










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol v.14 n.1 Bogotá jan./jun. 2012
ARTÍCULO CORTO
Caracterización de enzimas hidrolíticas de Aspergillus ficuum producidas en fermentación sólida sobre torta de canola
Characterization of Aspergillus ficuum hydrolytic enzymes produced in solid state fermentation of cold-pressed canola cake
Marcia Costa1, Marcelo Torres1 y Alejandro Reyes2.
1 Instituto de Ciencia y Tecnología de los Alimentos, Facultad de Ciencias Agrarias, Universidad Austral de Chile, Casilla 567, Valdivia, Chile mcosta@uach.cl
2 Instituto de Bioquímica y Microbiología, Facultad de Ciencias, Universidad Austral de Chile, Casilla 567, Valdivia, Chile
Recibido: abril 11 de 2008 Aprobado: octubre 23 de 2008
Resumen
En el presente trabajo, se describe la caracterización respecto al pH y temperatura de extractos semipurificados mediante ultrafiltración escalonada de las enzimas fitasa, celulasa y xilanasa de Aspergillus ficuum cepa DSM 932 producidos en fermentación en estado sólido (en adelante SSF) y usando torta de canola como sustrato. La fitasa presentó un pH y una temperatura óptima de 5.0 y 60°C respectivamente, en tanto que la celulasa presentó un pH óptimo de 7.0 con una temperatura óptima de 60°C y el extracto de xilanasa un pH óptimo de 5.4 y una temperatura óptima de 45°C.
Palabras clave: caracterización enzimas, fitasa, xilanasa, celulasa.
Abstract
This paper, describes the characterization respect to pH and temperature of semi-purified extracts by ultrafiltration step of the enzymes phytase, cellulase and xylanase produced by Aspergillus ficuum DSM 932 strain, in solid state fermentation (SSF) using cold-pressed canola cake as substrate. Phytase showed the optimal pH and temperature 6.0 and 60°C, respectively, while cellulase showed a pH optimal of 7.0 with an optimal temperature of 60°C and xylanase extract an optimal pH of 5.4 and an optimal temperature of 45°C.
Key words: Passiflora, competition, mycoparasitism, inhibition, antagonism.
Introducción
En los últimos años, se ha evidenciado un interés creciente por los procesos de SSF para desarrollar bio-procesos tales como tratamientos de biodegradación de compuestos peligrosos, desintoxicación biológica de residuos agroindustriales, descomposición de madera, enriquecimiento de residuos de cosecha, producción de alimentos con valor agregado, enzimas, ácidos orgánicos y metabolitos secundarios biológicamente activos, entre muchos otros.
Es así como en la SSF los microorganismos crecen sobre una matriz sólida en escasez ó ausencia de agua libre y se usa con gran éxito en la producción, entre otros, de antibióticos, micotoxinas, surfactantes, ácidos orgánicos, compuestos aromáticos, pesticidas y enzimas (Blandino et al., 2005; Krishna, 2005; De Ory et al., 2007).
La torta de canola es un subproducto de la industria productora de aceite comestible, tiene aplicación en la alimentación de rumiantes dada su composición de aproximadamente un 34% de proteínas, un 6% de ácido fítico, un 15% de fibra y un 26% de carbohidratos (Duvnjak y Nair, 1990). Este subproducto puede ser biotransformado mediante SSF en fuente de enzimas hidrolíticas de importancia comercial (fitasas, celulasas y xilanasas). Las fitasas son fosfatasas que pertenecen a la subfamilia de las fosfatasas ácidas histidina, las cuales catalizan la hidrólisis de las uniones fosfomonoester del fitato (sales de mio- inositol hexakisfosfato) ó del mio-inositol 1, 2, 3, 4, 5, 6-hexakisdihidrógeno fosfato (ácido fítico), produciendo derivados penta, tetra, tri, di y monofosfato del mioinositol y fosfato inorgánico (Pi) (Duvnjak y Al-Asheh, 1994; Krishna, 2005). Su uso principal es en la industria de alimentos, donde se emplea en forma de suplemento en piensos para animales monogástricos, reduciéndose los efectos contaminantes del fitato en las heces al poder acceder a una mayor biodisponibilidad de fosfato para sus necesidades fisiológicas (Roopesh et al., 2006; Walsh y Casey, 2003).
Celulasas es el nombre genérico que recibe un complejo de enzimas capaces de degradar celulosa y está conformado por la endo-1,4-b-D-glucanasa, que produce glucoligo-sacáridos; la exo-1,4-b-D-glucanasa (celobiohidrolasa), que produce celobiosa y la b-D-glucosidasa que degrada los gluco-oligosacáridos a glucosa. Se usa principalmente en las primeras etapas de la producción de glucosa para etanol y en el tratamiento de materiales que contienen lignocelulosa (Blandino et al., 2005; Krishna, 2005; De Vries y Visser, 2001; Yamamoto et al., 1995).
Por otra parte, xilanasas es el nombre genérico de una familia de enzimas que catalizan la ruptura de los enlaces xilosídicos internos de la molécula de heteroxilano (xilano), constituyente mayoritario de la hemicelulosa de la pared celular vegetal; siendo las endoxilanasas, que rompen el esqueleto de xilano en pequeños oligosacáridos y las b-xilosidasas, las que finalmente los degradan a moléculas de xilosa (Fang et al., 2008; Anthony et al., 2003; De Vries y Visser, 2001). Se usan en los procesos de clarificación de vinos y jugos, y en procesos de biodecoloración de pulpa de papel (Krishna, 2005; Zhaoxin et al., 2008).
La torta de canola biotransformada, una vez inactivado el A. ficuum, puede ser incorporada en dietas para monogástricos, ó las enzimas producidas pueden ser obtenidas y purificadas parcialmente, aplicando tecnologías de membrana a los extractos crudos obtenidos de la extracción sólido-líquido de la torta ya tratada (Costa et al., 2010).
Considerando los antecedentes antes señalados y para el caso de usar la torta de canola para producir extractos semipurificados de las enzimas fitasas, celulasas y xilanasas, se planteó como objetivo general de esta investigación caracterizar la actividad a diferentes pH y temperatura de extractos enzimáticos semipurificados de dichas enzimas obtenidos de la SSF con Aspergillus ficuum sobre torta de canola.
Materiales y métodos.
Cada uno de los diferentes extractos fue obtenido por duplicado, empleando los mismos protocolos desarrollados por Costa et al. (2010) y purificados parcialmente mediante ultrafiltración y diafiltración escalonada, tal como allí se indica. A modo de síntesis, la figura 1 presenta el proceso llevado a cabo con los extractos semipurificados a caracterizar. No se usaron los de menor tamaño por cuanto de acuerdo a lo realizado por Costa et al. (2010) la mayor actividad se presenta en las fracciones sobre 30 kDa para celulasas y xilanasas.
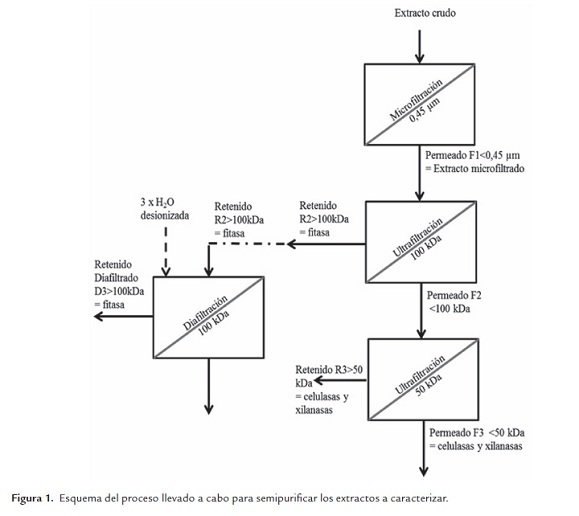
Para estos experimentos las SSF se trabajaron en bandejas de vidrio con canola a un ˜40% de humedad. Las bandejas fueron dispuestas semitapadas en estufas a 25°C con circulación de aire filtrado y saturado. Las fermentaciones fueron realizadas hasta el día en que se presentó la mayor actividad enzimática de cada enzima, a saber: 48 h para celulasa, 72 h para fitasa y 96 h para xilanasa, de acuerdo a lo reportado por Costa et al. (2010).
Las extractos crudos se obtuvieron con una solución de Triton X-100 al 0.1% a razón de 500 mL por cada 100 g de torta de canola fermentada, agitando a 35°C por 1 hora a 1000 rpm en un agitador de pedestal de velocidad variable de hélice (3 palas) (IKA EUROSTAR® digital), seguido de un prensado en muselina y de una centrifugación a 7.520 x G, 10 min a 4°C.
Las ultrafiltraciones y diafiltraciones (con agua desionizada) se realizaron a temperatura ambiente usando una celda Amicon de 50 mL con membranas Millipore de 100 kDa (60 rpm, 30 min) y 50 kDa (60 rpm, 90 min).
Medición de actividad enzimática
Para determinar la actividad fitásica se usó como sustrato ácido fítico (en forma de su sal dodecasódica, marca Sigma) y modificado de acuerdo a Harland y Harland (1980), donde se hace uso del reactivo de Taussky-Shorr, que permite detectar el fosfato inorgánico (Pi) luego de una reducción del ácido fosfomolíbdico por el sulfato ferroso, seguido de la formación de un complejo azul de Pi-molibdato reducido, cuya absorbancia se mide a 660 nm (Taussky y Shorr, 1953); el procedimiento se detalla en Costa et al. (2009).
Para determinar la actividad celulásica y xilanásica se utilizó el método propuesto por Miller (1959) modificado, donde los sustratos carboximetilcelulosa (marca Merck) o Birchwood xilano (marca Sigma) por acción de esas enzimas liberan azúcares los reductores glucosa y xilosa respectivamente, que son detectados mediante el reactivo DNS (ácido 3,5-dinitrosalicílico, color amarillo) que se reduce a ácido 3-amino-5-nitrosalicilico, midiéndose la absorbancia a 510 nm y cuyo procedimiento se detalla en Costa et al. (2010).
Cabe señalar que todas las mediciones de absorbancia se realizaron en un espectrofotómetro Spectronic® Genesys 5.
Caracterización de enzimas hidrolíticas respecto del pH y la temperatura.
Fitasa. Se usó el extracto correspondiente a un tercer diafiltrado (D3) de la membrana de 100 kDa. Para determinar el pH óptimo de la enzima se utilizó el protocolo descrito por Howson y Davis (1983) modificado. Las reacciones se hicieron a temperatura de trabajo constante (55°C) y la escala de pH usada fue de 2.0 a 7.5, con intervalos de 0.5 unidades. Los buffers usados para preparar las diluciones de la enzima y para la disolución de su sustrato (ácido fítico) fueron buffer Sorensen glicina I (glicina-HCl 0.1 M) (rango pH 2.0- 3.5), buffer acetato 0.2 M (rango pH 4.0-5.5) y buffer Sorensen citrato II (rango pH 6.0- 7.5).
Para determinar la temperatura óptima también se utilizó el método descrito por Howson y Davis (1983) modificado. Las reacciones se hicieron a pH de trabajo constante (pH = 5.15) y la escala de temperaturas usada fue de 30°C a 80 °C, con intervalos de 5°C.
Celulasa. Se usó el extracto correspondiente al cuarto ultrafiltrado (retenido y filtrado; R3 y F3 respectivamente) de la membrana de 50 kDa. Para determinar el pH óptimo de la enzima se utilizó el protocolo descrito por Yamamoto et al. (1995) modificado. Las reacciones se hicieron a temperatura de trabajo constante (50°C) y la escala de pH's usados fue de 3.0 a 11.0, con intervalos de 1.0 unidad. El buffer de trabajo usado para preparar las diluciones de la enzima y para la disolución de su sustrato (carboximetilcelulosa) fue glicina-NaOH 0.05 M (rango pH 9.0-11.0) y se equilibró con los siguientes buffers para alcanzar los restantes pH deseados: buffer citrato (para obtener un rango de pH 3.0-6.0) y buffer fosfato (para obtener un rango de pH 7.0-8.0).
Para determinar la temperatura óptima también se utilizó el método descrito por Yamamoto et al. (1995) modificado. Las reacciones se hicieron a pH de trabajo constante (9.0) y la escala de temperaturas usada fue de 45°C a 75 °C, con intervalos de 5°C.
Xilanasa. Se usó el extracto correspondiente al cuarto ultrafiltrado (retenido y filtrado; R3 y F3, respectivamente) de la membrana de 50 kDa. Para determinar el pH óptimo de la enzima se utilizó el protocolo descrito por Zhaoxin et al. (2008) modificado. Las reacciones se hicieron a temperatura de trabajo constante (50°C) y la escala de pH's usados fue de 3.0 a 7.0, con intervalos de 0.5 unidades. Para preparar las diluciones de la enzima y la disolución de su sustrato (Birchwood xilano) se usaron diversas disoluciones de buffer citrato 0.05 M que tenían los pH deseados.
Para determinar la temperatura óptima también se utilizó el método descrito por Zhaoxin et al. (2008) modificado. Las reacciones se hicieron a pH de trabajo constante (5.4) y la escala de temperaturas usada fue de 45°C a 75 °C, con intervalos de 5°C.
Resultados y discusión
Caracterización de los extractos enzimáticos respecto al pH y temperatura
- Fitasa
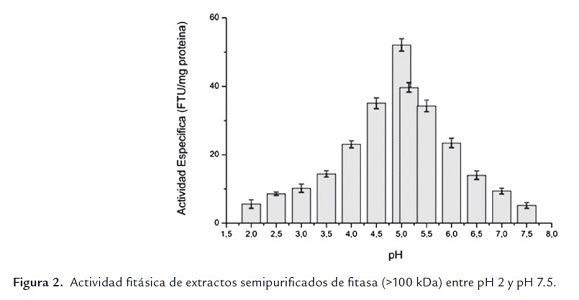
La figura 2 muestra los resultados obtenidos al caracterizar el extracto ultrafiltrado y diafiltrado (>100 kDa) de enzima fitasa (D3), se obtuvo un pH óptimo de 5.0 en las condiciones de trabajo realizadas. Dicho pH óptimo fue reportado también por Droráková et al. (1997) para una fitasa de Aspergillus niger; Duvnjak y Al-Asheh (1994) reportaron un pH óptimo de 4.7 para una fitasa de Aspergillus carbonarius NRC 401121, en tanto que Howson y Davis (1983) reportaron para una fitasa de Aspergillus ficuum NRRL 3135 (DSM 932) dos pH's óptimos, uno a 2.0 y otro a 5.5, aunque ellos explican posteriormente que el pH óptimo a 2.0 se debe a la acción de una fosfatasa ácida no específica que enmascara la actividad fitasa que a ese pH igual existe pero es muy baja y que el pH óptimo de 5.5 es el que corresponde a fitasa. Cabe destacar que la actividad enzimática en el pH óptimo es un 31% mayor que la obtenida al pH de trabajo (5.15) y que la enzima es estable en el rango de pH comprendido entre 4.5 y 5.5.
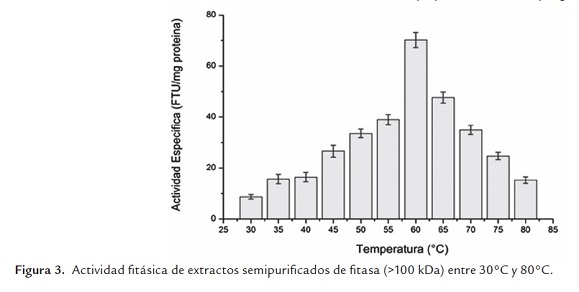
Por otra parte, la figura 3 muestra que la temperatura óptima de este extracto enzimático se alcanzó a los 60°C en las condiciones de trabajo realizadas. Dicha temperatura óptima es menor a la reportada por Walsh y Casey (2003) para la fitasa de Aspergillus niger, la que fue de 65°C, mayor a los 53°C reportado por Duvnjak y Al-Asheh (1994) para una fitasa de la cepa Aspergillus carbonarius NRC 401121, y los 55°C informado por Howson y Davis (1983) para una fitasa de Aspergillus ficuum NRRL 3135 (DSM 932). Es importante hacer notar que la actividad enzimática a la temperatura óptima es un 80% mayor que la obtenida a la temperatura de trabajo (55°C) y que la enzima es estable en el rango de temperaturas comprendidas entre 50°C y 70°C.
- Celulasas
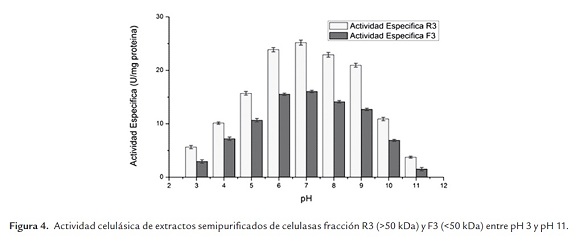
En la figura 4 es posible apreciar que para el extracto ultrafiltrado de celulasas (R3, retenido >50 kDa, y F3 permeado <50 kDa), se obtuvo un pH óptimo de 7.0 en las condiciones de trabajo realizadas. Dicho pH óptimo es también reportado por Yamamoto et al. (1995) quienes determinaron que el pH óptimo de una de las enzimas del complejo celulasa de Aspergillus niger IFO 31125 oscila entre 6.0 y 7.0. Es así como la actividad enzimática en el pH óptimo es un 20% mayor que la obtenida al pH de trabajo (9.0) y que la enzima es estable en el rango de pH 6.0 y pH 9.0.
Respecto de la temperatura óptima, en la figura 5 es posible observar que este extracto enzimático presentó una actividad óptima a los 60°C en las condiciones de trabajo realizadas. Dicha temperatura óptima es menor a la reportada por Yamamoto et al. (1995) quienes determinaron que una temperatura óptima de 70°C en una de las enzimas del complejo celulasa de Aspergillus niger IFO 31125. De lo anterior, resulta destacable que la actividad enzimática a la temperatura óptima es un 10% mayor que la obtenida a la temperatura de trabajo (50°C) y que la enzima es estable en el rango de temperaturas comprendidas entre 45°C y 65°C.
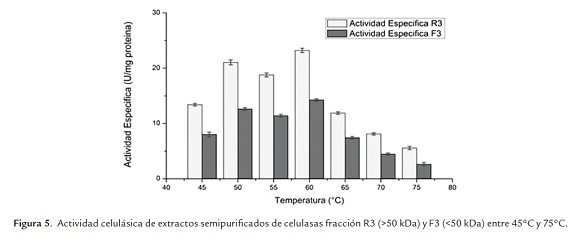
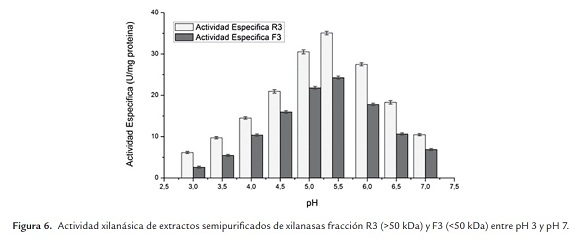
- Xilanasas
La figura 6 muestra los resultados de la caracterización del extracto de xilanasa respecto del pH donde es posible apreciar que para el extracto ultrafiltrado de xilanasa (R3, retenido >50 kDa y F3, permeado <50 kDa), se obtuvo un pH óptimo de 5.4 en las condiciones de trabajo realizadas. Dicho pH óptimo es mayor al reportado por Zhaoxin et al. (2008) quienes determinaron un pH óptimo de 5.0 en una de las enzimas del complejo xilanasa de Aspergillus ficuum AF-98, y menor al pH 6.0 reportado por Fang et al. (2008) quienes determinaron óptimo en una de las enzimas del complejo xilanasa de Aspergillus carneus M 34. De lo anterior entonces se destaca que la actividad enzimática en el pH óptimo correspondió al pH de trabajo (5.4) y que la enzima es estable en el rango de pH's comprendidos entre 5.0 y 6.0.
Por otra parte, la figura 7 muestra que la temperatura óptima de este extracto enzimático de xilanasa se alcanzó a los 45°C en las condiciones de trabajo realizadas. Dicha temperatura óptima es reportada por Zhaoxin et al. (2008) para una de las enzimas del complejo xilanasa de Aspergillus ficuum AF-98. Por su parte, Fang et al. (2008) reportaron 50°C como temperatura óptima de una de las enzimas del complejo xilanasa de Aspergillus carneus M 34. Cabe destacar que la actividad enzimática de xilanasa a la temperatura óptima es un 10% mayor que la obtenida a la temperatura de trabajo (50°C) y que la enzima es estable en el rango de temperaturas comprendidas entre 40°C y 55°C.
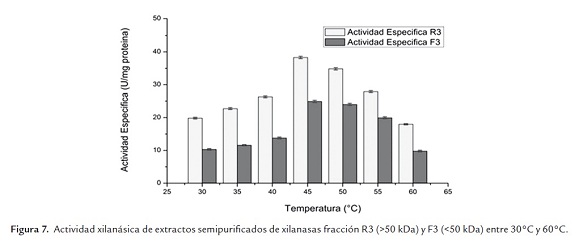
Conclusiones
Los diferentes extractos enzimáticos producidos en SSF de Aspergillus ficuum cepa DSM 932 en torta de canola y purificados parcialmente mediante ultrafiltración escalonada, presentan para fitasa un pH óptimo de 5.0 y una temperatura óptima de 60°C, en tanto que el complejo celulasas muestra un pH óptimo de 7.0 y una temperatura óptima de 60°C y el complejo xilanasas muestra un pH óptimo de 5.4 y una temperatura óptima de 45°C.
Agradecimientos
Este trabajo fue financiado por la Dirección de la Investigación de la Vicerrectoría Académica de la Universidad Austral de Chile, Proyecto DID-UACH S-2008-54.
Bibliografía
1 Anthony T., Chandra Raj K., Rajendran A., Gunasekaran P. 2003. High molecular weight cellulase - free xylanase from alkali - tolerant Aspergillus fumigatus AR 1. Enzyme and Microbial Technology. 32: 647-654. [ Links ]
2 Blandino A., Botella C., De Ory I., Webb C., Cantero D. 2005. Hydrolytic enzyme production by Aspergillus awamori on grape pomace. Biochemical Engineering Journal. 26: 100-106. [ Links ]
3 Costa M., Lerchundi G., Villarroel F., Torres M., Schöbitz R. 2009. Producción de enzima fitasa de Aspergillus ficuum con residuos agroindustriales en fermentación sumergida y sobre sustrato sólido. Rev. Colomb. Biotecnol. 11(1): 73-83. [ Links ]
4 Costa M., Torres M., Magariños H., Reyes A. 2010. Producción y purificación parcial de enzimas hidrolíticas de Aspergillus ficuum en fermentación sólida sobre residuos agroindustriales. Rev. Colomb. Biotecnol. 11(2):163-175. [ Links ]
5 De Ory I., Caro I., Díaz A. B., Blandino A. 2007. Evaluation of the conditions for the extraction of hydrolytic enzymes obtained by solid state fermentation from grape pomace. Enzyme and Microbial Technology. 41: 302-306. [ Links ]
6 De Vries R. P., Visser J. 2001. Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiol. Mol. Biol. Rev. 65 (4): 497-522. [ Links ]
7 Droráková J., Volfová O., Kopecký J. 1997. Characterization of phytase produced by Aspergillus niger. Folia Microbiologica. 42 (4): 349-352. [ Links ]
8 Duvnjak Z., Al - Asheh S. 1994. Characteristics of phytase produced by Aspergillus carbonarius NRC 401121 in canola meal. Acta Biotechnologica. 14 (3): 223 - 233. [ Links ]
9 Duvnjak Z., Nair V.C. 1990. Reduction of phytic acid content in canola meal by Aspergillus ficuum in solid state fermentation process. Appl. Microbiol. Biotechnol. 34: 183-188. [ Links ]
10 Fang H-Y., Chang S-M., Lan C-H., Fang T.J. 2008. Purification and characterization of a xylanase from Aspergillus carneus M 34 and its potential use in photoprotectant preparation. Process Biochemistry. 43: 49-55. [ Links ]
11 Harland B.F., Harland J. 1980. Fermentative reduction of phytase in rye, white, and whole wheat breads. Cereal Chem. 57 (3): 226-229. [ Links ]
12 Howson S.J., Davis R.P. 1983. Production of phytate - hydrolyzing enzyme by some fungi. Enzyme Microbiol. Technol. 5: 377-382. [ Links ]
13 Krishna C. 2005. Solid State Fermentation Systems - An Overview. Critical Reviews in Biotechnology. 25 (1-2): 1-30. [ Links ]
14 Lowry O.H., Rosebrough N.J., Farr A.L., Randall R.J. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193: 265 - 275. [ Links ]
15 Miller G. L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry. 31 (3): 426-428. [ Links ]
16 Roopesh, K., Ramachandran S., Nampoothiri K.M., Szakacs G., Pandey, A. 2006. Comparison of phytase production on wheat bran and oilcakes in solid state fermentation by Mucor racemosus. Bioresource Technology. 97: 506-511. [ Links ]
17 Taussky H., Shorr E. 1953. A microcolorimetric method for the determination of inorganic phosphorus. Journal of Biological Chemistry. 202 (2): 675-685. [ Links ]
18 Walsh G., Casey, A. 2003. Purification and characterization of extracellular phytase from Aspergillus niger ATCC 9142. Bioresource Technology. 86: 183 - 188. [ Links ]
19 Yamamoto K., Akiba S., Kimura Y., Kumagai H. 1995. Purification and characterization of a protease - resistant cellulase from Aspergillus niger. J. Ferment. Bioeng. 79 (2): 125-130. [ Links ]
20 Zhaoxin L., Fengxia L., Mei L., Xiaomei B., Haizhen Z., Yi W. 2008. Purification and characterization of xylanase from Aspergillus ficuum AF - 98. Bioresource Technology. 99: 5938 - 5941. [ Links ]

