










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Biotecnología
versión impresa ISSN 0123-3475
Rev. colomb. biotecnol vol.14 no.2 Bogotá jul./dic. 2012
ARTÍCULO CORTO
Influencia del tipo e intensidad de luz en la formación y multiplicación de embriones somáticos de soya
Influence of light type and intensity on induction and proliferation of somatic embryos in soybean
Jorge Liusvert Pérez Pérez1,2 , Teresa del Socorro Blanco Tirado3 , Lourdes García Rodríguez1 , Novisel Veitía Rodríguez1 , Idalmis Bermúdez Caraballoso 1 , Raúl Collado1 .
1Instituto de Biotecnología de las Plantas. Universidad Central "Marta Abreu" de Las Villas, Carretera a Camajuaní km 5.5, Santa Clara, Villa Clara, Cuba, CP 54830
2Centro de Estudios de Biotecnología Vegetal. Universidad de Granma, Carretera vía Manzanillo km 17.5, Peralejo, Bayamo, Granma, Cuba, CP 85100. *jperez@udg.co.cu
3Escuela de Ingeniería Agroindustrial. Instituto Universitario de la Paz, Barrancabermeja, Santander, Colombia.
Recibido: septiembre 24 de 2012 Aprobado: noviembre 28 de 2012
Resumen
La embriogénesis somática representa una alternativa para la regeneración de plantas, sobre la cual influyen diversos factores durante el cultivo in vitro. Entre estos, las condiciones de iluminación durante la formación de embriones somáticos es determinante. Sin embargo, no existen trabajos que refieran el efecto de la luz solar en la formación de embriones somáticos de soya. Este trabajo tuvo como objetivo evaluar el efecto del tipo e intensidad de luz en la formación y multiplicación de embriones somáticos de soya cultivar INCASoy-27. Para ello, cotiledones inmaduros de 3,0-4,0 mm de longitud fueron colocados en medio de cultivo con 40 mg.l-1 de ácido 2,4-diclorofenoxiacético, sacarosa 3% y pH 7,0 en cámaras de crecimiento con luz artificial e intensidades luminosas de 5-10 µmol.m-2.s-1 y 68-73 µmol.m-2.s-1, y en cámaras de crecimiento con luz solar con 50-65µmol.m-2.s-1. El mayor número de cotiledones embriogénicos (95,31) se obtuvo en condiciones de luz artificial con alta intensidad luminosa. La mayor formación de embriones somáticos se obtuvo en luz artificial (9,65) y luz solar (8,45) respectivamente con alta intensidad luminosa. No se encontraron diferencias significativas en la multiplicación de los embriones somáticos, al emplear ambas fuente de luz con alta intensidad luminosa. Esta es la primera referencia sobre formación y multiplicación de embriones somáticos de soya con empleo de luz solar.
Palabras clave: Cultivo de tejido, embriogénesis somática, fotoperíodo, Glycine max.
Abstract
The somatic embryogenesis represents an alternative for the regeneration of plants, in which influence diverse factors during the cultivation. Among these factors, the conditions of illumination during the induction of somatic embryos are determinant. However, works not exist that refer the effect of the sunlight on the formation of embryos in soybean. The goal of this work was to evaluate the effect of light type and intensity on the induction and proliferation of somatic embryos in soybean cultivar INCASoy-27. Immature cotyledons for 3,0-4,0 mm in length was placed on culture medium with 40 mg.l-1 2,4-diclorophenoxiacetic acid, sucrose 3%, and pH 7,0 under growth chamber with artificial light and range luminous intensity between 5-10 µmol.m-2.s-1 and 68-73 µmol.m-2.s-1 and growth chamber with sunlight from 50-65 µmol.m-2.s-1 were evaluated. The higher number of embryogenic cotyledons (95,31) was obtained into growth chamber with artificial light and intensity high. The results showed the major formation of embryos in artificial light (9,65) and sunlight (8,45) with luminous intensity high. They were not significant differences between two light types during somatic embryos multiplication. This is the first report on induction of somatic embryos in soybean with employment of sunlight as illumination source.
Key words: Tissue culture, somatic embryogenesis, photoperiod, Glycine max.
Introducción
La soya (Glycine max L. Merrill) constituye una fuente importante de aceite y proteína vegetal, empleada tanto para consumo humano como animal. Además, contiene metabolitos secundarios beneficiosos como, isoflavonas, tocoferoles y saponinas (Sakthivelu et al., 2008).
Debido a su importancia económica, se han realizado esfuerzos para establecer programas de mejoramiento genético con empleo de las técnicas de Ingeniería Genética. Estos procedimientos de transformación genética, requieren de un sistema de regeneración de plantas, un sistema de transferencia de genes y un mecanismo para la selección de los tejidos transformados (Yang et al., 2010).
Los sistemas de regeneración de plantas en soya, han estado ligados a dos rutas morfogenéticas fundamentales, organogénesis en nudos cotiledonales y embriogénesis somática en cotiledones cigóticos inmaduros. Sin embargo, la embriogénesis somática a partir de cotiledones inmaduros es genotipo dependiente y solo ha sido posible en un número reducido de cultivares que tienen capacidad para la respuesta embriogénica en condiciones de cultivo de in vitro (Hiraga et al., 2007; Regina et al., 2011).
La embriogénesis somática es un fenómeno complejo, influenciado por diversos factores donde los reguladores del crecimiento, la composición nutricional del medio de cultivo, el genotipo, las condiciones de luz y sus interacciones, influyen en la respuesta embriogénica y pueden ser manipulados para mejorar la eficiencia de este proceso morfogenético (Song et al., 2010).
Las condiciones de iluminación son particularmente importantes y existen evidencias del efecto de la calidad de la luz, en la formación y multiplicación de embriones somáticos, lo cual varía en dependencia de la especie y el estado de desarrollo del tejido (Rodríguez-Sahagún et al., 2011). Estos trabajos se refieren al efecto de la intensidad luminosa en condiciones de luz artificial (Bonacin et al., 2000; Rodríguez et al., 2004; Jin et al., 2004), pero ninguno evalúa la influencia de la luz solar en la formación de embriones somáticos de soya.
Una característica de la luz artificial y la luz solar, es que difieren en la calidad de su espectro luminoso, intensidad y duración (Hogewoning et al., 2010). Por ello, el objetivo de este trabajo fue evaluar el efecto del tipo e intensidad de luz en la formación y multiplicación de embriones somáticos de soya cultivar INCASoy-27.
Materiales y métodos
Material vegetal
Se empleó el cultivar cubano de soya INCASoy-27, obtenido en el Instituto Nacional de Ciencias Agrícolas de Cuba (INCA) por la vía de la hibridación natural del genotipo brasileño BR-32 en condiciones de campo (Ponce et al., 2003).
Para obtener plantas donantes se establecieron semillas maduras en bolsas de polietileno (26x36cm) que contenían como sustrato materia orgánica y zeolita (3:1). Las plantas fueron cultivadas en casa de cultivo del Instituto de Biotecnología de las Plantas, Villa Clara, Cuba. Alrededor de los 20 días posteriores a la ocurrencia de la antesis de la flor, fueron cosechadas las vainas que contenían embriones cigóticos inmaduros de 3,0-4,0 mm de longitud.
Desinfección
La desinfección se realizó en cabina de flujo laminar horizontal (modelo IKEM®). Las vainas fueron lavadas con agua corriente; luego sumergidas en etanol 70% (v/v) durante un minuto, seguido de tres enjuagues con agua destilada estéril. Se realizó una inmersión en hipoclorito de sodio 2% (v/v) con seis gotas de Tween-20 durante diez minutos y se lavaron cuatro veces con agua destilada estéril. Se tomaron los embriones cigóticos, se eliminó el eje embrionario y ambos cotiledones colocados con el lado abaxial sobre el medio de cultivo.
Condiciones de cultivo
Se emplearon frascos de vidrio transparentes de 250 ml de capacidad. Los frascos fueron colocados en cámaras de crecimiento con luz artificial, proporcionada por lámparas fluorescente de luz blanca (Phillips, 40 W), con fotoperíodo de 16/8 horas (luz/oscuridad) a 25±2°C; y cámaras de crecimiento con luz solar, con aproximadamente 12 horas de duración del período luminoso a 27±2°C.
La intensidad luminosa dentro de las cámaras de crecimiento se determinó mediante un luxómetro EXTECH 401025. En cámaras de crecimiento con luz artificial, los rangos de intensidad luminosa estuvieron influenciados por la distancia entre los frascos de vidrio y la fuente de luz. En cámaras de crecimiento con luz solar, fluctuó en dependencia de las condiciones ambientales, por ello se realizaron tres evaluaciones diarias y dos veces por semana durante el desarrollo experimental.
Formación de embriones somáticos
Para estudiar el efecto de las condiciones de iluminación en la formación de los embriones somáticos, se colocaron cuatro cotiledones inmaduros por frasco que contenían 30 ml de medio de cultivo compuesto por las sales de Murashige y Skoog (1962) (MS), vitaminas B5 (Gamborg et al., 1968), sacarosa 3%, ácido 2,4-diclorofenoxiacético (2,4-D) 40 mg.l-1, Gelrite® 0,3% (SIGMA), pH 7,0 (Samoylov et al., 1998).
Se establecieron 16 repeticiones (frascos) por rango de intensidad luminosa. En cámaras de crecimiento con luz solar con una densidad del flujo de fotones fotosintético (FFF) de 50-65 µmol.m-2.s-1 y, cámaras de crecimiento con luz artificial de alta (68-73 µmol.m-2.s-1) y baja (5-10 µmol.m-2.s-1) intensidad luminosa respectivamente.
A los 7, 14, 21 y 28 días de cultivo se cuantificó el número de cotiledones con embriones somáticos (%), referidos como cotiledones embriogénicos; y número de embriones somáticos en cotiledones con respuesta embriogénica. También, a los 21 y 28 días de cultivo se realizó una caracterización de los embriones somáticos basado en su morfología, consistencia y estado de desarrollo, con ayuda de un microscopio estereoscopio modelo WILD M8.
Para el análisis histológico de la ontogénesis de los embriones somáticos, se realizaron cortes histológicos en los cotiledones inmaduros. Estos fueron colocados durante 24 horas en una solución de fijación que contenía formaldehido 37% (v/v), ácido acético glacial 100% (v/v) y alcohol etílico 70% (v/v), en proporción 5:5:90. La deshidratación progresiva de las muestras se realizó en soluciones de alcohol 70% hasta llegar al alcohol etílico absoluto y la inclusión se realizó en bloques de Parafina (Kraus y Arduin, 1997).
Se realizaron cortes de 10 µm de espesor con un micrótomo de rotación (ZEISS), fijados en portaobjetos y teñidos con safranina (0,5%). Las muestras fueron observadas con ayuda de un microscopio óptico (AXIOSKOP) y las imágenes captadas con una cámara digital (OLYMPUS DP70) acoplada al ocular del microscopio.
Multiplicación de embriones somáticos
Posteriormente los embriones somáticos en etapa globular fueron transferidos a frascos de vidrio, que contenían 30 ml de medio de cultivo de multiplicación compuesto por las sales MS, vitaminas B5, sacarosa 3%, 2,4-D 20 mg.l-1, Gelrite® 0,3% y pH 5,8 (Wright et al., 1991).
Estos fueron colocados en dos cámaras de crecimiento con luz solar y luz artificial con alta intensidad luminosa. En cada tipo de luz se colocaron diez frascos de vidrio con cinco grupos de embriones somáticos, los cuales fueron cuantificados antes de su establecimiento.
Después de 28 días de cultivo se determinó la capacidad de multiplicación de los embriones somáticos en ambas cámaras de crecimiento, mediante la diferencia entre el número de embriones somáticos obtenidos al finalizar el ciclo de cultivo y el número de embriones somáticos colocados al inicio de la multiplicación.
Análisis estadístico
Para comparar las medias relacionadas con el número de cotiledones con respuesta embriogénica se aplicó la prueba de Kruskal-Wallis/Mann-Whitney. Las medias del número de embriones somáticos obtenidos en las tres intensidades de iluminación en un mismo periodo de tiempo, se analizó mediante la prueba de Dunnett´s C. La comparación de la tasa de multiplicación de los embriones somáticos en ambas cámaras de crecimiento se realizó mediante la prueba de hipótesis t-student.
En todos los casos se empleó un diseño factorial en bloques al azar. Para el procesamiento de los datos se empleó el programa estadístico SPSS (Statistic Package for Social Science) versión PASW® Statistics 18 y un intervalo de confianza del 95%.
Resultados
Se determinó que las condiciones de iluminación evaluadas influyeron en la formación de embriones somáticos a partir de cotiledones inmaduros de soya cultivar INCASoy-27.
Se observó que el número de cotiledones inmaduros que dieron origen a embriones somáticos tuvo un incremento exponencial con respecto al tiempo de cultivo, con la mayor respuesta embriogénica en alta intensidad luminosa independientemente de la fuente de luz empleada. De esta forma, del total de cotiledones establecidos en condiciones de luz solar solo el 67.19% tenían formación de embriones somáticos a los 14 días, hasta llegar al 71.88% (46 cotiledones embriogénicos) a los 28 días de cultivo.
Sin embargo, tomando como valor de referencia el total de cotiledones embriogénicos obtenidos a los 28 días de cultivo en cada intensidad luminosa (Figura 1a), se puedo determinar la frecuencia de aparición de los cotiledones embriogénicos y su distribución en el tiempo como se muestra en la figura 1b.
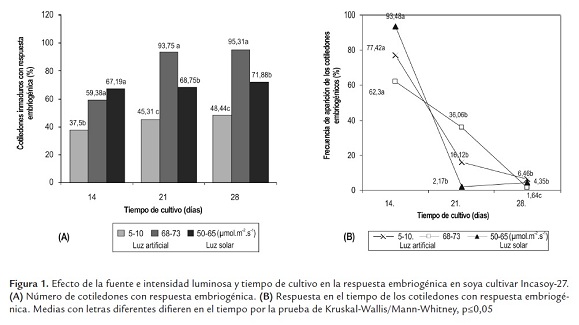
En esta figura se aprecia que en luz solar el 93,48% (43 cotiledones) y en luz artificial con baja intensidad luminosa el 77,42% de los cotiledones embriogénicos respectivamente, dieron origen a embriones somáticos durante los primeros 14 días de cultivo.
Esto demuestra que en las primeras dos semanas de cultivo, se obtuvo el mayor número de cotiledones con embriones somáticos y decreció con el tiempo sin diferencias estadísticas entre 21 y 28 días de cultivo. Por el contrario, en condiciones de luz artificial con alta intensidad luminosa, se obtuvo un incremento de 36,06% cotiledones embriogénicos con respecto a los obtenidos a los14 días de cultivo para un total de 93,75% cotiledones embriogénicos durante los primeros 21 días de cultivo.
Además, se evidenció mayor formación de embriones somáticos por cotiledones embriogénicos, cuando fueron cultivados en presencia de alta intensidad luminosa, independientemente del tipo de luz utilizada en un mismo período de tiempo (Figura 2).
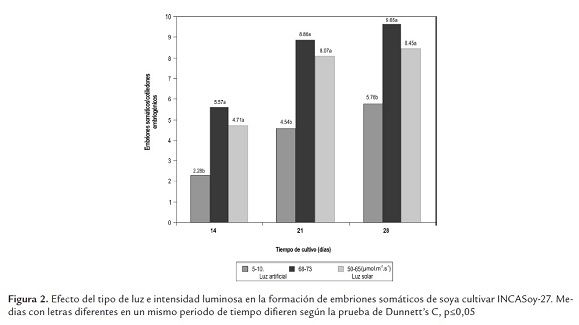
Al analizar la morfología de los embriones somáticos después de 21 días de cultivo, se encontró predominio de embriones somáticos en etapa globular y consistencia compacta (Figura 3a). Sin embargo, una semana después estos tenían un desarrollo asincrónico con diferentes tamaños y grados de desarrollo, siendo visible las etapas globular, corazón, torpedo y cotiledonal, típicas de esta ruta morfogenética y similares a la del embrión cigótico (Figura 3b).
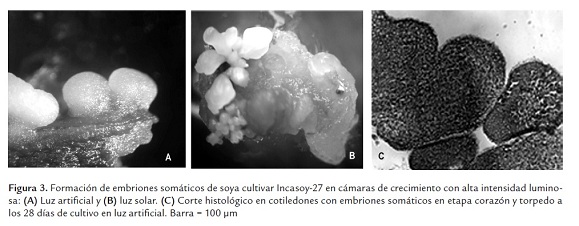
En la porción apical de los embriones somáticos en etapa globular, fue visible una invaginación, indicio del comienzo de su transición a la etapa de corazón. Los cortes histológicos corroboraron la presencia de embriones somáticos en diferentes etapas de desarrollo (Figura 3c).
Por otro lado, en cámaras de crecimiento con luz artificial y luz solar respectivamente, fue visible la formación estratificada de embriones somáticos, tanto en la superficie apical como basal de los embriones somáticos primarios, dando origen a masas embriogénicas (Figura 4).
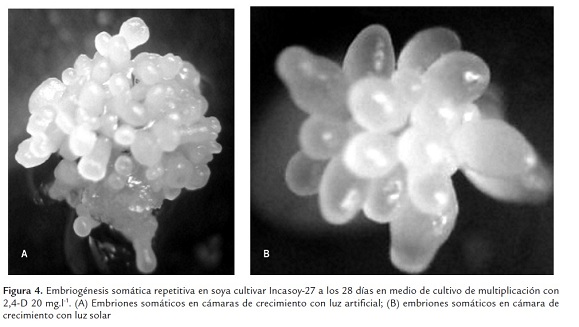
Esta continua multiplicación de los embriones somáticos de soya, permitió un incremento en el número de embriones somáticos al finalizar la fase de multiplicación con respecto al número de embriones somáticos inicialmente establecidos en cada una de las fuentes de luz. Sin embrago, no se observaron diferencias significativas en la tasa de multiplicación de los embriones somáticos en ambos tipos de luz.
Estos resultados indican que con alta intensidad luminosa es posible incrementar la formación de embriones somáticos, sin encontrar diferencias significativas entre los 21 y 28 días de cultivo, lo que permite disminuir el tiempo de exposición de los tejidos a las altas concentraciones de 2,4-D en el cultivar cubano de soya INCASoy-27.
Discusión
El empleo de alta intensidad luminosa tanto en condiciones de luz solar como artificial, favoreció la formación de embriones somáticos de soya cultivar Incasoy-27, quizás debido a que al tejido llegó una mayor cantidad de irradiación luminosa. Esta influencia de la luz en la formación de embriones somáticos, puede ser una de las causas por las que no se ha establecido la embriogénesis somática en explantes de soya cultivados en condiciones de oscuridad.
Estas variaciones en la capacidad embriogénica de los cotiledones en las diferentes fuentes de luz evaluadas, pudieron estar influenciadas por el espectro de luz que llegó al tejido de los cotiledones. Es conocido que en cámaras de crecimiento con luz artificial el espectro luminoso es continuo. Sin embargo, bajo condiciones de luz solar los tejidos están expuestos a las diferencias espectrales que varían en dependencia de las condiciones climáticas, duración del período luminoso y el ambiente de cultivo in vitro (Hogewoning et al., 2010).
A su vez, la variación observada en este trabajo en cuanto al número de cotiledones con embriones somáticos, refleja que durante los primeros 14 días de cultivo ocurre una intensa actividad mitótica en los tejidos cotiledonales, dando origen a las estructuras embriogénicas principalmente en presencia de alta intensidad luminosa.
La literatura científica refiere que las lámparas fluorescentes usadas en el cultivo in vitro aportan luz roja y luz azul, lo que unido a la intensidad luminosa activa los fotoreceptores del tejido e inducen dicho proceso morfogenético. El color rojo del espectro luminoso inducen la formación de embriones somáticos, y el color azul los inhibe aunque favorece su desarrollo (Hoshino y Cuello, 2006).
Los resultados alcanzados en este trabajo difieren con los referidos por Bonacin et al. (2000) y Mauro et al. (2001), quienes no encontraron diferencias en la respuesta embriogénica en cinco genotipos brasileños cultivados en rangos de intensidad luminosa entre 8-12 μmol.m-2.s-1 y 27-33 μmol.m-2.s-1, aludido a las bajas intensidades luminosas evaluados. Mientras que Jin et al. (2004), al evaluar intensidades de iluminación de 10, 20 y 40 μmol.m-2.s-1, obtuvieron la mejor respuesta de formación de embriones somáticos en cotiledones inmaduros expuestos a 20 μmol.m-2.s-1, sin diferencias entre 20 y 40 μmol.m-2.s-1 durante el desarrollo embrionario.
De igual manera, indican una relación directa entre el número de estructuras embriogénicas obtenidas en las fases de formación y multiplicación de embriones somático, y difiere de los resultados obtenidos por Bailey et al. (1993) y Simmonds y Donaldson (2000), quienes observaron que cultivares con alta capacidad de formación de embriones somáticos desarrollaron un bajo potencial de proliferación.
Una de las causas que pudieron haber influido en los resultados alcanzados en este trabajo es el espectro de luz. Al respecto, Hogewoning et al. (2010) plantearon que en cámaras de crecimiento con luz artificial esta es continua, mientras que en condiciones de luz solar, los tejidos son expuestos a las diferencias espectrales que varían en dependencia de las condiciones de tiempo, duración del día y el ambiente de cultivo. En este sentido, Franklin y Whitelam (2007) describen que en días nublados el espectro de luz solar contiene más azul y menos rojo lejano entre 700-750 nm que en días normales, y una baja proporción del rojo a rojo lejano.
Algunos trabajos refieren que la luz influye en la inducción y morfología de embriones somáticos diferenciados. Una posible explicación de la diferenciación de los embriones somáticos, es la correlación entre la proporción de luz del rojo lejano y el rojo absorbido en los fitocromos, lo cual puede producir efectos inhibitorios en dependencia de la cantidad de fitocromos activados (Torné et al., 2001).
Por su parte, D’Onofrio et al. (1998), informaron sobre los efectos de la luz en la diferenciación de embriones somáticos en Sidonia sp. Al correlacionar la proporción de embriones somáticos diferenciados con el fotoequilibrio, encontraron un incremento exponencial desde 0%-30% de embriones somáticos diferenciados al incrementar el fotoequilibrio de cero a uno, lo que sugiere una actividad de los fitocromos para inducir la embriogénesis somática.
Se ha encontrado que el espectro de la luz roja y alta intensidad luminosa favorece la formación de embriones somáticos en células embriogénicas de Zanahoria (Daucus carota). Por otro lado, una duración del fotoperíodo de 16 horas luz, incrementa el número y tamaño de los embriones somáticos en la especie Miraguano (Araujia sericifera); mientras que en Agave azul (Agave tequilana), 16 horas luz durante la fase de formación de embriones somáticos, estimula la sensibilidad de los callos embriogénicos (Torné et al., 2001; Rodríguez-Sahagún et al., 2011). Algunos trabajos refieren que la luz influye en la inducción y morfología de embriones somáticos, pero poco es conocido acerca del efecto de la calidad y cantidad de luz sobre parámetros fisiológicos y bioquímicos del explante, los cuales varían entre especie y genotipo. Se plantea que el 2,4-D y la luz, son capaces de inducir una explosión oxidativa en el tejido para producir especies de oxígeno reactivo. Estos cambios en el ambiente celular del cultivo in vitro, inducido por factores de estrés del medio de cultivo o el ambiente físico, son responsables de la reprogramación de la expresión génica. En este contexto, se promueve la desdiferenciación celular e induce la formación de embriones somáticos (Zavattieri et al., 2010).
Sustancias celulares como el peróxido de hidrógeno y óxido nítrico, inducen la expresión del gen oxidasa alternativo (del inglés, alternative oxidase: AOX) y la embriogénesis somática. Este gen reduce la producción de especies de oxígeno reactivo en la mitocondria de las células vegetales. Así, puede ser considerada una correlación entre la expresión de dicho gen, las respuestas de estrés y reprogramación celular vía embriogénica (Arnholdt-Schmitt et al., 2006).
Esto concuerda con Widholm et al. (2010), quienes plantearon que si la concentración de 2,4-D disminuye por debajo del umbral requerido para formar embriones somáticos, estos continúan su desarrollo utilizando los nutrientes del medio de cultivo. Además, el 2,4-D una vez absorbido por el tejido vegetal, puede ser degradado o conjugado con otras auxinas pues está expuesto a factores físicos y sujeto a la conversión enzimática (Machakova et al., 2008).
Conclusiones
Los resultados obtenidos en este trabajo, permitieron demostrar el efecto del tipo e intensidad de luz en la formación y multiplicación de embriones somáticos de soya cultivar INCASoy-27. Igualmente, el empleo de la luz solar representa una alternativa para el establecimiento de una metodología de regeneración de plantas vía embriogénesis somática de soya en Cuba.
Referencias bibliográficas
1 Arnholdt-Schmitt, B.; Costa, J.H.; Fernándes, D. 2006. AOX - a functional marker for efficient cell reprogramming under stress?. Trends in Plant Science. 11 (6): 281-287. [ Links ]
2 Bailey, M.A; Boerma, H.R.; Parrott, W.A. 1993. Genotype-specific optimization of plant regeneration from somatic embryos of soybean. Plant Science. 93:117-120. [ Links ]
3 Bonacin, G.A.; Di, A.O.; Carlos, R.; Perecin, D. 2000. Induction of somatic embryogenesis in soybean: physicochemical factors influencing the development of somatic embryos. Genetics and Molecular Biology. 23 (4): 865-868. [ Links ]
4 D"Onofrio, C.; Morini, S.; Bellocchi, G. 1998. Effect of light quality on somatic embryogenesis of quince leaves. Plant Cell, Tissue and Organ Culture. 53:91-98. [ Links ]
5 Franklin, K.; Whitelam, G. 2007. Red: far-red ratio perception and shade avoidance. En: Whitelam G, Halliday K, (eds), Light and plant development. (Ed.) Oxford: Blackwell Publishing, pp: 211-234. [ Links ]
6 Gamborg, O.L.; Miller, R.A.; Ojima, K. 1968. Plant cell cultures. I. Nutrient requirements of suspension cultures of soybean root cells. Experimental Cell Research. 50 (1):151-158. [ Links ]
7 Hiraga, S.; Minakawa, H.; Takahashi, K.; Takahashi, R.; Hajika, M.; Harada, K.; Ohtsubo, N. 2007. Evaluation of somatic embryogenesis from immature cotyledons of Japanese soybean cultivars. Plant Biotechnology. 24 (4): 435 440. [ Links ]
8 Hogewoning, S.W.; Douwstra, P.; Trouwborst, G.; van Leperen, W.; Harbinson, J. 2010. An artificial solar spectrum substantially alters plant development compared with usual climate room irradiance spectra. Journal of Experimental Botany. 61 (5): 1267-1276. doi:10.1093/jxb/erq005. [ Links ]
9 Hoshino, T.; Cuello, J.L. 2006. Environmental design considerations for somatic embryogenesis. Plant Cell Monographs (2). En: A. Mujib y J. Samaj (eds). Somatic Embryogenesis. (Ed) Springer-Verlag, Berlin Heidelberg, pp: 25-34. [ Links ]
10 Jin, Y.; ll, T.; Soon, H.; Ki, H.; Uk, S.; Joong, S. 2004. Factors affecting somatic embryogenesis from immature cotyledon of soybean. Journal of Plant Biotechnology. 6 (1): 45-50. [ Links ]
11 Kraus, J.E.; Arduin, M. 1997. Manual Básico de Métodos em Morfología vegetal. Editora da Universidade Federal Rural do Rio de Janeiro, Seropédica, Rio de Janeiro, Brasil, 25p. [ Links ]
12 Machakova, I.; Zazimalova, E.; George, E.F. 2008. Plant Growth Regulators I: Introduction; Auxins, their Analogues and Inhibitors. En: E.F. George, M.A. Hall, G.J. De Klerk (eds), Plant Propagation by Tissue Culture, 3rd Edition, Volume 1. The Background. (Ed.) Springer, pp: 175-204. [ Links ]
13 Mauro, A.O.; Oliveira, R.C.; Bonacin, G.A.; Oliveira, J.A.; Mauro, S.M.Z.; Collins, G.B. 2001. A quantitative nature of somatic embryogenesis in soybean. In vitro Cellular & Developmental Biology-Plant. 37: 773 777. [ Links ]
14 Murashige, T.; Skoog, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiology Plant. 15 (3): 473-497. [ Links ]
15 Ponce, M.; de la Fé, C.; Ortiz, R.; Moya, C. 2003. Informe de nuevas variedades. INCASOY-24 e INCASOY-27: Nuevas variedades de soya para las condiciones climáticas de Cuba. Cultivos Tropicales. 24 (3): 49. [ Links ]
16 Regina, L.; Lucca, A., Martinho, B.G.; Negra, E.S.; Kelly, P., Schuster, I. 2011. Evaluation of soybean cultivars on the embryogenic and organogenic potential. Acta Scientiarum Agronomy. 33 (1): 67-74. doi: 10.4025/actasciagron.v33i1.12344. [ Links ]
17 Rodríguez, L.R.; Forte B.C.; Oliveira, J.M:S.; Mariath, J.E.A.; Bodanese-Zanettini, M.H. 2004. Effects of light conditions and 2,4-D concentration in soybean anther culture. Plant Growth Regulation. 44: 125-131. [ Links ]
18 Rodríguez-Sahagún, A.; Acevedo-Hernández, G.; Rodríguez-Domínguez, J.M.; Rodríguez-Garay, B.; Cervantes-Martínez, J.; Castellanos-Hernández, O.A. 2011. Effect of light quality and culture medium on somatic embryogenesis of Agave tequilana Weber var. Azul. Plant Cell, Tissue and Organ Culture. 104 (2): 271-275. doi: 10.1007/s11240-010-9815-4. [ Links ]
19 Sakthivelu, G.; Akitha, M.K.; Giridhar, P.; Rajasekaran, T.; Ravishankar, G.A.; Nikolova, M.T.; Angelov, G.B.; Todorova, R.M.; Kustorkova, G. 2008. Isoflavone composition, phenol content and antioxidant activity of soybean seeds from India and Bulgaria. Journal of Agricultural and Food Chemistry. 56 (6): 2090-2095. [ Links ]
20 Samoylov, V.M.; Tucker, D.M.; Parrott, W.A. 1998. Soybean [Glycine max (L.) Merrill] embryogenic cultures: the role of sucrose and total nitrogen content on proliferation. In vitro Cellular & Developmental Biology-Plant. 34: 8-13. [ Links ]
21 Simmonds, D.; Donaldson, P. 2000. Genotype screening for proliferative embryogenesis and biolistic transformation of short season soybean genotypes. Plant Cell Reports. 19(5): 485-490. [ Links ]
22 Song, X.; Han, Y.; Teng, W.; Sun, G.; Li, W. 2010. Identification of QTL underlying somatic embryogenesis capacity of immature embryos in soybean [Glycine max (L.) Merrill]. Plant Cell Reports. 29 (2): 125-131. doi: 10.1007/s00299-009-0804-1. [ Links ]
23 Torné, J.M.; Moysset, L.; Santos, M.; Simón, E. 2001. Effect of light quality on somatic embryogenesis in Arauija sericifera. Physiology Plant, 111: 405-411. [ Links ]
24 von Arnold, S. 2008. Somatic embryogenesis. En: Plant Propagation by Tissue Culture 3rd Edition. Volume 1. The Background. (eds) Edwin F. George, Michael A. Hall, Geert-Jan De Klerk, (Ed.) Springer, p. 335-354. [ Links ]
25 Widholm, J.M.; Finer, J.J.; Vodkin, L.O.; Trick, H.N.; LaFayette, P.; Li, J.; Parrott, W. 2010. Chapter 24 Soybean. En: F. Kempken y C. Jung (eds.), Genetic Modification of Plants, Biotechnology in Agriculture and Forestry 64, (Ed.) Springer-Verlag, Berlin Heidelberg, pp. 473 - 498. [ Links ]
26 Wright, M.S.; Launis, K.L.; Novitzky, R.; Duesing, J.H.; Harms, C.T. 1991. A simple method for the recovery of multiple fertile plants from individual somatic embryos of soybean [Glycine max (L.) Merrill]. In vitro Cellular & Developmental Biology-Plant. 27P:153-157. [ Links ]
27 Yang, J.L.; Seong, E.S.; Kim, M.J.; Ghimire, B.K.; Kang, W.H.; Yeon, C.; Hao, C. 2010. Direct somatic embryogenesis from pericycle cells of broccoli (Brassica oleracea L. var. Italica) root explants. Plant Cell, Tissue and Organ Culture. 100 (1): 49-58. doi: 10.1007/s11240-009-9615-x. [ Links ]
28 Zavattieri, M.A.; Frederico, A.M.; Lima, M.; Sabino, R.; Arnholdt-Schmitt, B. 2010. Induction of somatic embryogenesis as an example of stress-related plant reactions. Electronic Journal of Biotechnology. 13 (1) January 15. doi: 10.2225/vol13-issue1-fulltext-4. [ Links ]
