalina" extrema facilitaba el proceso de purificación mediante cromatografía de intercambio iónico, reduciendo las probabilidades de contaminación cruzada con otras proteínas residuales reduciendo la resolución y calidad del método; (ii) El desarrollo de biomasa durante los cultivos líquidos era cualitativamente menor en contraste con su homologo 8D, lo que facilitaba los procesos de recuperación del extracto crudo mediante filtración; (iii) El morfotipo 21D ha sido escasamente reportado en el campo del biodeterioro documental, en contraste con su homologo 8D que ha sido ampliamente reportado como agente biodeteriorante, lo que le otorgaba a 21D el carácter de rareza y especialidad.
Discusión
Usualmente durante los procesos de restauración de obras de arte y documentos históricos se hace necesaria la eliminacion selectiva y especifica de materiales de naturaleza proteica empleados como encolantes o adhesivos; es el caso de la cola de origen animal, clara y yema de huevo, y la caseína, entre otros. Los métodos tradicionales empleados para la remoción de estos compuestos se basan en la utilización de reactivos ácidos o alcalinos poco específicos y generadores de condiciones extremas de pH, así como el empleo de diversos solventes orgánicos nocivos para el operario y el ambiente. En ese sentido la tendencia actual a la hora de desarrollar un protocolo de limpieza se basa en la identificación fisicoquímica de los componentes de la obra ha ser removidos, y en la elección de un método de limpieza controlable y específico para cada caso en particular. En ese contexto, la utilización de proteasas para la fragmentación y remoción de sustancias de origen proteico se convierte en una alternativa viable a los procesos tradicionales de limpieza sin afectar la estabilidad de la obra (Castiñeyra et al., 2004; Decoux 2002; Segal y Cooper 1977).
En este trabajo se planteó un diseño experimental encaminado a la identificación y selección de un hongo filamentoso que produjera una proteasa con potencial para la remoción "limpia" de compuestos proteicos residuales, a manera de detergente, de forma eficiente, especifica y económica. Varios de los géneros microbianos empleados en esta investigación han sido previamente reportados en estudios relacionados con el biodeterioro de diversos tipos de materiales y con la producción industrial de enzimas (Rojas et al., 2009; Gorbushina et al., 2004; Florian y Manning 2000; Jellison y Jasalavich 2000; Szczepanowska y Cavaliere 2000; Baraznenok et al., 1999; Berner et al., 1997; Das et al., 1997; Zyska 1997).
La determinación de la actividad proteolítica sobre placas de agar Caseinato de Calcio libre de lactosa, atendiendo las recomendaciones enunciadas por Cruz (2008), Kudryavtseva et al. (2008), Wang et al. (2005), y Vermelho et al. (1996) referentes a la no inclusión de sacáridos en los medios de cultivo para evitar la represión por catabólito y la generación de ácidos que pudiesen afectar la viabilidad de las cepas, permitió seleccionar aislados con potencial proteolítico y visualizar la distribución del perfil catalítico en la población de estudio. De 74 morfotipos viables evaluados el 43.2% de los aislados presentaron actividades proteolíticas evidentes bajo las condiciones experimentales establecidas. De ellos, inicialmente se seleccionaron los morfotipos 23D y 55D (Penicillium spp.) debido a que el tamaño del halo de hidrólisis excedía claramente el tamaño de la cepa, lo que optimizaba un futuro proceso de purificación y caracterización enzimática teniendo en cuenta que, a menor biomasa menor fuente de proteínas y otros metabolitos contaminantes, y a mayor halo de hidrólisis mayor producción proteolítica extracelular.
Varios de los morfotipos viables no reportados presentaron un crecimiento miceliar estéril sin generación de halos de hidrólisis evidente, o no presentaron desarrollo biológico sobre las placas diferenciales, lo que sugiere especialización en otro tipo de sustratos orgánicos (por ejemplo almidón, celulosa, lípidos) e inorgánicos (por ejemplo citrato, fosfato, nitrato), estrés biótico por una conservación prolongada en agua refrigerada en el cepario del Archivo de Bogotá generando baja adaptabilidad inmediata a las condiciones diferenciales del medio, crecimiento a expensas de reservas endógenas de energía, o producción de proteasas fuertemente asociadas a la pared celular microbiana (Dienes et al., 2007; Iwashita 2002;, Reichard et al., 2000; Monod et al., 2002; Busconi et al., 1984).
Para los morfotipos seleccionados se ejecutaron cultivos líquidos tendientes a la producción de proteasas extracelulares bajo tres fases de fermentación: generación de biomasa sobre un medio rico en peptona y bajo en glucosa como fuentes de carbono y nitrógeno (caldo Sabouraud modificado), seguido de un sometimiento a estrés nutricional, culminando con una fase inductiva sobre extracto de carne como única fuente de carbono y nitrógeno. Esta estrategia esta soportada por estudios relacionados como el de Beg et al. (2002), citado por Gupta et al. (2002), los cuales concluyen que la producción de proteasas para Bacillus mojavensis se vio incrementada en cerca de cuatro veces cuando se empleaban métodos de fermentación de semi-batch o fed-batch a través de la separación de la fase de inducción de biomasa y producción de proteasas mediante el empleo de una fase de represión nutricional durante el crecimiento del microorganismo. Así mismo, Naidu y Lakshmi Devi (2005), citados por Savitha et al. (2011), reportaron un incremento significativo en la producción de proteasas alcalinas extracelulares a partir de la bacteria Bacillus sp. K30, mediante la inclusión de extracto de carne en los medios de cultivo inductivos como fuente de carbono y nitrógeno, sobre fuentes orgánicas como el almidón, salvado de arroz, caseína, extracto de levadura, y triptona.
El extracto crudo del morfotipo 23D presentó una actividad proteolítica evidente (4.3Ueq/mL) en contraste con el extracto crudo del morfotipo 55D que presentó una actividad enzimática depreciable (0.7Ueq/mL) a pesar de haber sido uno de los principales productores de proteasas mediante la estimación de halos de hidrólisis durante el screening en placa. Wang et al. (2005), Pandey (2003), e Iwashita (2002), mencionan que dada la naturaleza de los hongos filamentosos por desarrollarse sobre la superficie de plantas y animales, o inmersos en matrices complejas como el suelo, se puede esperar que la producción de proteínas extracelulares se vea favorecida por el empleo de sistemas de fermentación sólidos en contraste con los sistemas líquidos. De hecho, la diferencia en las condiciones de cultivo puede alterar la expresión de genes y afectar fenotipos como el crecimiento y desarrollo, la producción de micotoxinas, y la generación de enzimas. Varias de las enzimas producidas en fermentaciones líquidas generalmente se encuentran asociadas a la pared celular del micelio, mientras que en condiciones solidas generalmente se encuentran secretadas en el medio de cultivo.
Aunque el perfil isoenzimático para el extracto crudo del morfotipo 23D presentó una única isoforma proteolítica de pI 4.81 que hacia factible su purificación en el menor número de pasos metodológicos posibles, se decidió realizar la búsqueda de proteasas de pI alcalino de fácil purificación en los seis hongos filamentosos "documentales" subsecuentes con mayores factores de hidrolisis durante el screening en placa, así como en dos morfotipos proteolíticos promisorios seleccionados en un trabajo previo (Rojas et al., 2009). Estos hongos podían presentar proteasas de mayor estabilidad en procesos de biolimpieza sobre superficies alcalinas teniendo en cuenta que varios de los procesos de restauración documental que actualmente se emplean en el Archivo de Bogotá se llevan a cabo bajo dichas condiciones de pH.
De hecho, una de las principales características para la selección de una proteasa a incluir en los procesos de limpieza es su pI. Es bien conocido que las proteasas tienen un mejor desempeño cuando el valor de pH ambiental se aproxima al pI de la enzima, sin ser esto una generalidad (Shankar et al., 2011; Gupta et al., 2002). El pH afecta la ionización de los aminoácidos, lo que dictamina la conformación de la estructura primaria y secundaria de la enzima, controlando por ende su estabilidad y actividad enzimática. En ese sentido, entre más lejano se encuentre el pH ambiental del pI de la proteína, mayor será la tendencia a la desnaturación por: (i) reducción de la cisteína en condiciones alcalinas extremas; (ii) hidrólisis de los enlaces peptídicos proximales, generalmente, al acido aspártico en condiciones acidas extremas; (iii) repulsión electrostática intramolecular en zonas de la proteína con alta densidad de carga (positiva o negativa); (iv) competencia por sitios de unión a cofactores catiónicos debido al incremento en la concentración de hidroxilos, hidróxidos o hidrogeniones, según corresponda (Savitha et al., 2011; Vesterberg 1990; Garfin 1990).
Luego de obtener los extractos crudos y evaluar los perfiles isoenzimáticos mediante isoelectroénfoque analítico para los ocho morfotipos seleccionados, se pudo determinar la presencia de isoformas con puntos isoeléctricos predominantemente ácidos en seis morfotipos [12D (Stachibotrys sp.), 22D, 29D, 64D, 66D y 73D (Penicillium spp.)], y la existencia de una unica isoforma de pI alcalino en los dos morfotipos restantes [8D (Chaetomium sp.), y 21D (Eladia saccula)]. En este punto se destacó la recuperación en la producción de proteasas extracelulares para el morfotipo 21D (E. Saccula ) con una actividad específica de 34.2Ueq/mg, y la "amplificación" en la producción de las mismas en los morfotipos 8D (Chaetomium sp.) y 12D (Stachybotrys sp.), con 15.22Ueq/mg y 13.65Ueq/mg, respectivamente, mediante la ejecución de cultivos líquidos diferenciales continuados (presión de producción).
La ocurrencia mayoritaria de isoformas con pIs ácidos encontrada en este trabajo concuerda con los resultados obtenidos por el equipo profesional de químicos del laboratorio de física, química y biología del Archivo de Bogotá, cuyas investigaciones (reporte interno) concluyen que los documentos que han sufrido envejecimiento acelerado como consecuencia de su exposición a agentes físicos como los rayos UV (simulación de documentos deteriorados) presentan una disminución gradual de pH hacia niveles ácidos, lo que finalmente favorece el desarrollo microbiano sobre los soportes, y consigo la producción de enzimas especificas que les permitan colonizar microambientes heterogéneos circundantes. De hecho, es igualmente conocido que los hongos en general tienden a acidificar los medios sobre los que se desarrollan mediante la acumulación de acido cítrico o acido oxálico a pH 2-6, y a controlar el nivel de acidez mediante la biosíntesis de de-carboxilasas para prevenir la acumulación de altas cantidades de acido oxálico, según sus necesidades fisiológicas (Kudryavtseva et al., 2008).
Sin embargo, el giro de este trabajo concomitante con la búsqueda de proteasas con características (por ejemplo pH optimo de actividad enzimática, estabilidad enzimática, efecto de quelante y surfactantes, entre otros) asociadas a la catálisis y remoción de proteínas residuales en ambientes alcalinos propios de los métodos de limpieza documental a manera de detergente, conllevo a la selección de las isoformas alcalinas producidas por los morfotipos 8D y 21D como candidatas para su purificación y caracterización parcial de forma económica y eficiente, en trabajos posteriores.
Conclusiones
El diseño experimental planteado en este trabajo ha conducido a la selección de dos morfotipos proteolíticos promisorios, 8D (Chaetomium sp.) y 21D (E. Saccula ), por cuanto fueron los únicos microorganismos que presentaron una isoforma alcalina "extrema" susceptible de ser purificada, caracterizada y empleada de forma económica y eficiente en la eliminación selectiva de proteínas residuales en ambientes alcalinos (a manera de detergente), apoyando las practicas locales de restauración y conservación del patrimonio cultural. Si bien este trabajo acorta la brecha existente entre las prácticas de restauración tradicionales y las nuevas tecnologías "limpias" basadas en la búsqueda y aplicación de productos biológicos altamente específicos y mínimamente intrusivos, tóxicos y contaminantes, abre igualmente una serie de posibilidades hacia la obtención de enzimas de la misma naturaleza o de diferente naturaleza con actividades hidrolíticas promisorias (por ejemplo amilasas, celulasas, hemicelulasas, lipasas, etc.) haciendo uso de la biodiversidad existente en nuestros sistema de archivos distritales.
Agradecimientos
Los autores agradecen la financiación y el soporte técnico otorgado a este trabajo en el marco del Convenio interinstitucional de cooperación por ciencia, tecnología e innovación No. 2215100-513-2009, celebrado entre la Secretaría General de la Alcaldía Mayor de Bogotá-Dirección Archivo de Bogotá - y el Centro Internacional de Física (C.I.F.), y en el marco del proyecto 12113 aprobado por la Dirección de Investigaciones Sede Bogotá de la Universidad Nacional de Colombia.
Referencias bibliográficas
1 Baraznenok, V. A., Becker, E. G., Ankudimova, N. V., Okunev, N. N. (1999) Characterization of neutral xylanases from Chaetomium cellulolyticum and their biobleaching effect on eucalyptus pulp. Enzyme and Microbial Technology. 25: 651. [ Links ]
2 Berner, M., Wanner, G., Luhitz, W. (1997) A comparative study of the fungal flora present in medieval wall paintings in the chapel of the Castle Herberstein and in the Parish Church of St Georgen in Styria, Austria. International Biodeterioration & Biodegradation. 40(1): 53-61. [ Links ]
3 Busconi, L., Fob, E., Martone, C., Trucco, R., Sanchez, J. (1984) Identification of two alkaline proteases and a trypsin inhibitor from muscle of white croaker (Micropogon opercularis). FEBS Letters. 176(1): 211-214. [ Links ]
4 Charney, J., Tomarelli, R. M. (1947) A colorimetric method for the determination of the proteolytic activity of duodenal juice. Journal of Biological Chemistry. 171(2): 501-505. [ Links ]
5 Castiñeyra, I., Ferrer, S., San Andres, M. (2004) Estudio de la especificidad de varios preparados enzimáticos de proteasas mediante SDS-PAGE. Departamento de pintura-restauración, Facultad de Bellas Artes. Universidad Complutense de Madrid. En el marco del proyecto 06/HSE/0045/2004 titulado "La utilización de enzimas proteolíticas en procesos de limpieza de obras de arte". [ Links ]
6 Cowan, D. A., Daniel, R. M. (1982) A modification for increasing the sensitivity of the casein-agar plate assay: a simple semiquantitative assay for thermophilic and mesophilic proteases. J Biochem Biophys Methods. 6: 31-7. [ Links ]
7 Cruz, C. A. (2011) Caracterización parcial de una proteasa alcalina a partir de hongos filamentosos implicados en el deterioro de documentos históricos. Tesis de Maestría para optar al título de Magister en Microbiología, Universidad Nacional de Colombia. p. 22. [ Links ]
8 Cruz, C. A. (2008) Purificación y caracterización parcial de proteasas alcalinas a partir de hongos filamentosos implicados en la degradación del patrimonio documental en el Archivo Distrital de Bogotá. Tesis de Pregrado. Universidad Militar Nueva Granada. Bogotá, Colombia. [ Links ]
9 Das, M. K. L., Prasad, J. S., Ahmad, S. K. (1997) Endoglucanase production by paper - degrading mycoflora. Letters in Applied Microbiology. 25: 313 - 315. [ Links ]
10 Decoux, S. (2002) Enzymes used for adhesive removal in paper conservation: a literature review. Journal of the Society of Archivists. 23(2): 187-195. [ Links ]
11 Dienes, D., Börjesson, J., Häagglund, P., Tjerneld, F., Lidén, G., Réczey, K., Stalbrand, H. (2007) Identification of a trypsin-like serine protease from Trichoderma reesei QM9414. Enzyme and Microbial Technology. 40: 1087-1094. [ Links ]
12 Florian, M. L., Manning, L. (2000) SEM analysis of irregular fungal fox spots in an 1854 book: population dynamics and species identification. International Biodeterioration & Biodegradation. 46: 205. [ Links ]
13 Garfin, D. E. (1990) Methods in Enzymology. Academic Press, San Diego, CA. Volume 182; Chapter 35. [ Links ]
14 Gorbushina, A., Heyrman, J., Dornieden, T., Gonzalez-Delvalle, M., Krumbein, W. E., Laiz, L., Petersen, K., Saiz-Jimenez, C., Swings, J. (2004) Bacterial and fungal diversity and biodeterioration problems in mural painting environments of St. Martins church (Greene-Kreiensen, Germany). International Biodeterioration & Biodegradation. 53: 13. [ Links ]
15 Gupta, R., Beg, Q. K., Khan, S., Chauhan, B. (2002) An overview on fermentation, downstream processing and properties of microbial alkaline proteases. Appl Microbiol Biotechnol. 60: 381-395. [ Links ]
16 Ho, W. C., Ko, W. H. (1997) A simple method for obtaining single-spore isolates of fungi. Bot. Bull. Acad. Sin. 38: 41-44. [ Links ]
17 Hueck, H. J. (2001) The biodeterioration of materials - An appraisal. International Biodeterioration & Biodegradation. 48: 5-11. [ Links ]
18 Iversen, S. L., Jorgensen, M. H. (1995) Azocasein assay for alkaline protease in complex fermentation broth. Biotechnology Techniques. 9(8): 573-576. [ Links ]
19 Iwashita, K. (2002) Recent studies of protein secretion by filamentous fungi. Journal of Bioscience and Bioengineering. 94(6): 530-535. [ Links ]
20 Jellison, J., C. Jasalavich. (2000) A review of selected methods for the detection of degradative fungi. International Biodeterioration & Biodegradation. 46: 241. [ Links ]
21 Kudryavtseva, O. A., Dunaevsky, Y. E., Kamzolkina, O. V., Belozersky, M. A. (2008) Fungal proteolytic enzymes: features of the extracellular proteases of xylotrophic Basidiomycetes. Microbiology. 77(6): 643-653. [ Links ]
22 Mateus, J., Peña, D., Peña G., Rojas, A., Rojas, J., Zambrano, S., Martinez, M., Florez, C., Santander, M. (2001) Seguimiento y control de biodeterioro microbiológico en documentos de interés histórico en el Archivo General de la Nación. Universitas Scientiarum, Revista de la Facultad de Ciencias, Pontificia Universidad Javeriana. 9(1): 37-46. [ Links ]
23 Mikán, J. F. (2001) Arabinoxylan degrading enzymes of the wheat pathogen Stagonospora nodorum. Thesis PhD. The University of Bath, U. K. [ Links ]
24 Monod, M., Capoccia, S., Lechenne, B., Zaugg, C., Holdom, M., Jousson, O. (2002) Secreted proteases from pathogenic fungi. Int. J. Med. Microbiol. 292: 405 - 419. [ Links ]
25 Pandey, A. (2003) Solid-state fermentation. Biochemical Engineering Journal. 13: 81-84. [ Links ]
26 Pedroza, A. M., Matiz, A., Gomez, D. (2001) Manual de laboratorio: Introducción a la biotecnología. Pontificia Universidad Javeriana. 27-32p. [ Links ]
27 Peyronel, D. V., Cantera, A. M. (1995) A simple and rapid technique for postelectrophoretic detection of proteases using azocasein. Electrophoresis. 16: 1894-1897. [ Links ]
28 Reichard, U., Cole, G. T., Hill, T. W., Rachel, R., Monod, M. (2000) Molecular characterization and influence on fungal development of ALP2, a novel serine proteinase from Aspergillus fumigatus. Int. J. Med. Microbiol. 290: 549 - 558. [ Links ]
29 Rojas, J. A. (2007) Caracterización isoenzimática de proteasas, amilasas y purificación parcial de proteasas de hongos filamentosos causantes de biodeterioro de papel industrial. Tesis de Maestría en Ciencias Biológicas. Universidad de los Andes. Bogotá, Colombia. [ Links ]
30 Rojas, J. A., Cruz, C. A., Mikán, J.F., Villalba, L. S., Cepero de Garcia, M. C., Restrepo, S. (2009) Isoenzyme characterization of proteases and amylases and partial purification of proteases from filamentous fungi causing biodeterioration of industrial paper. International Biodeterioration & Biodegradation. 63: 169-175. [ Links ]
31 Savitha, S. Sadhasivam, S., Swaminathan, K., Huei Lin, F. (2011) Fungal protease: Production, purification and compatibility with laundry detergents and their wash performance. Journal of the Taiwan Institute of Chemical Engineers. 42: 298-304. [ Links ]
32 Segal, J., Cooper, D. (1977) The use of enzymes to released adhesives. The Paper Conservator. 2: 47-50. [ Links ]
33 Shankar, S., Rao, M., Laxman, R. (2011) Purification and characterization of an alkaline protease by a new strain of Beauveria sp. Process Biochemistry. 46: 579-585. [ Links ]
34 Stoschek, C. M. (1990) Increased uniformity in the response of the Coomasie Blue G protein assay to different proteins. Analytical Biochemistry. 184: 111-116. [ Links ]
35 Szczepanowska, H., Cavaliere, A. (2000) Fungal deterioration of 18th and 19th century documents: a case study of the Tilghman Family Collection. International Biodeterioration & Biodegradation. 46: 245:249. [ Links ]
36 Vermelho, A., Meirelles, M., Lopes, A., Gonçalves, S., Chaia, A., Branquinha, M. (1996) Detection of extracellular proteases from microorganisms on agar plates. Mem Inst Oswaldo Cruz, Rio de Janeiro. 91(6): 755-760. [ Links ]
37 Vesterberg, O. (1990) Methods in Enzymology. Academic Press, San Diego, CA. Volume 182; Chapter 33. [ Links ]
38 Villalba, L. S., Mikan, J. F. (2005) Degradación del Archivo General de Colombia: actividades hidrolíticas y caracterización isoenzimática de poblaciones microbianas aisladas del patrimonio documental. Revista Internacional del Patrimonio Histórico. 3: 60-65. [ Links ]
39 Villalba, L. S., Mikan, J. F., Sanchez, J. (2004) Actividades hidrolíticas y caracterización isoenzimática de poblaciones microbianas aisladas del patrimonio documental del Archivo General de Colombia. Nova. 2(2): 50-58. [ Links ]
40 Wang, L., Ridgway, D., Gu, T., Moo-Young, M. (2005) Bioprocessing strategies to improve heterologous protein production in filamentous fungal fermentations. Biotechnology Advances. 23: 115-129. [ Links ]
41 Zyska, B. (1997) Fungi isolated from library materials: A review of the literature. International Biodeterioration & Biodegradation. 40(1): 43-5. [ Links ]

