










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.14 no.2 Bogotá jul./Dec. 2012
ARTÍCULO DE INVESTIGACIÓN
Embriogénesis somática y producción de callo embriogénico friable de dos cultivares de yuca (Manihot esculenta Crantz)
Somatic Embryogenesis and friable embryogenic callus production in two cassava cultivars (Manihot esculenta Crantz)
Juan Camilo Ochoa1 , Paul Chavarriaga2 , Camilo López3 .
1 BSc., MSc. Laboratorio Biología Molecular. Departamento de Biología. Universidad Nacional de Colombia, sede Bogotá.
2 BSc., MSc. Centro Internacional de Agricultura Tropical (CIAT). Biotechnology Research Unit.
3 BSc., MSc., PhD. Profesor Asociado. Laboratorio Biología Molecular. Departamento de Biología. Universidad Nacional de Colombia, sede Bogotá. celopezc@unal.edu.co
Recibido: agosto 30 de 2011 Aprobado: noviembre 30 de 2012
Resumen
La yuca (Manihot esculenta Crantz) es un cultivo de alta importancia en países tropicales. La transformación genética de yuca ha sido posible desde hace 15 años mediante la producción que callo embriogénico friable (CEF) a partir de embriones somáticos. En el presente trabajo se evalúan la inducción de embriones somáticos usando tres diferentes auxinas sintéticas y la producción de CEF a partir de éstos en los cultivares de yuca SG107-35 y BRA685. Estos cultivares son resistentes a la bacteriosis vascular de yuca cuyo agente causal es Xanthomonas axonopodis pv. manihotis, una de las principales limitantes del cultivo. Los resultados obtenidos muestran que en ambos cultivares la hormona Picloram a una concentración de 12 mg/l fue más eficiente que 2,4-D y Dicamba para producir embriones somáticos. Adicionalmente se consiguió la producción de CEF y la regeneración de plantas mediante embriogénesis somática en el cultivar BRA685. Los resultados del presente trabajo son importantes para evaluar la transformabilidad de distintos cultivares de yuca. Actualmente este número es bastante reducido principalmente porque la producción de CEF es fuertemente influenciada por el genotipo. Por tal razón solo se transforma de manera rutinaria y eficiente en el cultivar 60444. La posibilidad de transformación de distintos cultivares de yuca permitirá explotar la enorme variabilidad del cultivo, invitándonos a aumentar los esfuerzos para mejorar y universalizar los protocolos de transformación de yuca.
Palabras clave: Picloram, regeneración, dependencia del genotipo.
Abstract
The cassava (Manihot esculenta Crantz) crop has a very important role as a food, feed and a raw material in developing countries; therefore it is a priority to develop technologies oriented to the solution of problems and agronomic improvement of the crop. The genetic transformation of cassava was developed 15 years ago by producing friable embryogenic callus (FEC) from somatic embryos as target tissue for transformation. In the present work we evaluated the induction of somatic embryos by using three different synthetic auxins and the production of FEC from both SG107-35 and BRA685 cassava cultivars; both are resistant to cassava bacterial blight caused by Xanthomonas axonopodis pv. manihotis, the most important bacterial disease affecting the crop. Our results showed that in both cultivars gave rise to somatic embryos in media containing Picloram at a concentration of 12 mg/l being more efficient than using 2,4-D or Dicamba. Additionally the cultivar BRA685 produced regenerative FEC giving rise to plants through somatic embryogenesis. However compared to the model cultivar 60444, FEC production was greatly lower. This work shows new efforts to increase the number of transformable cultivars of cassava and take advantage of the enormous genetic variability of the crop.
Key words:Picloram, regeneration, genotype dependence.
Introducción
La yuca, Manihot esculenta Crantz es una planta dicotiledónea de la familia Euphorbiaceae con reproducción sexual monoica, hábito arbustivo, simpodial y raíces capaces de almacenar grandes cantidades de carbohidratos (Cadavid, 2005). Este cultivo es de gran importancia en países tropicales donde se siembra en alturas que van desde 0 hasta 1800 msnm (Ceballos, 2002). A nivel mundial la yuca se sitúa como el cuarto cultivo de mayor importancia como fuente de calorías y como componente básico de la alimentación de alrededor de 1000 millones de personas en el mundo (FAO, 1998). Entre los principales usos de la yuca se destacan el consumo de hojas y raíces por parte de animales y humanos, la extracción de almidón para procesos industriales y la producción de etanol para su potencial uso como biocombustibles (Ceballos, 2002). Adicionalmente, la yuca se destaca por su alta tolerancia a la sequía, capacidad de producción en suelos ácidos y degradados típicos de algunas regiones de países tropicales y la resistencia a diversas plagas y enfermedades (Cadavid, 2005).
A diferencia de otros cultivos importantes como el maíz, arroz y el trigo, la yuca ha sido poco estudiada comparativamente en términos de fitomejoramiento debido a algunas de sus características genéticas y reproductivas. Entre las principales se encuentran su largo ciclo de vida, la alta heterocigocidad, las bajas tasas de reproducción sexual por el bajo número de semillas fértiles que produce y la multiplicación asexual (Ceballos, 2002). La amplia utilización industrial de la yuca se ha visto limitada por la presencia de glucósidos cianogénicos que le pueden conferir determinados niveles de toxicidad, la falta de cultivares específicos para sus distintos usos y la poca duración postcosecha de las raíces tuberosas, las que no superan los siete días (Ceballos, 2002).
La transformación genética es una herramienta fundamental que permite acelerar el mejoramiento genético con miras a obtener productividad y rendimientos mayores. Además de esto la introducción de genes de resistencia a insectos y a herbicidas se ha constituido en uno de los principales medios para incrementar la producción y reducir los costos de producción y el empleo de insecticidas que comprometen la salud y el ambiente. El empleo de este tipo de estrategia permite superar varios de los obstáculos mencionados en el mejoramiento de la yuca.
La transformación genética de la yuca ha sido posible desde el año 1996 mediante la aplicación de dos protocolos distintos. El primero es mediado por Agrobacterium tumefaciens y utiliza cotiledones de embriones somáticos maduros producidos a partir de lóbulos de hojas jóvenes, provenientes de plantas in vitro, seguida de la regeneración bajo selección por medio de organogénesis (Li et al., 1996). El segundo de los métodos reportados, también en el año 1996, se realizó a partir de suspensiones de callo embriogénico friable (CEF) los cuales se transformaron mediante biobalística y posteriormente la fase de regeneración se realizó mediante embriogénesis somática (Schopke et al., 1996; Taylor et al., 1996). Estudios posteriores mostraron la posibilidad de transformar CEF usando A. tumefaciens para aumentar la efectividad de producción de plantas transgénicas de yuca (González et al., 1998).
Una vez desarrollados los protocolos de transformación se ha ensayado la incorporación de rasgos de interés agronómico en este cultivo tales como resistencia a herbicidas (Sarria et al., 2000), reducción de compuestos cianogénicos (Jørgensen et al., 2005; Siritunga et al., 2004; Siritunga y Sayre, 2003), aumento en el contenido y calidad del almidón almacenado en raíz (Ihemere et al., 2006; Raemakers et al., 2005) entre otros.
A pesar de lo anterior, la transformación genética de la yuca presenta todavia limitantes debido a que los procesos de embriogénesis somática, producción de CEF y regeneración son cultivar dependientes y la transformación solo ha sido exitosa en cultivares específicos (Raemakers et al., 2001). Además los procesos de transformación mencionados con anterioridad presentan algunas desventajas (Bull et al., 2009). Por ejemplo, el método basado en transformación de cotiledones y regeneración por organogénesis a pesar de ser más corto presenta mayor número de plantas regeneradas no transgénicas y la producción de plantas quiméricas (Bull et al., 2009). Por el contrario el segundo método tiene un número menor de plantas regeneradas no transgénicas y quimeras pero requiere de cerca del doble de tiempo que el primero (Bull et al., 2009).
Por las razones mencionadas, se hace necesario trabajar en mejorar los protocolos de transformación y ampliar el número de cultivares transformables. Antes de iniciar la transformación de nuevos cultivares de yuca es necesario estandarizar procesos que permitan la producción eficiente de embriones somáticos y CEF. Actualmente esto solo es posible con el cultivar 60444, por lo cual se ha convertido en un modelo de cultivo de tejidos y transformación genética. Previos estudios con cultivares de yuca provenientes de diferentes países han pretendido mejorar la producción de embriones somáticos con fines de certificación de material vegetal y transformación. Los estudios han evaluado distintos tipos de explantes y auxinas sintéticas a diferentes concentraciones (Feitosa et al., 2007; Mathews et al., 1993; Medero et al., 2000; Medina et al., 2003; Sofiari et al., 1997). Estos estudios encontraron que los meristemos axilares y apicales son capaces de producir eficientemente embriones somáticos (Medero et al., 2000), también se ha determinado que las auxinas sintéticas Picloram y Ácido 2,4-Diclorofenoxi acético (2,4-D) son altamente eficientes para la inducción de embriones somáticos. Sin embargo se han encontrado cultivares específicos en los cuales solo se inducen embriones somáticos con auxinas diferentes como Dicamba (Ácido 3,6-Dicloro-o-anísico) (Medina et al., 2003). De igual manera se han realizado ensayos sobre la producción de CEF con miras a la transformación genética de distintos cultivares de yuca encontrándose que la respuesta se restringe a un número reducido de cultivares; esto requiere le realización de evaluaciones sistemáticas de cultivares con mayor interés comercial o investigativo que 60444 (Hankoua et al., 2006; Taylor et al., 2001).
Los cultivares de yuca BRA685 y SG107-35 presentan altos niveles de resistencia a la bacteriosis vascular, una de las principales limitantes bióticas en la producción de yuca (Restrepo et al., 2000). Esta enfermedad es producida por la bacteria Xanthomonas axonopodis pv. manihotis y actualmente el mejor método de control consiste en el uso de cultivares resistentes y la aplicación de buenas prácticas agronómicas (Cadavid, 2005). En consecuencia la posibilidad de transformar estos cultivares se convierte en un paso imprescindible con miras a validar la función de posibles genes de resistencia mediante el empleo de estrategias de silenciamiento génico mediado por transgénesis, ya sea por medio de microARNs artificiales o el empleo de construcciones sentido-antisentido. Aprovechando la ventaja que para el empleo de estas estrategias no se requiere la identificación y clonación del gen completo. Una vez confirmada la función de estos genes mediante silenciamiento génico en variedades resistentes, se puede proceder a identificar el gen completo e introducirlo en variedades comerciales susceptibles para mejorar el control de la enfermedad y la productividad del cultivo.
El objetivo del presente trabajo consiste en evaluar los protocolos existentes para producción de embriones somáticos y callo embriogénico friable en dos variedades de yuca resistentes a bacteriosis vascular con miras a la validación funcional de genes implicados en la resistencia por medio de silenciamiento.
Materiales y métodos
Material vegetal
Plantas in vitro de los cultivares 60444, BRA685 y SG107-35 fueron obtenidas del banco de germoplasma del Centro Internacional de Agricultura Tropical (CIAT). Estas plantas fueron propagadas mediante segmentos nodales en el medio 4E que contiene contiene sales Murashige y Skoog (MS), sacarosa 2%, Tiamina 1ppm, Mio-inositol 100ppm, CuSO4 2µM, ANA 0.02ppm, GA3 0.05ppm y BAP 0.04ppm (Roca, 1984). Las plantas propagadas se mantuvieron a 28°C con un fotoperiodo de 12 horas luz y 12 horas de oscuridad en una incubadora Sanyo MLR 350.
Embriogénesis somática
La embriogénesis primaria se realizó a partir de yemas axilares en medio líquido MS250pi que consta de sales MS suplementado con vitaminas B5 de Gamborg, sacarosa 2%, CuSO4 2µM, caseína hidrolizada 50mg/l y Picloram 12mg/l o 2,4D 8mg/l o Dicamba 10mg/l de acuerdo al experimento (Beltrán et al., 2010). Las yemas axilares se incubaron a 28°C durante 26 días en oscuridad. Posterior a la embriogénesis primaria se realizó una embriogénesis cíclica en medio MS250pi endurecido con gelrite 0.2% partiendo de los embriones somáticos obtenidos en la embriogénesis primaria. Los embriones se incubaron por 30 días a 28°C en oscuridad.
Producción de callo embriogénico friable
Para la inducción del callo embriogénico friable (CEF) se aislaron grupos de embriones globulares cortados provenientes de la embriogénesis cíclica en fragmentos de cerca de 3mm2 y se transfirieron a medio GD250pi (Sales Greshoff y Doy (GD), Vitaminas GD, Sacarosa 2%, Picloram 50µM, Agar 0.6%) por 45 días a 28°C y fotoperiodo 12/12 (Bull et al., 2009). A partir de los clúster celulares obtenidos se realizó el aislamiento de CEF en medio GD250pi y se renovó el medio a los 22 días. El CEF aislado se incubó a 28°C con un fotoperiodo 12/12.
Regeneración de plantas
El CEF obtenido a partir del cultivar BRA685 se transfirió al medio de organogénesis reportado por Li et al., (1996) el cual conteine sales MS suplementados con vitaminas MS, sacarosa 2%, CuSO4 2µM, BAP 1ppm e IBA 0.5ppm y fue endurecido con gelrite 0.23% para la formación y maduración de embriones durante 60 días a 28°C y fotoperiodo 12/12. Los embriones fueron transferidos posteriormente a medio compuesto con sales MS, Vitaminas MS, sacarosa 2%, carbón activado 0.5% y gelrite 0.23% (Beltrán, et al.,2010). Estos se incubaron a 28°C con fotoperiodo 12/12 durante 45 días. Finalmente los ápices de los embriones germinados fueron enraizados en medio 17N el cual contiene 1/3 de sales MS, Vitaminas MS, ANA 0.01 mg/l y GA3 0.01mg/l (Beltrán et al., 2010).
Resultados
Embriogénesis somática primaria y embriogénesis cíclica
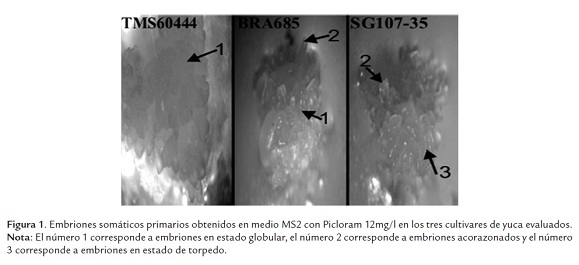
Inicialmente se evaluó el porcentaje de yemas axilares que formaron embriones somáticos en medio líquido de inducción de embriones somáticos suplementado con Picloram 12mg/l en los cultivares bajo estudio. El porcentaje de yemas axilares que produjeron embriones somáticos en los tres cultivares fue mayor del 60% (tabla 1). Sin embargo en el caso de SG107-35 y BRA685 se observó que los embriones maduraron prematuramente dado que se encontraron en estados acorazonados y de torpedo (figura 1). Esto no sucedió en el cultivar 60444, el cual fue utilizado como control, y en el que la mayoría de los embriones fueron globulares (figura 2).
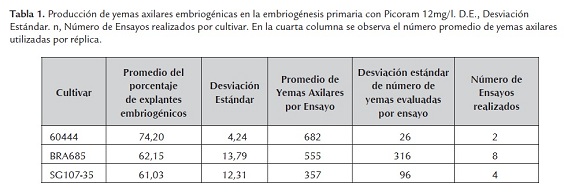
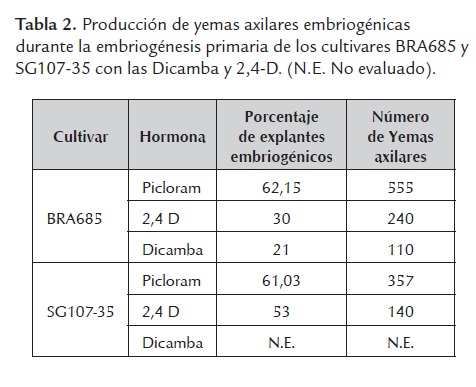
En concordancia con lo anterior la tendencia de maduración embriogénica prematura se mantuvo durante la embriogénesis cíclica. Por tal razón se evaluaron las hormonas 2,4-D a una concentración de 8mg/l y Dicamba a una concentración de 10mg/ durante los procesos de embriogénesis somática primaria y embriogénesis cíclica. Bajo las concentraciones hormonales mencionadas se observó una reducción del porcentaje de embriones producidos, particularmente en el cultivar BRA685 en el cual se redujo el porcentaje de yemas axilares embriogénicas a 30% con la hormona 2,4-D y a 21% con la hormona Dicamba (tabla 2). Adicionalmente la calidad de los embriones somáticos producidos con estas hormonas fue menor dado que se observó mayor cantidad de callo friable no embriogénico y los clústeres de embriones dentro de este eran de menor tamaño.
Producción de callo embriogénico friable
Los embriones globulares y acorazonados obtenidos a partir de los tres cultivares mencionados anteriormente fueron utilizados para inducir callo embriogénico friable en medio GD250pi sólido. Para ello estos fueron fraccionados y dispuestos sobre el medio organizados en clústeres celulares. En comparación con el cultivar control 60444 el comportamiento del cultivar BRA685 fue similar en términos macroscópicos dado que se observó crecimiento del clúster celular y una coloración amarillo claro. Por el contrario los clústeres celulares del cultivar SG107-35 presentaron un crecimiento reducido y una coloración parda típica de la producción de compuestos fenólicos indicativos de muerte celular.
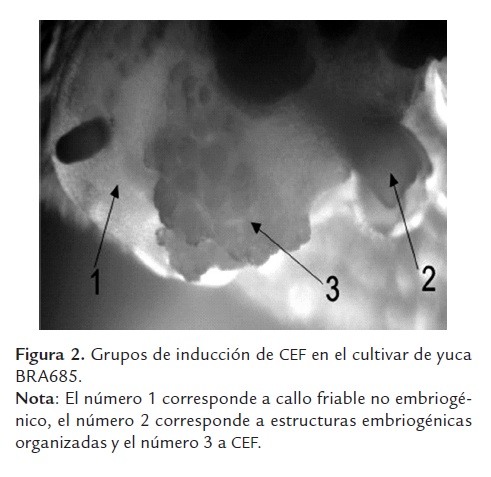
Microscópicamente se observó la producción de los cuatro tipos de grupos celulares descritos por Taylor et al.,(1996) en los cultivares 60444 y BRA685: i) estructuras embriogénicas organizadas, ii) callo friable no embriogénico, iii) callo marrón rígido (en inglés Hard Brown Callus) y iv) CEF (Fig 2). Por el contrario en el cultivar SG107-35 se observó únicamente la formación de los tres primeros grupos celulares. En la figura 2 se observa CEF que logró inducirse en el cultivar BRA685, sin embargo las cantidades que se produjeron comparadas con las producidas en el cultivar 60444 son bastante inferiores en más de un 90%. Adicionalmente, una vez aislado el CEF de los clústeres celulares solo el del cultivar 60444 fue capaz de proliferar y micropropagarse, por el contrario el CEF obtenido de BRA685 no produjo más cantidades de CEF que las que se obtuvieron en el aislamiento desde la inducción.
Regeneración de CEF de BRA685
A pesar de que el CEF producido del cultivar BRA685 no pudo proliferar una vez aislado este si fue capaz de inducir embriones somáticos y regenerar plantas cuyo aspecto no mostró signos de variación somaclonal al igual que plantas producidas a partir del CEF obtenido en 60444 a nivel in vitro (Fig. 3). Sin embargo dadas las pequeñas cantidades de CEF obtenidas no fue posible calcular un porcentaje de regeneración comparable con 60444.

Discusión
Dado que el cultivo de yuca es considerado un cultivo multipropósito, implica que es necesario desarrollar variedades específicas a regiones agroecológicas con características particulares y con rendimiento óptimo dependiendo del objetivo con el que se va a emplear. Igualmente, en términos investigativos, es de vital importancia el desarrollo de métodos que permitan explotar la enorme variabilidad genética que posee la yuca (Fregene y Puonti-Kaerlas, 2002). Una de las grandes bondades de los procesos de transformación genética es el hecho de poder realizar análisis funcionales o modificaciones sin la necesidad de cambiar el acervo genético de una planta en estudio que pudo ser obtenida por procesos intensivos de fitomejoramiento o en cultivares silvestres con alguna característica de interés.
Actualmente en yuca solo el cultivar 60444 se ha logrado transformar rutinariamente con protocolos iguales o similares al descrito por Bull et al. (2009). Por tal razón es necesario realizar estudios que permitan que la transformación de yuca pueda ser transferida a un número más amplio de cultivares ya sea mediante la generación de nuevos protocolos de trasformación, la modificación de aquellos existentes o la evaluación de estos en variedades de interés sobre las cuales no se ha evaluado el proceso de transformación.
La base inicial de producción de CEF con fines de transformación genética es la embriogénesis somática. Por tal razón el punto de partida para determinar la posibilidad de transformación de plantas de yuca consiste en tener una producción eficiente de embriones somáticos. Diversos estudios han evaluado varias auxinas sintéticas con fines de producción de embriones somáticos y han encontrado que Picloram y 2,4-D son las hormonas que presentan la mayor eficiencia con valores superiores al 50% (Groll et al., 2001; Medero et al., 2000; Medina et al., 2003; Raemakers et al., 1993). En este estudio se encontró que la hormona Picloram es más eficiente para producir embriones somáticos en BRA685 y SG107-35 que otras hormonas como 2,4-D o Dicamba. Sin embargo la alta heterocigocidad de la yuca implica que se observen diversas respuestas a nivel de cultivo de tejidos, por tal razón se hace necesario realizar modificaciones a los protocolos para que puedan ser adaptados a otros cultivares. Un ejemplo claro de esto ocurre en el cultivar "palomita", proveniente de Argentina, el cual solo induce embriones somáticos con la hormona Dicamba y no con Picloram o 2,4-D (Medero et al., 2000).
Trabajos previos han evaluado la posibilidad de producción y transformación CEF de cultivares alternativos de yuca (Hankoua et al., 2006; López, 2000; Taylor et al., 2001). Sin embargo la capacidad de transformación es mucho más reducida principalmente debido a la posibilidad de producción de líneas de CEF y regeneración del mismo. Taylor et al. (2001) evaluaron en 20 cultivares distintos de yuca la posibilidad de transformación genética de CEF, sin embargo en cinco de estos cultivares fue imposible producir CEF. De los 15 cultivares restantes no fue posible generar suspensiones embriogénicas con proliferación autónoma y solo en ocho fue posible la regeneración de plantas a partir del CEF producido en las suspensiones celulares (Taylor et al., 2001). De estos ocho cultivares solo tres pudieron ser transformados exitosamente. Hankoua et al. (2006) evaluaron cinco cultivares africanos distintos a 60444, de los cuales solo dos produjeron CEF, pero ninguno logró ser transformado porque no se lograron producir líneas de CEF que proliferaran de forma autónoma. En el presente estudio se encontró que en uno de los cultivares evaluados, BRA685, es posible la producción de CEF y regeneración de plantas a partir del mismo, sin embargo la reducida cantidad obtenida hace necesario incrementar esfuerzos que permitan mejorar y optimizar el procedimiento para que sea posible realizar transformaciones eficientes y se pueda explotar su característica de resistencia a bacteriosis vascular.
Uno de los principales limitantes para realizar mejoras a los protocolos de transformación, es la falta de información respecto a las variables que pueden influir en una respuesta determinada a nivel de cultivo de tejidos vegetales. Como se observa en la transformación de yuca, existen múltiples etapas tanto en la producción de tejido a transformar como en regenerarlo un vez transformado. En consecuencia esto hace que no pueda ser implementada exitosamente en otros cultivares y por ende que haya limitaciones en explotar la enorme variabilidad genética del cultivo.
Conclusiones
El presente trabajo constituye la etapa inicial para desarrollar protocolos de transformación en nuevos cultivares de yuca que permitan explotar las características de interés que presentan distintos cultivares. Adicionalmente se espera que el desarrollo biotecnológico del cultivo pueda ser transferido a los cultivadores para aumentar su rentabilidad, teniendo en cuenta la importancia que este tiene en países tropicales.
Agradecimientos
Los autores agradecen a todos los integrantes del grupo Manihot biotec por el constante apoyo. Este proyecto fue financiado por Colciencias y el Ministerio de Agricultura y Desarrollo Rural (MADR).
Referencias bibliográficas
1 Beltrán J., Prías M., Al-Babili S., Ladino J., López D., Beyer P., Chavarriaga P., Tohme J.. 2010. Expression pattern conferred by a glutamic acid-rich protein gene promoter in Weld-grown transgenic cassava (Manihot esculenta Crantz). Planta 231: 1413-1424. [ Links ]
2 Bull S. E., Owiti J.A., Niklaus M., Beeching J., Gruissem W., Vanderschuren H. 2009. Agrobacterium-mediated transformation of friable embryogenic calli and regeneration of transgenic cassava. Nat Protoc. 4 (12): 1845-54. [ Links ]
3 Cadavid L. F. 2005. Produccion de Yuca. Cali. CIAT. [ Links ]
4 Ceballos H. 2002. La yuca en Colombia y el mundo: nuevas perspectivas para un cultivo milenario. En: CIAT (eds.). La yuca en el Tercer Milenio: Sistemas modernos de producción, procesamiento, utilización y comercialización. 586. [ Links ]
5 FAO. 1998. Food and Agriculture Organization of the United Nations (FAO). FAO, Roma. [ Links ]
6 Feitosa T., Pinheiro J., Aguiar L., Jucá T., Paiva F. 2007. Somatic Embryogenesis in Cassava Genotypes from the Northeast of Brazil. Brazilian Archives of Biology and Technology. 50 (2): 201-206. [ Links ]
7 Fregene M., Puonti-Kaerlas J. 2002. Cassava Biotechnology. CAB International. 10. 179-207. [ Links ]
8 González A. E., Schöpke C., Taylor N., Beachy R., Fauquet C. 1998. Regeneration of transgenic cassava plants (Manihot esculenta Crantz) through Agrobacterium-mediated transformation of embryogenic suspension cultures. Plant Cell Reports. 17 827-831. [ Links ]
9 Groll J., Mycock D., Gray V,. Laminski, S. 2001. Secondary somatic embryogenesis of cassava on Picloram supplemented media. Plant Cell, Tissue and Organ Culture. 65 201-210. [ Links ]
10 Hankoua B. B., Taylor N., Ng S. Y. C, Fawole I., Puonti-Kaerlas J., Padmanabhan J., Yadav J. S., Fauquet C.M., Dixon A., Fondong V. 2006. Production of the first transgenic cassava in Africa via direct shoot organogenesis from friable embryogenic calli and germination of maturing somatic embryos. African Journal of Biotechnology. 5 (19): 13. [ Links ]
11 Ihemere U., Arias-Garzon D., Lawrence S., Sayre R. T. 2006. Genetic modification of cassava for enhanced starch production. Plant Biotechnology Journal. 4 453-465. [ Links ]
12 Jørgensen K., Bak S., Busk P., Sørensen C., Olsen C., Puonti-Kaerlas J., Møller B. 2005. Cassava Plants with a Depleted Cyanogenic Glucoside Content in Leaves and Tubers. Distribution of Cyanogenic Glucosides, Their Site of Synthesis and Transport, and Blockage of the Biosynthesis by RNA Interference Technology. Plant Physiology. 139 363-374. [ Links ]
13 Li H. Q., Sautter C., Potrykus I., Puonti-Kaerlas J. 1996. Genetic transformation of cassava (Manihot esculenta Crantz). Nat Biotechnol. 14 (6): 736-40. [ Links ]
14 López D. 2000. Inducción de callo embriogénico friable, CEF, y regeneración de plantas de la variedad de yuca Manihot esculenta, Crantz, MCol 2215. Facultad de Ciencias Agropecuarias. Bsc 70. [ Links ]
15 Mathews H., Schopke C., Carcamo R., Chavarriaga P., Fauquet C., Beachy R. 1993. Improvement of somatic embryogenesis and plant recovery in cassava. Plant Cell Reports. 12 328-333. [ Links ]
16 Medero V. R., Borroto C., Rodríguez S., Gómez R., López J., García M., Ventura J., Espinosa J., Cabrera M., Martínez M., Torres M., Torres Y., Alvarez M., García J. 2000. Embriogénesis somática a partir de meristemos axilares en yuca. Biotecnología Vegetal. 1 21-26. [ Links ]
17 Medina R. D., Faloci M. M., Solís-Neffa V y Mroginski, V. 2003. Embriogénesis Somática y Regeneración de Plantas de Mandioca (Manihot esculenta Crantz) de Cultivares de Interés Para Argentina. Revista de Investigaciones Agropecuarias. 32 (3): 143-160. [ Links ]
18 Raemakers C. J. J. M., Schavemaker C. M., Jacobsen E., Visser R. G. 1993. Improvements of cyclic somatic embryogenesis of cassava (Manihot esculenta Crantz). Plant Cell Reports. 12 226-229. [ Links ]
19 Raemakers K., Schreuder M., Pereira I., Munyikwa T., Jacobsen E., Visser R. 2001. Progress made in FEC transformation of cassava. Euphytica. 120 15-24. [ Links ]
20 Raemakers K., Schreuder M., Suurs., Furrer-Verhorst L., Vincken J. P., Vetten N. D., Jacobsen E., Visser R. 2005. Improved cassava starch by antisense inhibition of granule-bound starch synthase I. Molecular Breeding. 16 163-172. [ Links ]
21 Restrepo S., Duque M. C., Verdier V. 2000. Characterization of pathotypes among isolates of Xanthomonas axonopodis pv. manihotis in Colombia. Plant Pathology. 49 (6): 680-687. [ Links ]
22 Roca W. M. 1984. Cassava. Macmillan Publishing; London, GB : Collier Macmillan Publishers. New York, NY, USA. . 269-301. [ Links ]
23 Sarria R., E. Torres, F. Angel, P. Chavarriaga y W. M. Roca. 2000. Transgenic plants of cassava (Manihot esculenta) with resistance to Basta obtained by Agrobacterium-mediated transformation. Plant Cell Reports. 19 339-344. [ Links ]
24 Schopke C., Taylor N., Carcamo R., Konan N. K., Marmey P., Henshaw G. G., Beachy R., Fauquet C. 1996. Regeneration of transgenic cassava plants (Manihot esculenta Crantz) from microbombarded embryogenic suspension cultures. Nat Biotechnol. 14 (6): 731-5. [ Links ]
25 Siritunga, D., Arias-Garzon D., White W., Sayre R. 2004. Over-expression of hydroxynitrile lyase in transgenic cassava roots accelerates cyanogenesis and food detoxification. Plant Biotechnology Journal. 2 37-43. [ Links ]
26 Siritunga D., Sayre R. T. 2003. Generation of cyanogen-free transgenic cassava. Planta. 217 367-373. [ Links ]
27 Sofiari E., Raemakers C., Danso K., Van Lammeren A., Jacobsen E., Visser R. 1997. Comparison of NAA and 2,4D induced somatic embryogenesis in cassava. Plant Cell Tiss Org Cult. 50 45-56. [ Links ]
28 Taylor N. J., Edwards M., Kiernan R. J., Davey C. D., Blakesley D., Henshaw D. D. 1996. Development of friable embryogenic callus and embryogenic suspension culture systems in cassava (Manihot esculenta Crantz). Nat Biotechnol. 14 (6): 726-30. [ Links ]
29 Taylor N. J., Masona M. V., Carcamo R., Ho T., Schöpke C., Fauquet C. M. 2001. Production of embryogenic tissues and regeneration of transgenic plants in cassava (Manihot esculenta Crantz). Euphytica. 120 25-34. [ Links ]
