










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol vol.14 no.2 Bogotá jul./dez. 2012
ARTÍCULO DE REVISIÓN
Métodos y usos agrícolas de la ingeniería genética aplicada al cultivo del arroz
Methods and agricultural uses of genetic engineering applied to rice crop
Cristina DiazGranados D.1 , Alejandro Chaparro-Giraldo.2 .
1 Microbióloga Industrial, M.Sc., Estudiantes del doctorado en Biotecnología de la Universidad Nacional de Colombia, sede Bogotá, FEDEARROZ, Grupo de Ingeniería Genética de Plantas. E-mail: ecdiazg@unal.edu.co
2 I.A., MSc., PhD. Grupo de Ingeniería Genética de Plantas, Departamento de Biología- Instituto de Genética, Universidad Nacional de Colombia, sede Bogotá. E-mail: achaparrog@unal.edu.co
Recibido: abril 11 de 2012 Aprobado: noviembre 29 de 2012
Resumen
En biotecnología de arroz se han logrado avances en transformación genética, con importantes resultados en el mejoramiento genético de variedades elite de las subespecies japónica e índica. Con el propósito de revisar los métodos y los usos agrícolas de la ingeniería genética aplicada al cultivo del arroz, se usaron varias palabras claves en idioma inglés en algunas de las bases de datos de revistas científicas indexadas, disponibles en el Sistema Nacional de Bibliotecas de la Universidad Nacional de Colombia (SINAB), seleccionando documentos publicados entre 2000 y 2011. La base de esta revisión inicial, se complementó con artículos publicados en fechas anteriores, que se consideraron relevantes, debido a que implicaban cambios metodológicos importantes. Desde que se logró producir la primera planta transgénica de arroz a finales de los 80´s, varios protocolos para la transferencia de genes se han empleado con éxito logrando la modificación genética de más de 60 cultivares de arroz. Para ello se han empleado sistemas de transformación tanto directos como indirectos. Se han realizado modificaciones de rasgos importantes en el cultivo, tales como la resistencia a factores bióticos (insectos, hongos, bacterias, virus, nematodos), tolerancia a factores abióticos (salinidad, sequía, altas y bajas temperaturas, inmersión), y mejoramiento de características agronómicas (calidad nutricional, rendimiento, uso de nutrientes, tolerancia a herbicidas).
Palabras clave: Arroz, cultivos transgénicos, factores bióticos, factores abióticos.
Abstract
In rice biotechnology advances have been made in genetic transformation, with significant results in breeding elite varieties of japonica and indica subspecies. In order to review the methods and agricultural uses of genetic engineering applied to rice, calves were used several words in English in some of the databases of scientific journals available in the National Libraries National University of Colombia (SINAB), selecting papers published between 2000 and 2011. Based on this initial review, it addition to some articles published at earlier dates, which were considered relevant because they involved significant methodological changes. Since it was able to produce the first transgenic rice plant in the late 80's, several protocols for gene transfer have been used successfully achieving the genetic modification of more than 60 rice cultivars. For this transformation systems have been used both direct and indirect. There have been significant changes in the crop traits such as resistance to biotic (insects, fungi, bacteria, viruses, nematodes), tolerance to abiotic (salinity, drought, high and low temperatures, immersion), improved features agronomic (nutritional quality, yield, nutrient use, herbicide tolerance).
Key words: Rice, transgenic crops, biotic factor, abiotic factor.
Introducción
El arroz (Oryza sativa) es cultivado en 113 países y en todos los continentes, está profundamente integrado en el patrimonio cultural de muchas sociedades. Es considerado como uno de los cultivos de mayor importancia para la alimentación mundial, ya que es el alimento básico de más de la mitad de la población mundial y el 40% depende de éste para el 80% de su dieta. Los sistemas basados en el arroz apoyan enormes reservas de agrobiodiversidad, que sirven para salvaguardar el medio ambiente, aumentar los medios de subsistencia y enriquecer la alimentación de la población (FAO, 2004; SMIA, 2006).
Existen más de 2000 variedades de arroz cultivadas en el mundo. Las diferencias se refieren a la morfología de la planta y del grano, la calidad del grano, la resistencia al volcamiento, la precocidad, la ramificación, la resistencia y tolerancia a los factores bióticos (malezas, insectos y enfermedades) y abióticos (frío, sequía, acidez del suelo, carencias en elementos minerales primordiales, etc.) y la productividad física (FAO,2002; Nene, 2005)
La declaración del 2004 como año internacional del arroz por las Naciones Unidas refleja el papel primordial que el cultivo ha adquirido. Además el arroz ha sido seleccionado como especie modelo para la investigación dentro del grupo de los cereales debido a sus características genéticas únicas. Como resultado se han obtenido mapas genéticos, físicos y comparativos, la caracterización de varios genes y la secuencia completa de su genoma. Con estas herramientas se espera apoyar y optimizar conjuntamente con otras tecnologías los procesos de mejoramiento de la especie y de otras relacionadas (Shimamoto & Kyozuka, 2002; Bajaj & Mohanty, 2005).
El mejoramiento genético de plantas a través de la ingeniería genética ha resultado en la siembra de 160 millones de hectáreas de variedades vegetales genéticamente modificadas en el 2011, con características de resistencia a herbicidas e insectos principalmente, sembradas en 29 países por 16.7 millones de agricultores (James, 2011). La ingeniería genética juega un papel fundamental en el mejoramiento de las especies vegetales permitiendo la introducción de genes de interés, además de ampliar el pool genético existente.
Las modificaciones genéticas producidas en el arroz han tenido diversos fines. Muchas de ellas fueron realizadas solo como ensayos para posteriores manipulaciones. Entre las características introducidas al arroz transgénico encontramos principalmente tolerancia a diferentes condiciones ambientales de estrés, resistencia a patógenos o plagas y tolerancia a herbicidas (Roy & Wu, 2001; Agrawal et al, 2003). Entre los grandes avances logrados en la biotecnología del arroz esta la implementación del uso del sistema de Agrobacterium que hasta hace poco estaba limitado a dicotiledóneas (Bajaj & Mohanty, 2005) y las diversas modificaciones en el grano, como el arroz dorado con beta caroteno (Al-Babili & Beyyer, 2005; Li, 2006).
Existe una gran cantidad de información relacionada con la transformación genética de arroz. Organizar esta información es una tarea gigantesca, pues día a día se encuentran nuevos trabajos relacionados, no solo con propósitos académicos, sino también comerciales. Para la realización de este documento se consultaron varia bases de datos, como "Science Direct", "Medline", "Pubmed", "Annual reviews" y "Springerlink" disponibles en el portal del sistema nacional de bibliotecas (SINAB) de la Universidad Nacional de Colombia (http://www.sinab.unal.edu.co/). Se emplearon palabras claves en ingles como por ejemplo: "transgenic rice", "herbicide tolerance", "insect resistence", "drought tolerance", solas o en combinación, limitando la búsqueda a solo titulo, resumen y palabras claves, y a partir del año 2000. La cantidad de entradas era enorme, por ejemplo el mayor número de entradas se encontró en la base de datos "Springerlink" con 13.677. Por ello, se seleccionaron los documentos que se consideraban relevantes, bien sea como hitos en la historia de la tecnología del DNA recombinante aplicada en el arroz, o que describían cambios importantes en la aplicación de las técnicas y en las aproximaciones experimentales. Se seleccionaron algunos documentos anteriores al año 2000, sobre la base de las anteriores consideraciones. Algo importante de resaltar es que cuando se usaban las palabras "transgenic indica rice" el número de entradas se disminuía drásticamente, para la misma base de datos "Springerlink" el número disminuyó a 29. Esto confirma lo planteado por Delteil y colaboradores (2010), que la mayoría de la información y trabajos realizados en arroz se ha desarrollado en la subespecie japónica, esto puede deberse a que la respuesta al cultivo de tejidos y transformación genética es muy buena en esta subespecie, que además es la más cultivaba en el mundo. Mientras que la subespecie indica es considerada como recalcitrante y de difícil manejo, en cuanto a la ingeniería genética de plantas.
Análisis crítico
Generalidades del cultivo de arroz.
El arroz (Oryza sativa) es una planta monocotiledónea, que en los trópicos puede completan su ciclo de vida en un período de 110 a 210 días. Esta planta anual, posee tallos redondos y huecos compuestos de nudos y entrenudos muy ramificados, puede medir entre 0,6 y 1,8 metros de altura, con hojas de lámina plana unidas al tallo por la vaina. Los tallos terminan en una "inflorescencia" o panícula de 20 a 30 cm de largo. Cada panícula se compone entre 50 y 300 flores o "espiguillas", a partir de las cuales se formarán los granos. El fruto obtenido es un "cariópsis". Los granos de arroz pueden clasificarse según su longitud en: Extralargo (EL) 7,6 mm o más; Largo (L) 7,5 mm a ,6 mm; Medio (M) 6,5 mm a 5,6 mm; Corto (C) 5,5 mm o menos (CIAT, 2005; Degiovanni et al, 2010). Los compuestos que constituyen el arroz son esencialmente: almidón, proteínas, grasas, ligninas y cenizas, con indicios de numerosos metales y vitaminas. Con excepción del almidón, todos los demás compuestos se encuentran en las capas externas de la cariópside y en el germen (Xu et al, 2008; Degiovanni et al, 2010).
El arroz es una gramínea autógama, que crece con mayor facilidad en los climas tropicales. Originariamente era una planta cultivada en seco pero con las mutaciones se convirtió en semi-acuática, aunque puede crecer en medios bastante diversos, crece más rápidamente y con mayor vigor en un medio caliente y húmedo (Degiovanni et al, 2010).
Inicialmente se planteo que el género Oryza se originó hace cerca de 130 de años en el megacontinente Gondwana y tras su fragmentación las especies de la época se distribuyeron en distintos continentes. Actualmente esto se ha replanteado de acuerdo a la nueva evidencia que se tiene sobre el origen de las monocotiledóneas y los pastos, donde se plantea que los ancestros del arroz se separaron hace 50 millones de años, y que el origen del género Oryza es de unos 14 a 9 millos de años (Khush, 1997; Vaughan et al ,2003; Tang et al, 2010).
El complejo genético del arroz está constituido por veinticuatro especies del género Oryza, donde se incluyen perennes, anuales, diploides y tetraploides. Con diez genomas distintos, conocidos como: AA, BB, CC, EE, FF, GG, CCDD, BBCC, HHJJ y HHKK. De estas veintidós son silvestres y solo dos son cultivadas (Rana, 2004). De acuerdo al tipo de genoma y afinidad genética, las distintas especies de Oryza, se han agrupado en cuatro complejos (Vaughan et al,2003; Londo et al, 2006;Kumar et al, 2008), estos son: (1) Complejo O. sativa: Incluye las especies O. sativa, O longistaminata, O. barthii, O. glaberrima, O. rufipogon, O. meridionales, O. nivara y O. glumaepatula. Su genoma es del tipo AA. (2) Complejo O. officinalis: Comprende especies con genomas BB, BBCC, CC, CCDD y EE. Hacen parte de este Oryza officinalis, O. minuta, O. eichingeri O. punctata O. latifolia O. alta O. grandiglumis O. australiensis y O. rhizomatis. (3) Complejo O. granulata y O. brachyantha: Además de estas dos especiesse incluye a O. meyeriana. Algunos también consideran la especie O. neocaledonica. Las especies que corresponden a este grupo presentan genomas FF y GG. (4) Complejo O. ridleyi y O. schlechteri: Comprende estas dos especies más la especie O. longiglumis. El genoma de las especies de este grupo es de tipo HHJJ y HHKK.
La diversidad genómica y de especies del complejo Oryza se centra en las islas del sureste de Asia en el océano Pacífico, de las 22 especies silvestres 9 están en Indonesia y 7 de los 10 tipos de genomas se encuentran en las islas del Pacífico Asiático. Algunos grupos distintivos de especies también están asociados con el Sur de Asia, África y las Américas (Toriyama et al, 2005; Johns & Mao, 2007; Ikehashi, 2009).
Las dos especies cultivadas Oryza sativa y Oryza glaberrima son diploides con genoma AA (Rana, 2004; Purugganan, 2010). Según varios estudios se considera que los ancestros de O. sativa pueden ser O. nivara o O. rufipogon, especies que se encuentran en varias partes de la India y en áreas adyacentes; mientras que Oryza glaberrima probablemente se originó de O. longistaminata o O. barthii especies que se encuentran exclusivamente en África (Khush, 1997; Vaughan et al ,2003; Sang & Ge, 2007). El arroz africano cultivado pudo haberse originado en el oriente africano y domesticado en el delta del Río Níger, y su cultivo está prácticamente confinado a la región de origen (Rana, 2004; Ikehashi, 2009). Mientras que por la amplia dispersión de O. sativa, esta es cultivada en regiones que abarcan latitudes entre 5ºN y 36ºS; y por su capacidad de adaptación a diferentes condiciones agroclimáticas y por la selección humana, se ha obtenido numerosos cultivares. Se estima que existen alrededor de 120.000 variedades en el mundo (Rana, 2004; Londo et al, 2006)
Los centros de origen de O. sativa se ubican en el Sur y Este de Asía, desde allí se extendió hacia otros lugares, hasta cubrir prácticamente todos los continentes a excepción de la Antártica, y su aparición data en más de 9000 años. En el proceso de dispersión y adaptación a nuevos ambientes, surgieron formas diferentes clasificadas en dos grupos Indica y japónica, considerados por algunos como subespecies y por otros como tipos. Se considera que probablemente estos dos grupos se domesticaron independientemente (Vaughan et al ,2003; Johns & Mao, 2007; Ikehashi, 2009; Purugganan, 2010) .
Las formas indicas de O. sativa, probablemente se domesticaron en las partes bajas de los Himalayas y oriente de la India, dispersándose por las regiones tropicales y subtropicales de India, y posteriormente a otras áreas del mundo. Las formas japónicas se domesticaron en alguna parte del Sur de China, dispersándose hacia el norte conformando las formas de tipo templado, y otras se extendieron hacia el Sur de Asia y al oeste de África generando los tipos tropicales, también llamados arroces jabanicos (Vaughan et al ,2003; Toriyama et al,2005; Garris et al, 2005; Ikehashi, 2009; Purugganan, 2010)
El genoma de la subespecie Indica posee entre 45.000 y 55.000 genes, mientras que el genoma de la subespecie japónica contiene entre 43.000 y 50.000 genes (Yu et al, 2005). Los centros primarios de diversidad de O. sativa se relacionan directamente con los centros de origen, sin embargo se considera un centro adicional que es el sur de Asia y las islas cercanas, el cual esta asociado al entrecruzamiento de las formas indica y japónica (Vaughan et al ,2003; Toriyama et al, 2005; Purugganan, 2010). Con la llegada de la especie a otras áreas, hace 2000 años aproximadamente, se dio el surgimiento de centros secundarios de diversidad con formas distintivas, los cuales son: Océano Indico, oriente asiático y Europa. Actualmente se han considerado centros más recientes en África y Suramérica (Londo et al, 2006; Ikehashi, 2009; Purugganan, 2010)
Importancia del cultivo de arroz
El arroz (Oryza sativa L.), es cultivado en 113 países y en todos los continentes, está profundamente integrado en el patrimonio cultural de muchas sociedades. Muchos consideran el cultivo de arroz el más importante del mundo, principalmente si se toma en cuenta la extensión cultivada y la cantidad de gente que depende de su cosecha. Es el alimento básico de más de la mitad de la población mundial, ya que más del 40% depende de este para el 80% de su dieta y proporciona el 20% del consumo de calorías per cápita en todo el mundo. El arroz es el alimento básico en 17 países de Asia y el Pacífico, 8 países de África, 7 países de América Latina y el Caribe, y 1 país del Cercano Oriente. Si se consideran en conjunto todos los países en desarrollo, el arroz proporciona el 27% de la energía alimentaría y el 20% de la ingesta de proteína alimentaría (FAO, 2002; FAO, 2004; SMIA, 2006; IDEAS, 2007; Degiovanni et al, 2010).
Los sistemas basados en el arroz apoyan enormes reservas de agrobiodiversidad, que sirven para salvaguardar el medio ambiente, aumentar los medios de subsistencia y enriquecer la alimentación de la población. Es un cultivo rico en diversidad genética y sus dos especies cultivables, Oryza sativa L. (originaria de Asia) y O. glaberrima (originaria del África occidental), han producido miles de variedades diferentes con gran variedad de propiedades nutritiva (IDEAS, 2007; Degiovanni et al, 2010). Los sistemas de producción basados en el arroz y las operaciones conexas poscosecha emplean casi mil millones de personas en las zonas rurales de los países en desarrollo. Alrededor de las cuatro quintas partes del arroz del mundo es cultivada por pequeños agricultores en países en desarrollo de bajos ingresos (FAO, 2002; SMIA, 2006)
El arroz es producido fundamentalmente por los países para satisfacer su propio consumo. Los países asiáticos son los que más consumen, 90% del consumo mundial de arroz elaborado es realizado por ellos. China e India son los principales consumidores, el país no asiático con mayor consumo de arroz es Brasil (FAO, 2002; FAO, 2004; Benavides & Segura, 2005)
Cultivo del arroz
Existen más de 2.000 variedades de arroz cultivadas en el mundo. Las diferencias se refieren a la morfología de la planta y del grano, la calidad del grano, la resistencia al volcamiento, la precocidad, la ramificación, la resistencia y tolerancia a los factores bióticos (malezas, insectos y enfermedades) y abióticos (frío, sequía, acidez del suelo, carencias en elementos minerales primordiales, etc.) y la productividad física (UNO,2002; Nene, 2005; IDEAS, 2007)
Para obtener cosechas optimas, el arroz requiere una combinación de varios factores como: temperaturas suaves, para la subespecie japónica una temperatura mínima de 12°C y de 13°C para la subespecie indica; disponibilidad suficiente de agua y trabajo minucioso (Lozano, 2002; Diago, 2002). La importancia del calor no es un obstáculo mayor, las temperaturas elevadas son realmente necesarias solamente durante el periodo de maduración (al menos 20°C durante 25 a 40 días). En las regiones mediterráneas donde el invierno puede ser fresco, el arroz soporta las variaciones estacionales de temperatura siempre y cuando el aporte de agua sea regular y abundante durante el crecimiento. En altitud, las variedades adaptadas soportan bajas temperaturas nocturnas (12 °C) (Nene, 2005; IDEAS, 2007)
Dependiendo del clima y del ciclo de las variedades, se obtienen entre 1 y 4 cosechas de arroz al año. En climas tropicales se obtienen generalmente dos cosechas al año, a veces tres como en Vietnam en el Delta del Mekong. En China se llegó a realizar hasta cuatro ciclos por año. En climas templados (en altitud), se obtiene una sola cosecha al año, durante los meses de septiembre y octubre en el hemisferio Norte y durante los meses de marzo y abril en el hemisferio Sur (Nene, 2005; IDEAS, 2007)
Hay dos sistemas principales de cultivo del arroz: el sistema de sequía o de secano en el cual el cultivo es crecido sobre tierra seca, al igual que otros cereales, y el sistema de riego o de inundación, en el que los arrozales son irrigados, alimentados por la lluvia o por la corriente de un estanque local de recepción, y por gravitación de un arrozal a otro (Lozano, 2002).
El arroz de secano es a menudo cultivado en áreas montañosas con precipitación pluvial natural y sin bordos o diques para embalsar el agua. Un período de lluvia asegurado de 3 a 4 meses es necesario si el cultivo es abastecido por la lluvia y el suministro de agua no es controlado (Diago, 2002). El segundo sistema es el húmedo o inundado, en el cual el arrozal es inundado y el cultivo es crecido en agua estancada desde la plantación hasta cerca de la cosecha, en campos rodeados de pequeños diques que pueden retener el agua hasta una profundidad que puede variar entre 0-25 cm (agua poco profunda) y 25-50 cm (profundidad media). Este arroz pluvial de bajo fondo se cultiva también en aguas profundas (50-100 cm). Los sistemas de producción del arroz se clasifican más específicamente de acuerdo a la ecología en términos de agua: (1) de altiplanicie o secano (2) regados, (3) de bajío de temporal o abastecido por la lluvia, y (4) de aguas profundas o anegado (Nene, 2005; IDEAS, 2007)
El cultivo de arroz puede ser atacado por diversos agentes como insectos, los cuales pueden llegar a tener poblaciones muy altas. Pero estás se regulan entre sí por la acción de sus enemigos naturales, el clima, y por las labores propias del cultivo. Hay que tener especial cuidado de no afectar el equilibrio en que se encuentran estas poblaciones para evitar que los insectos fitófagos se conviertan en plagas y afecten el cultivo (Pérez et al, 2002; IDEAS, 2007). Entre los agentes también se encuentran las malezas las cuales forman parte del ecosistema propio, y deben ser controladas para evitar su competencia con el cultivo (Cuevas et al, 2003).
Finalmente se encuentran las enfermedades que pueden ser de origen infeccioso como las causadas por hongos, bacterias, virus y nematodos; y las no infecciosas o desordenes nutricionales causadas por deficiencias o exceso de algún nutrimento. Para que se presente en las plantas cualquier tipo de enfermedad deben darse las condiciones adecuadas, además es de gran importancia tener en cuenta la susceptibilidad que puede tener el cultivo a cualquiera de estas enfermedades (Olmos, 2000).
Todos estos agentes que afectan el rendimiento del cultivo de arroz son denominados plagas. El manejo de plagas en el cultivo del arroz es una de las principales limitantes en la producción arrocera. A nivel mundial las plagas del arroz destruyen el 35% de la producción, distribuyéndose las pérdidas de las siguiente manera: 12% insectos dañinos, 12% patógenos, 10% malezas, y 1% vertebrados que se alimentan del grano (FAO, 2002; Nene, 2005; IDEAS, 2007)
Transformación de arroz
Se han logrado rápidos avances en la transformación genética del arroz, con importantes resultados en el mejoramiento genético de variedades elite especialmente de las subespecies japónica e indica (O. sativa), sin embargo también se ha trabajado más recientemente en la transformación de arroces africanos (O. glaberrima). Desde que se logro producir la primera planta transgénica de arroz a finales de los 80´s, varios protocolos para la transferencia de genes se han empleado con éxito para la introducción de genes foráneos al arroz, más de 60 cultivares de arroz pertenecientes a japónica, indica, javanica y cultivares africanos han sido transformados exitosamente (Cocking, 2000; Ignacimuthu et al, 2000; Vasil, 2005).
Para la transformación genética del arroz se ha empleado sistemas de transformación tanto directos como indirectos. Inicialmente se trabajo fuertemente con métodos directos como PEG, Electroporación y Biobalística, luego se comenzó a trabajar en la implantación del método indirecto en el cual se emplea a Agrobacterium tumefaciens. Entre los avances logrados en la biotecnología del arroz esta la implementación del uso del sistema de Agrobacterium que hasta hace poco estaba limitado a dicotiledóneas. Así el sistema Agrobacterium y Biobalística son los dos sistemas de transformación de plantas más empleados en la transformación de arroz (Nadolska-Orczk et al, 2000; Repellin et al, 2001; Bajaj & Mohanty, 2005).
Se ha logrado un enorme progreso en el desarrollo de protocolos de transformación reproducibles, se ha implementado el uso de promotores constitutivos y de promotores tejido-específico, el uso de marcadores de selección negativa y positiva, así como la producción de plantas libres de marcadores de selección. También se ha incursionado en la transformación empleando varios genes a la vez. Sin dejar de mencionar el uso de genes reporteros que ha sido de gran ayuda para el desarrollo de protocolos eficientes (Tyagi & Mohanty, 2000; Ignacimuthu et al, 2000; Vasil, 2005).
Las modificaciones producidas en el arroz han tenido diversos fines. Se han realizado modificaciones de tipo agronómico con el fin de modificar rasgos importantes en la planta como: aumentar el valor nutricional del arroz, reducción de pérdidas en el rendimiento causadas por diferentes factores abióticos y bióticos, tolerancia a herbicidas, y tolerancia a condiciones ambientales extremas (Giri & Laxm, 2000; Tyagi & Mohanty, 2000; Ignacimuthu et al, 2000). En tolerancia a herbicidas, lo mas trabajado en plantas transgénicas de arroz ha sido la tolerancia al glufosinato de amonio y al glifosato (Roy & Wu ,2001)
Entre las modificaciones genéticas realizadas al grano de arroz se encuentran como ejemplos, el arroz dorado que presenta contenidos altos de beta caroteno, el arroz fortificado con un gen de la ferritina para aumentar la concentración de hierro, y el arroz con aminoácidos esenciales y con lactoferrina. (Al-Babili & Beyyer, 2005; Li, 2006). También se han trabajado en arroces con altos niveles de tolerancia a diferentes condiciones ambientales de estrés: bajas temperaturas, sequía, acidez, salinidad (Roy & Wu, 2001; Agrawal et al,2003). En el caso de genes de resistencia a patógenos o plagas de arroz se han buscado en especial genes de resistencias a Pyricularia, Rhizoctonia y algunos virus de importancia económica. Los arroces modificados de interés sanitario principalmente se han obtenido para resistencia a insectos lepidópteros con genes de Bacillus thurigiensis (BT) (Bajaj & Mohanty,2005;Riaz, 2006).
Breve historia de la transformación genética de arroz
Los primeros trabajos desarrollados en arroz fueron destinados al cultivo de tejidos y a establecer protocolos para la regeneración de este a partir de diferentes explantes. Entre 1976 y 1986 se publicaron seis artículos que reportaban la producción de callo a partir de protoplastos de arroz, y entre 1985 y 1990 se dieron 13 publicaciones relacionadas con la regeneración de las plantas de arroz a partir de protoplastos. Inicialmente la mayoría de los trabajos se realizaron con variedades japónica, y posteriormente se dio con variedades indicas, esto debido a que desde un principio los cultivares pertenecientes a esta subespecie mostraron ser de difícil manejo y poca respuesta al cultivo de tejidos, hasta el punto de ser llamados recalcitrantes (Tyagi & Mohanty, 2000; Cocking, 2000)
Una vez se contaba con trabajos previos en esta área se comenzó a trabajar en la transformación genética de arroz, ya que la regeneración de plantas fértiles de arroz a partir de protoplastos le abrió las posibilidades a la ingeniería genética de arroz. Los métodos escogidos para la transformación de arroz fueron la electroporación y el PEG, usando protoplastos (Tyagi & Mohanty, 2000; Bajaj & Mohanty,2005)
En 1988, Toriyama y colaboradores, así como Zhang y Wu, recuperaron los primeras plantas transgénicas de arroz empleando PEG. En ese mismo año Zhang y colaboradores, lograron la recuperación de plantas de arroz transgénicas usando electroporación. Estos trabajos solo tuvieron aplicación en la subespecie japónica. Los primeros en recuperar plantas transgénicas de arroz fértiles fueron Shimamoto y colaboradores en 1989 usando electroporación en la subespecie japónica y Datta et al, un año después en 1990 empleando PEG en arroz indica. En 1994, Biswas y colaboradores, obtuvieron por primera vez plantas transgénicas de arroz indica usando electroporación (Tyagi & Mohanty, 2000; Ignacimuthu et al, 2000; Bajaj & Mohanty,2005).
También se emplearon otros métodos de transformación para la transformación de arroz como el método del tubo polínico empleado por lou & wu en 1988, microinyección por Potrikus y colaboradores, en 1995 y la imbibición de semillas en una suspensión de DNA por Aryon et al, en 1991(Vasil , 2005).
Posterior a estos avances, estos métodos fueron ampliamente empleados para la transformación de arroz por varios grupos. Sin embargo, la regeneración de plantas fértiles a partir de protoplastos es lenta, laborioso y genotipo-dependiente. Existen otros problemas tales como variación somaclonal, integración de múltiples copias y regeneración de plantas albinas. Por lo tanto el uso de estos dos métodos decayó casi por completo y se comenzó a explorar otras posibilidades (Tyagi & Mohanty, 2000; Vasil, 2005).
El método de transformación directa de plantas llamado Biobalística, desarrollado por Sanford y colaboradores en 1987 (Sanford, 2000), se convirtió en el método de transformación de arroz más empleado. Debido a que era considerado un mecanismo universal por que permite introducir DNA sin necesidad de vectores especializados, en cualquier tipo de tejido o célula alcanzando capas profundas (Altpeter et al, 2005).
Christou y colaboradores en 1991 obtuvieron a partir de embriones inmaduros de cultivares de japónica, indica y javanica, las primeras plantas transgénicas de arroz desarrolladas mediante Biobalística. En 1996 Siamavni y colaboradores, reportaron la transformación de variedades elite de arroz indica, mediante el bombardeo de callos embriogénicos. Un avance significativo fue la transformación realizada por Chen y colaboradores en 1998 en arroz japónica con múltiples genes utilizando Biobalística; Bombardearon tejido de arroz con 14 plásmidos diferentes. Posteriormente Tang y colaboradores en el año 1999, transfirieron cuatro genes al genoma del arroz, mediante co-transformación empleando Biobalística. Dos de los cuatro genes utilizados eran de importancia económica, ya que proporcionaron resistencia contra el tizón bacteriano e insectos chupadores (Christou, 1997; Repellin et al, 2001; Chen et al, 2009).
El método indirecto de transformación de plantas que comenzó a ser estudiado para ser empleado en la transformación de plantas de arroz fue el sistema Agrobacterium. Este sistema se basa en la capacidad única que posee Agrobacterium tumefaciens para transferir e integrar en el genoma de las células vegetales DNA (T-DNA) proveniente de un plásmido de la misma bacteria. Agrobacterium tumefaciens naturalmente infecta plantas dicotiledóneas, razón por la cual este método no se había intentado emplear para la transformación de monocotiledóneas, consideradas como fuera del espectro de hospederos naturales de Agrobacterium, además de ser consideradas como recalcitrantes a esta tecnología (Pitzschke & Hirt, 2010; Ignacimuthu & Raveendar, 2011; Pacurar et al, 2011)
Por lo anterior las plantas monocotiledóneas permanecieron por muchos años inaccesibles a la manipulación genética mediada por Agrobacterium, sin embargo entre 1987 y 1992 varios laboratorios en todo el mundo hicieron un esfuerzo enorme para lograr transformar plantas de arroz. Pero el éxito era limitado y controvertido principalmente porque no se obtuvo un número significativo de plantas transgénicas y no se había comprobado completamente la integración del transgen. Pero aun así se puede afirmar que la primera planta transgénica de arroz obtenida por el sistema Agrobacterium fue en 1992 por Chan y colaboradores en la subespecie japónica (Roy et al, 2000; Shrawat & Lörz, 2006; Chen et al, 2009).
En 1994 Hiei y colaboradores, lograron resolver la controversia al obtener plantas transgénica de arroz japónica, empleando callos embriogénicos de tres semanas de edad derivados del escutelo y usando la cepa de Agrobacterium tumefaciens LBA4404. Solo hasta 1996 Rashid y colaboradores, lograron la transformación mediante Agrobacterium de cultivares de arroz índica y en este mismo año Dong y colaboradores, obtuvieron plantas transgénicas de arroz javanica (Repellin et al, 2001; Vasil , 2005; Chen et al, 2009).
Ahora se entiende que una serie de factores son de vital importancia para la transformación de arroz mediada por Agrobacterium, y son estos requisitos técnicos los que explican por qué inicialmente fue tan difícil de aplicar esta técnica a plantas monocotiledóneas. En los últimos años la transformación de plantas de arroz mediada por Agrobacterium se ha convertido en una técnica confiable y altamente reproducible para la transferencia de genes de interés al genoma en el arroz, además de ser el sistema preferido por las ventajas que presenta frente a las otras técnicas de transformación empleadas (Tyagi & Mohanty, 2000; Ignacimuthu et al, 2000; Bajaj & Mohanty,2005).
En la siguiente tabla se resumen los desarrollos más sobresalientes en la historia de la transformación de plantas de arroz (Tabla 1).
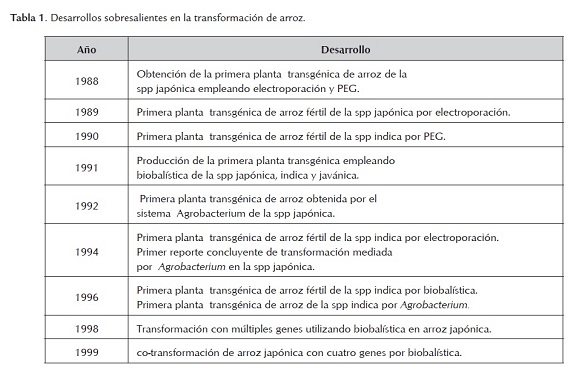
Es importante anotar que el arroz fue seleccionado como modelo para la investigación del genoma en los cereales debido al tamaño pequeño de su genoma y la importancia mundial que tiene como alimento. Es la primera planta cultivable que todo su genoma ha sido secuenciado, al ser secuenciado el genoma de las dos principales subespecies cultivables japónica e indica. En la actualidad, sirve como un genoma de referencia para estudios biológicos, moleculares, y para entender la biología de las plantas, y en especial para proporcionar una mejor visión y entendimiento de las monocotiledóneas (Shimamoto & Kyozuka, 2002; Gowda et al, 2003; Xu et al, 2005; Jiang et al, 2011)
Introducción de genes de importancia agronómica.
Una vez que se logró la transformación de las dos especies cultivadas de arroz Oryza sativa y Oryza glaberrima, ya sea empleando biobalística o Agrobacterium, se dio un auge en la transformación de diferentes cultivares de arroz, y en especial de variedades elite empleando genes de interés que permiten mejoras sustanciales en el cultivo, sin dejar de lado el uso de genes reporteros y transformaciones puramente académicas.
Debido a que la producción del cultivo del arroz se tiene que estar incrementando para satisfacer la demanda de la creciente población mundial, los esfuerzos en el mejoramiento genético del arroz mediante la biotecnología se han centrado en aumentar la producción mediante la reducción de pérdidas en las cosechas causadas por factores bióticos y abióticos. Asimismo, se ha trabajado en el aumento de la producción a través de mejoras en el rendimiento (Repellin et al, 2001; Lu & Yang, 2009).
También se ha trabajado en aumento en la calidad nutricional del grano, ya que al ser el arroz el alimento básico de la mayoría de la población mundial, se puede ayudar a reducir la desnutrición. Se ha intentado la producción de compuestos recombinantes para diferentes usos especialmente nutracéuticos. En la tabla 2 se enumeran los diferentes factores biótico y abióticos así como otras características importantes que se han buscado modificar o mejorar a través de la ingeniería genética de plantas en el cultivo de arroz (Datta, 2004; Bajaj & Mohanty,2005).

Factores bióticos
Las interacciones que se presentan en un cultivo con los factores bióticos (insectos, hongos, bacterias, virus, animales) pueden resultar ser los elementos más limitantes para la producción, ya que estos pueden ocasionar grandes pérdidas que pueden llevar hasta el detrimento total del cultivo. En el cultivo del arroz los factores bióticos más limitantes son los insectos y los organismos causantes de enfermedades, razón por la cual la búsqueda de resistencia a estos ha sido foco central para el mejoramiento convencional y para la ingeniería genética de plantas.
Resistencia a insectos
Se ha establecido que entre el 10 y 13 % son las pérdidas anuales en el cultivo del arroz causadas por insectos, y que el empleo de insecticidas químicos son la principal herramienta empleada para combatir el ataque de insectos a los cultivos. Sin embargo el uso indiscriminado de insecticidas ha generado aumento en los costos de producción, riesgos para la salud, contaminación ambiental y hasta preocupación por la seguridad alimentaria (Babu et al, 2003; Chen et al, 2009; Deka & Barthakur, 2010).
Se ha buscado entre el genoma del arroz genes que permita resistencias estables, duraderas y contra diversas clases de insectos, pero no se han encontrado. Además usar las técnicas de mejoramiento convencional para este propósito toma mucho tiempo. Por lo anterior se ha implementado el uso de la ingeniería genética de plantas con el fin de buscar algunas soluciones a este problema (Tyagi & Mohanty, 2000; Babu et al, 2003; Deka & Barthakur, 2010).
En el arroz se han introducidos varios genes con el fin de lograr resistencia a los insectos plaga. Los genes más empleados con este propósito son los genes Bt o genes cry provenientes Bacillus thuringiensis, que codifican para una δ-endotoxina. La importancia de estos genes radica en la toxicidad de la proteína codificada contra larvas de insectos-plaga de los ordenes lepidóptero, coleóptero, díptero, himenóptero, homóptero, ortóptero, malófago, y contra organismos como ácaros, platelmintos y nematodos. Existen más de 500 genes cry descritos y clasificados en 67 grupos. Los genes cry más empleados en transformación de arroz para generar resistencia contra las principales plagas del cultivo son: Cry1A, Cry1Ab, Cry1Ac, Cry1C y Cry2A. (Soberon et al, 2007; Sauka & Benintende, 2008; Deka & Barthakur, 2010; Bravo et al, 2011)
En el arroz se han empleado los genes cry especialmente en contra de insectos lepidópteros. Se pueden contar más de 50 variedades de arroz que han sido transformadas con estos, principalmente japónicas, y unas pocas indicas. La primera planta transgénica de arroz japónica con genes Bt que se obtuvo fue en 1993 por Fujimito y colaboradores. En 1996 se obtuvo la primera planta con genes Bt de la subespecie indica por Wunn y colaboradores. Se han reportado evaluaciones de campo de arroz Bt desde el año 2000, y fue Irán el primer país en autorizar siembras y sembrar arroz Bt con fines comerciales en el año 2004 (James, 2005). Actualmente China e India adelantan evaluaciones de campo de arroz Bt. Hasta ahora, ningún país cultiva con fines comérciales arroz transgénico (Tyagi & Mohanty, 2000; Babu et al, 2003; High et al, 2004; Deka & Barthakur, 2010).
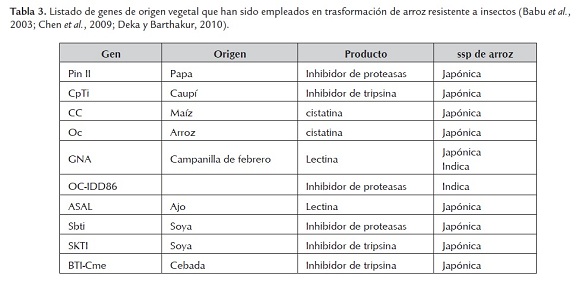
También se han empleado otros genes de origen vegetal para generar resistencia a insectos en el cultivo del arroz, entre estos genes están los inhibidores de proteasas y las lectinas. En general los productos de estos genes actúan interfiriendo en los procesos digestivos del insecto. En la tabla 3 están listados los genes de origen vegetal con propiedades insecticidas que se han empleado en la transformación de arroz. Adicionalmente se ha transformado una variedad japónica con un gen de origen animal proveniente de araña con características insecticidas, el gen SpI (Babu et al, 2003; Chen et al, 2009; Lu & Yang, 2009; Deka & Barthakur, 2010).
Resistencia a enfermedades
Los ataques de agentes patógenos causan entre el 10 al 40 por ciento de reducción en el rendimiento de los cultivos, dependiendo de la ubicación geográfica de estos. De ahí la importancia de desarrollar cultivos resistentes a enfermedades, esperando que aumenten los rendimientos de los cultivos, así como reducir la cantidad de agroquímicos utilizados disminuyendo costos para los agricultores (Babu et al, 2003; Chen et al, 2009; Ahmab etal, 2011)
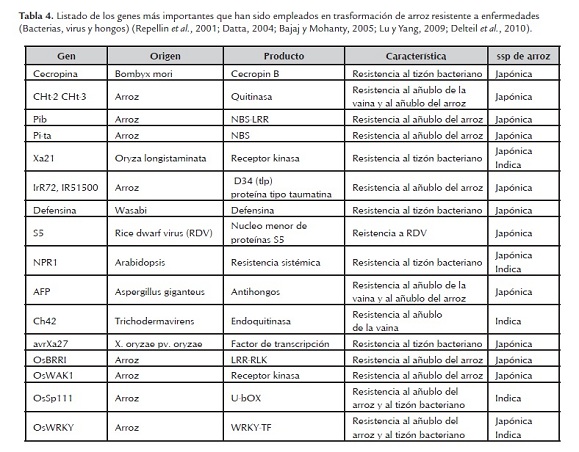
En el arroz existen varias familias de genes que son responsables de la resistencia o tolerancia a diferentes enfermedades que atacan el cultivo. Para las principales enfermedades del arroz se han identificado varios genes. Más de 30 genes de resistencia al tizón bacteriano se han identificado en el genoma del arroz, entre ellos, seis han sido clonados (XA1, XA3 / Xa26, xa5, xa13, Xa21 y Xa27), y más de 60 genes de resistencia al añublo del arroz han sido identificados, de los cuales 10 han sido clonados (Pib, Pi-d2, Pikm, Pi-ta, Pizt, Pi 2, PI5, Pi9, Pi36,y Pi37) (Giri & Laxm, 2000; Babu et al,2003; Chen et al, 2009; Delteil et al, 2010).
Cuando se habla de transformación genética empleando genes de resistencia hay que tener mucho cuidado, debido a que se está trabajando con familias de múltiples genes que están muy asociadas entre sí. Por consiguiente es muy importante la conservación de las vías de resistencia, no solo para que pueda funcionar la resistencia, si no para que se permita la transferencia de genes, ya que la transferencia de un gen de una especie a otra puede dar lugar a efectos inesperados (Tyagi & Mohanty, 2000; Giri & Laxm, 2000; Leung et al, 2003; Delteil et al, 2010).
Hay más de 60 genes de diferente procedencia que han sido sobre-expresados o mutados en el arroz, que han demostrado efectos beneficiosos con respecto a la resistencia a enfermedades. 41 de estos genes pertenecen al arroz, 5 a otras especies vegetales y 9 de origen no vegetal, los demás casos son piramidación de genes o genes sintéticos. En la tabla 4 se muestran los principales genes que se han empleado en transformación de arroz con resistencia a enfermedades. Un 75% de los casos de plantas transgénicas se hicieron en la subespecie japónica, y en la mayoría de las plantas transformadas la resistencia fue obtenida por la sobre-expresión de genes (Repellin et al, 200; Leung et al, 2003; Chen et al, 2009; Delteil et al, 2010).
Entre las estrategias empleadas para la producción de plantas transgénicas resistente a enfermedades aparte de la introducción de genes de resistencia, también está la sobreexpresión de las proteínas relacionadas con patogenicidad como las quitinasa, las Β-1,3-glucanasas, proteínas tipo taumatina, además de proteínas de anti-fúngicas de origen vegetal o bacteriano. Para generar resistencia a virus se han empleado genes que codifican para proteínas del mismo virus (Bajaj & Mohanty, 2005; Chen et al, 2009;Delteil et al, 2010;Ahmab et al, 2011).
Se han logrado importantes avances en la producción de arroz transgénico resistente a las diferentes plagas del arroz, sin embargo ahora el principal interés está en la producción de plantas resistentes contra múltiples patógenos, que muestren una alta y duradera resistencia (Giri & Laxm, 2000; Babu et al, 2003; Chen et al, 2009; Delteil et al, 2010).
Factores abióticos
Presiones ambientales tales como sequía, salinidad y temperaturas extremas son factores limitantes en la productividad de las plantas. Las plantas han desarrollado diferentes estrategias fisiológicas y bioquímicas para adaptarse a condiciones de estrés en respuesta a varios entornos. Con el propósito de buscar niveles más altos de producción, ampliar las posibilidades de terrenos productivos y estar un paso a delante en cuanto al cambio climático, la tolerancia a factores abióticos se ha convertido en el principal tema de investigación y estudio de los últimos años (Tyagi & Mohanty, 2000; Sreenivasulu et al, 2007; Chen et al, 2009).
En cuanto al cultivo del arroz la introducción de tolerancias a estreses abióticos ha sido el blanco principal en las transformaciones genéticas que se han venido desarrollando en los últimos años. Aunque los trabajos se han enfocado en tolerancia a sequia y salinidad, también se está trabajando en otros estreses como las bajas temperaturas y los golpes de calor.
Tolerancia a sequia y salinidad
La productividad de las plantas está fuertemente influenciada por los factores ambientales. En el cultivo del arroz factores como déficit hídrico y las altas concentraciones de sal en los suelos, son los factores más importantes y que mayor incidencia tiene en la reducción del rendimiento de la producción de los cultivos de arroz (Cattivelli et al, 2008; Dolferus et al, 2011;).
El déficit hídrico puede ser el factor que más afecta el cultivo del arroz, ya que consume grandes cantidades de agua, este ha comenzado a ser un factor limitante en la producción, más aún con el efecto del cambio climático. El desarrollo de variedades de arroz tolerantes a sequía es crucial para incrementar los rendimientos, reducir el consumo de agua en el cultivo, y contribuir a garantizar la seguridad alimentaria (Bajaj & Mohanty, 2005; Chen et al, 2009; Ahmab etal, 2011).
En los últimos años se ha venido trabajando para entender cómo funcionan los mecanismos de resistencia a los diferentes estreses ambientales que desarrollan las plantas. Se ha logrado identificar varios genes que están relacionados con las diferentes estrategias que las plantas utilizan con este fin. En la aclimatación al déficit hídrico y a la alta salinidad las plantas emplean como estrategia la acumulación de solutos compatibles u osmolitos, como glicina, betaina, prolina y trealosa. A nivel fisiológico la adaptación de las plantas a la alta salinidad proviene de proteínas implicadas en el transporte de iones y agua. Cabe anotar que las estrategias empleadas por las plantas no solo son eficaces para un estrés determinado, se ha visto que pueden influir contra varios estreses ambientales (Roy et al, 2000; Ignacimuthu et al, 2000; Cattivelli et al, 2008; Lu & Yang, 2009; Chen et al, 2009; Dolferus et al, 2011)
Para el desarrollo de plantas transgénicas de arroz que presenten toleración a sequia y/o a salinidad, se han empleado estrategias como la introducción de genes y/o la expresión inducible de proteínas, que estén relacionadas con las estrategias que emplean las plantas para enfrentar los diferentes estreses abióticos. Sin embargo, cabe señalar que la expresión constitutiva de estos factores puede causar un fenotipo aberrante por lo que estos genes tienen que expresarse bajo regulación de promotores inducibles (Bajaj & Mohanty, 2005; Chen et al, 2009; Ahmab et al, 2011).
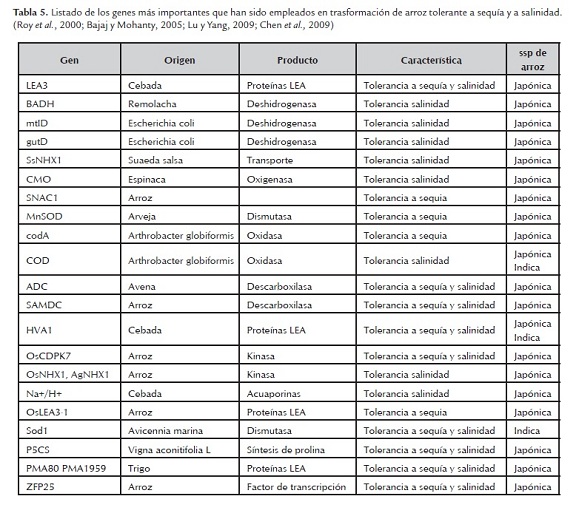
También se ha sugerido que el efecto que tiene cada gen en la adquisición de tolerancia al estrés abiótico es más bien pequeña, por lo tanto se ha propuesto un enfoque multigénico que puede conferir tolerancia mucho mayor. Actualmente se han identificado más de 70 genes relacionados con tolerancia a estreses abióticos. En la Tabla 5 se muestran los principales genes que se han empleado en transformación de arroz con tolerancia a sequia y salinidad (Roy et al, 2000; Bajaj & Mohanty, 2005; Chen et al, 2009; Ahmab et al, 2011).
Otras características
Cabe mencionar que han sido muchas las características que se han buscado mejorar, modificar e introducir al cultivo del arroz, y por lo tanto la lista de transformaciones en arroz es larga. Entre este grupo se puede señalar como los más sobresalientes: las mejoras en la calidad nutricional del grano de arroz y la tolerancia a herbicidas.
Calidad nutricional
El arroz es la principal fuente de hidratos de carbono y proteínas, su consumo puede ser suficiente para sobrevivir, pero es una fuente pobre para muchos de los micronutrientes esenciales en la nutrición humana. Una dieta poco variada y la dependencia en el arroz como principal fuente de alimento, son causa de malnutrición en gran parte del mundo en desarrollo principalmente en las regiones donde hay un alto consumo de arroz (Asia, India, África y Latinoamérica) (Dawe et al, 2002; Stein et al, 2008; Ahmab et al, 2011). Estas deficiencias reducen la productividad en el trabajo, generan disminución de la capacidad mental, retraso en el crecimiento, ceguera, aumento de la mortalidad infantil, afectada la integridad epitelial contra las infecciones, y la reducción de la respuesta inmune, entre otros efectos (Welch & Gram, 2004; WHO & FAO, 2004).
Por lo tanto, un aumento en la calidad nutricional del arroz seria de gran importancia y una solución parcial para los problemas de salud de gran parte de la población mundial. Varias estrategias se han planteado con este fin, y es aquí donde la producción de plantas transgénicas de arroz con mejoras en la calidad nutricional del grano comienza a ganar importancia. Los blancos principales para la mejora genética del arroz son los componentes del grano como los minerales, las vitaminas y las proteínas. Específicamente se ha buscado aumentar los niveles de aminoácidos esenciales como la lisina, de micronutrientres como el hierro, el zinc y el Β caroteno, precursor de la provitamina A (Roy et al, 2000; Giri & Laxm, 2000; Tyagi & Mohanty, 2000; Bajaj & Mohanty, 2005; Chen et al, 2009). En la tabla 6 se resumen los principales genes que han sido empleados para la transformación genética de arroz con mejoras en la calidad nutricional.
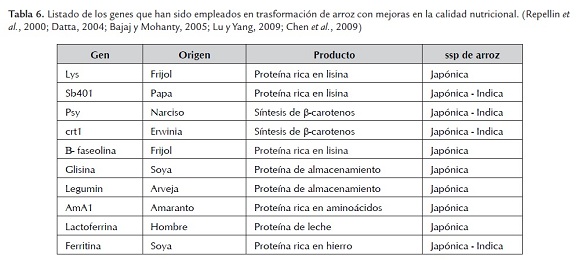
No se puede dejar de mencionar el más importante desarrollo de la ingeniería genética de plantas, el Arroz Dorado. Este es un arroz modificado genéticamente para acumular en su embrión el precursor de la vitamina A, llamado provitamina A o beta-caroteno, además de otros carotenos de importancia nutricional. Este ß-caroteno es el que le otorga un característico y peculiar color dorado dándole su nombre (Al-Babili & Beyer, 2005; Schaub et al, 2005; Bajaj & Mohanty, 2005).
El Arroz Dorado se produjo por la introducción del gen que codifica para la fitoeno- sintasa de Narcissus, y del gen que codifica para la fitoeno-deshidrogenasa de Erwinia (Al-Babili & Beyer, 2005; Mayer, 2007); con el fin de modificar la ruta metabólica de producción de Β carotenos en el endospermo del arroz. Consiguiéndose completar la única ruta metabólica entera que se haya insertado en una planta por medio de métodos de ingeniería genética, demostrando que era posible diseñar la ruta bioquímica completa. Inicialmente se desarrollo en cultivares de la subespecie japónica y más recientemente se ha trabajado en cultivares de la subespecie indica. (Ye et al, 2000; Potrykus, 2001; Ahmab et al, 2011).
Se han producido tres generaciones de Arroz dorado, la primera en 1999 llamada prototipo o GR (Golden Rice) (Ye et al, 2000; Mayer, 2007). La segunda se obtuvo en el 2003, que fue la primera línea mejorada de arroz dorado (SGR1) la cual presentaba niveles más altos de provitamina A, y además los genes estaban bajo el control del promotor tejido específico para endospermo de la glutelina (Potrykus, 2001; Mayer, 2007). En 2005 se presentó la tercera generación de arroz dorado o SGR2, la cual produce 23 veces más de provitamina A que la versión inicial, es decir 37 μg/g de provitamina A. Esta versión se diferencia de las anteriores por la utilización de genes de gramíneas como arroz o maíz para reemplazar el gen de la fitoeno sintasa de Narcissus pseudonercissus (Al-Babili & Beyer, 2005; Paine et al, 2005).
El Arroz Dorado busca ser una fuente de vitamina A para que las poblaciones que no consumen la suficiente cantidad de esta vitamina imprescindible en su dieta diaria, y de este modo contribuir a disminuir la avitaminosis en los países en vías de desarrollo. La investigación fue financiada por la Fundación Rockefeller, el Instituto Federal Suizo de Tecnología (Zurich), el programa Biotech de la Comunidad Europea y la Oficina Federal Suiza de Educación y Ciencia (Potrykus, 2001; Mayer, 2007).
Tolerancia a herbicidas
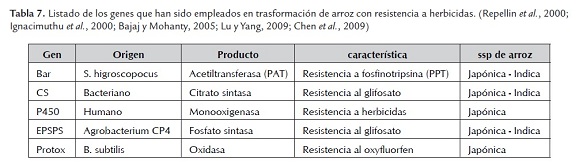
Las malezas compiten con los cultivos por los nutrientes disponibles y la energía lumínica, reduciendo el rendimiento de los cultivos entre el 10 y 15%. En algunos cultivos las malezas pueden ser controladas de manera eficiente mediante el empleo de herbicidas, pero en la mayoría de los cultivos esta posibilidad está restringida por la incapacidad de los herbicidas para distinguir entre las plantas de cultivo y las malezas (Repellin et al, 2000; Bajaj & Mohanty, 2005; Lu & Yang, 2009; Chen et al, 2009; Ahmab et al, 2011)
A través de la biotecnología, mediante la producción de plantas transgénicas ha sido posible la manipulación genética de la tolerancia a herbicidas, lo que permite la eliminación selectiva de las malezas en los campos cultivados. Convirtiéndose la tolerancia a los herbicidas en el rasgo más empleado en los cultivos transgénicos, con la mayor área cultiva a nivel mundial y de mayor crecimiento anual. Para la transformación genética de plantas tolerante a herbicidas se han empleado tres estrategias: 1) introducir genes de la enzima diana con reducida afinidad por el herbicida; 2) alterar el lugar de acción de herbicida, y 3) incorporar un gen que detoxifique al herbicida (Ignacimuthu et al, 2000; Villalba, 2009; Chen et al, 2009).
Los trabajos en transformación de plantas tolerantes herbicidas se ha centrado en herbicidas con alta actividad, baja toxicidad, escasa movilidad en los suelos, biodegradación rápida y amplio rango de acción. La principal fuente de genes de resistencia a herbicidas se encuentra en las bacterias (Ignacimuthu et al, 2000; Bajaj & Mohanty, 2005; Villalba, 2009; Chen et al, 2009).
Desde 1991 cuando Christou y colaboradores produjeron plantas transgénicas de arroz resistentes a herbicidas de las subespecies indica y japónica, se ha venido produciendo un gran número de cultivares de arroz con resistencia herbicida. Sin embargo, por percepción pública y legislación no han salido al mercado. En la Tabla 7 se listan los principales genes de resistencia a herbicidas que se han empleado en transformación genética de arroz (Giri & Laxm, 2000; Tyagi & Mohanty, 2000; Bajaj & Mohanty,2005; Chen et al, 2009).
Conclusiones
La importancia del arroz, se puede medir con las siguientes cifras: cultivado en 113 países, es el alimento básico para la mitad de la población humana, emplea casi mil millones de personas, y se han desarrollado más de 2000 diferentes variedades. Se siembran dos especies O. sativa y O. glaberrima. De la primera, se reconocen dos subespecies, japónica e indica. Los dos principales sistemas de producción son el de arroz secano, y el arroz de riego. Por efecto del ataque de las plagas se pierde cerca del 35% de la producción (12% por insectos, 12% por patógenos, 10% por animales, 1 % por animales vertebrados). Para enfrentar estos problemas desde finales de la década de los 80´s del pasado siglo XX, se viene trabajando en el desarrollo de sistemas de ingeniería genética para la transferencia de genes foráneos al genoma del arroz. Los principales hitos de esta historia son los siguientes: 1988, primera planta transgénica; 1989, primera planta transgénica fértil de japónica usando electroporación; 1990, primera planta transgénica fértil de indica usando PEG; 1991, implementación de la biobalística para japónica e indica; 1992, implementación de Agrobacterium tumefaciens en japónica; 1999, co-transformación de una variedad japónica con 4 genes diferentes mediante biobalística. El progreso en la apropiación de la tecnología del DNA recombinante para el cultivo del arroz, ha incluido el desarrollo de protocolos reproducibles, el uso de genes reporteros, el uso de marcadores de selección positiva y de selección negativa, la implementación del uso de promotores constitutivos y de promotores tejido-especifico, la producción de plantas transgénicas libres de marcadores de selección, y la piramidación de genes. Se han realizado modificaciones para incrementar el valor nutricional, para reducir las pérdidas en el rendimiento causada por diferentes agentes bióticos y abióticos, la tolerancia a herbicidas, y la tolerancia a condiciones ambientales extremas. La integración de diversas aproximaciones de las "omicas" con la ingeniería genética, en el contexto del denominado "Smart Breeding", augura una profundización del uso del conocimiento, para la solución de los problemas del cultivo del arroz y de la agricultura en general.
Agradecimientos
Este trabajo se realizo en el marco del convenio específico de cooperación técnica y científica celebrado entre la Federación Nacional de Arroceros (FEDEARROZ) y la Universidad Nacional de Colombia, para la ejecución del proyecto "Desarrollo de sistemas de ingeniería genética para cultivares colombianos de arroz". Los autores agradecen a las dos instituciones por el apoyo recibido.
Referencias bibliográficas
1 Agrawal G. 2003. Diverse environmental cues transiently regulate OsOPR1 of the octadecanoid pathway; revealing its importance in rice defense/stress and development. Biochemical and Biophysical Research Communications. 310(4): p. 1073-1082. [ Links ]
2 Ahmad P., Ashraf M., Younis M., Hu X., Kumar A., Akram N., Nudrat A. & Al-Qurainy F. 2011. Role of transgenic plants in agriculture and biopharming. Biotechnology Advances. Article in press. doi:10.1016/j.biotechadv.2011.09.006 [ Links ]
3 Altpeter F., Baisakh N., Beachy R., Bock R., Teresa Capell T, Christou P., Daniell H., Datta K., Datta S., Philip J., Dix P., Fauque C., Huang N., Kohli A., Mooibroek H., Nicholson L., Nguyen T., Nugent G., Raemakers K., Romano A., Somers D., Stoger E., Taylor N. & Visser R. 2005. Particle bombardment and the genetic enhancement of crops: myths and realities Molecular Breeding. 15: p. 305-327 [ Links ]
4 Al-Babili S. & Beyer P. 2005.Golden Rice - five years on the road - five years to go? Trends in Plant Science. 10(12): p. 565-573. [ Links ]
5 Babu R., Sajeena A., Seetharaman K. & Reddy M. 2003. Advances in genetically engineered (transgenic) plants in pest management-an over view. Crop Protection. 22: p. 1071-1086 [ Links ]
6 Bajaj S. & Mohanty A. 2005. Recent advances in rice biotechnology-towards genetically superior transgenic rice. Plant Biotechnology Journal. 3: p. 275-307 [ Links ]
7 Benavides H. & Segura O. 2005. El Entorno Internacional del Sector Arrocero Centroamericano. IICA/Unidad de Políticas y Negociaciones Comerciales, Federación Centroamericana del Arroz. Costa Rica. p. 16-38 [ Links ]
8 Bravo A., Likitvivatanavong S., Gill S. & Soberón M. 2011. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochemistry and Molecular Biology. 41: p. 423-431 [ Links ]
9 Cattivelli L., Rizza F., Badeck F., Mazzucotelli E., Mastrangelo A., Francia E., Mare C., Tondelli A. & Stanca A. 2008. Drought tolerance improvement in crop plants: An integrated view from breeding to genomics. Field Crops Research. 105: p. 1-14 [ Links ]
10 CIAT. 2005. Guía de estudio: Morfología de la planta de arroz. Centro Internacional de la Agricultura Tropical. Colombia. p. 16. Sitio web: http://www.ciat.cgiar.org/riceweb/esp/inicio.htm [ Links ]
11 Cocking E. 2000. Progress in Rice Biotechnology. Cahiers Options Méditerranéennes. 8: p. 61-65 [ Links ]
12 Chen H., Lin Y., & Zhang Q. 2009. Review and prospect of transgenic rice research. Chinese Sci Bull.54: p. 4049-4068 [ Links ]
13 Christou P. 1997. Rice transformation: bombardment. Plant Molecular Biology. 35: p. 197-203 [ Links ]
14 Cuevas A., Salive A., Puentes B. & Bastidas H. 2003. Manejo integrado de malezas. FEDEARROZ Fondo Nacional del Arroz. Colombia. p. 52 [ Links ]
15 Datta S. 2004 .Rice Biotechnology: A Need for Developing Countries. AgBioForum. 7(1&2): 31-35 [ Links ]
16 Dawe D., Robertson R. & Unnevehr L. 2002. Golden rice: what role could it play in alleviation of vitamin A deficiency. Food Policy. 27: p. 541-560 [ Links ]
17 Degiovanni V.,Martinez C. & Motta F. 2010. Produccion Eco-Eficiente del arroz en America Latina. Tomo I. Cali- Colombia. Centro Internacional de la Agricultura Tropical CIAT. p. 35-59 [ Links ]
18 Deka S. & Barthakur S. 2010. Overview on current status of biotechnological interventions on yellow stem borer Scirpophaga incertulas (Lepidoptera: Crambidae) resistance in rice. Biotechnology Advances. 28: p. 70-81 [ Links ]
19 Delteil A., Zhang J., Lessard P. & Morel J. 2010. Potential Candidate Genes for Improving Rice Disease Resistance. Rice. 3: p. 56-71 [ Links ]
20 Diago M. 2002. El arroz un regalo de Dios. FEDEARROZ Fondo Nacional del Arroz. Colombia. p. 18 [ Links ]
21 Dolferus R, Ji X. & Richards R. 2011. Abiotic stress and control of grain number in cereals. Plant Science. 181: p. 331- 341 [ Links ]
22 Garris A., Tai T., Coburn J., Kresovich S. & McCouch S. 2005. Genetic Structure and Diversity in Oryza sativa L. Genetics. 169: p. 1631-1638 [ Links ]
23 Giri C. & Laxm V. 2000.Production of transgenic rice with agronomically useful genes: an assessment. Biotechnology Advances. 18: p. 653 - 683 [ Links ]
24 Gowda M., Venu R., Roopalakshmi K., Sreerekha M. & Kulkarni S. 2003. Advances in rice breeding, genetics and genomics. Molecular Breeding. 11: p. 337-352 [ Links ]
25 High S., Cohen M., Shu Q. & Altosaar I. 2004. Achieving successful deployment of Bt rice. Trends in Plant Science. 9 (6): p. 287-292 [ Links ]
26 Ignacimuthu S., Arockiasamy S. & Terada R. 2000 .Genetic transformation of rice: Current status and future prospects. Current Science. 79 (2): p. 186-195 [ Links ]
27 Ignacimuthu S. & Raveendar S. 2011. Agrobacterium mediated transformation of indica rice (Oryza sativa L.) for insect resistance. Euphytica. 179: p. 277-286 [ Links ]
28 Ikehashi H. 2009. Why are There Indica Type and Japonica Type in Rice ? History of the Studies and a View for Origin of Two Types. Rice Science. 16 (1): p. 1-13 [ Links ]
29 Iniciativa de economía alternativa y solidaria (IDEAS). 2007. La producción y el comercio internacional del arroz Boletin 16. IDEAS España. p. 1-56 [ Links ]
30 James C. 2005. Global Status of Commercialized Biotech/GM Crops: 2005. ISAAA Briefs No. 34. ISAAA: Ithaca, NY. p. 1-13 [ Links ]
31 James C. 2010. Global Status of Commercialized Biotech/GM Crops: 2010. ISAAA Brief No. 42. ISAAA. Ithaca, NY. p 1-30 [ Links ]
32 Jiang Y., Cai Z., Xie W., Long T., Yu H. & Zhang Q. 2011. Rice functional genomics research: Progress and implications for crop genetic improvement. Biotechnology Advances. Article in press. doi:10.1016/j.biotechadv.2011.08.013 [ Links ]
33 Johns M. & Mao L. 2007. Differentiation of the two rice subspecies indica and japonica: a Gene Ontology perspective. Funct Integr Genomics. 7: p.135-151 [ Links ]
34 Khush S. 1997. Origin, dispersal, cultivation and variation of rice. Plant Molecular Biology. 35: p. 25-34 [ Links ]
35 Kumar V., Bellinder R., Brainard D., Malik R. & Gupta R. 2008. Risks of herbicide-resistant rice in India: A review. Crop Protection. 27: p. 320-329 [ Links ]
36 Leung H., Zhu Y., Fan Y., Chen H., Pangga I., Cruz C., Mew T. & Revilla-Molina I. 2003. Using Genetic Diversity to Achieve Sustainable Rice Disease Management. Plant Disease. 87(10): p. 1156-1169 [ Links ]
37 Li A. 2006. Screening for and Genetic Analysis on T-DNA-inserted Mutant Pool in Rice. Acta Genetica Sinica. 33(4): p. 319-329 [ Links ]
38 Londo J., Chiang Y., Hung K., Chiang T. & Schaal B. 2006. Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. PNAS. 103(25): p. 9578-9583 [ Links ]
39 Lozano R. 2002. El manejo integrado del cultivo del arroz. FEDEARROZ Fondo Nacional del Arroz. Colombia. p. 60- 82 [ Links ]
40 Lu B. & Yang C. 2009. Gene flow from genetically modified rice to its wild relatives: Assessing potential ecological consequences. Biotechnology Advances. 27: 1083-1091 [ Links ]
41 Mayer J. 2007. Golden rice, Golden crops, Golden prospects. Revista Colombiana de Biotecnología. 9 (1): p. 22- 34 [ Links ]
42 Nadolska-Orczk A., Orczvk W. & Ptvetakiewicz A. 2000. Agrobacterium-mediated transformation of cereals - from technique development to its application. Physiologiae Plantarum. 22 (1): p. 77-88 [ Links ]
43 Nene Y. 2005. Rice Research in South Asia through Ages. Asian Agri-History Foundation. 9 (2): p 85-106 [ Links ]
44 Olmos R. 2000. Guía de reconocimiento y manejo de las principales enfermedades del arroz. FEDEARROZ Fondo Nacional del Arroz. Colombia. p 52 [ Links ]
45 Organización de las Naciones Unidas (ONU). 2002. Noticiario de la Comisión Internacional del Arroz Vol. 51. FAO. Rome. p. 1-89 [ Links ]
46 Organización de las Naciones Unidas (ONU). 2004. 28ª Conferencia Regional de la FAO Para América Latina y el Caribe Año Internacional del Arroz. FAO. Guatemala. p. 1-26 [ Links ]
47 Pacurar D., Thordal-Christensen H., Pacurar M., Pamfil D., Botez C. & Bellini C. 2011. Agrobacterium tumefaciens: From crown gall tumors to genetic transformation. Physiological and Molecular Plant Pathology. Article in press. doi:10.1016/j.pmpp.2011.06.004 [ Links ]
48 Paine J., Shipton C., Chaggar S., Howells R., Kennedy M., Vernon G., Wright S., Hinchliffe E., Adams J., Silverstone A. & Drake R. 2005.Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nature Biotecnology. 23: p. 482-487 [ Links ]
49 Pérez C., Cuevas A. & Reyes L. 2001. Manejo integrado de insectos en el cultivo del arroz. FEDEARROZ Fondo Nacional del Arroz. Colombia. p 51 [ Links ]
50 Pitzschke A. & Hirt H. 2010. New insights into an old story: Agrobacterium induced tumour formation in plants by plant transformation. The EMBO Journal. 29: p. 1021-1032 [ Links ]
51 Potrykus I. 2001A. Golden Rice and Beyond. Plant Physiology.125: p. 1157-1161 [ Links ]
52 Potrykus I. 2001B. The "golden rice" tale. in vitro cell. Dev. Biol.Plant 37: p. 93-100 [ Links ]
53 Purugganan M. 2010. The evolution of rice: molecular vignettes on its origins and spread. Archaeol Anthropol Sci. 2: p. 61-68 [ Links ]
54 Rana R. 2004. Emerging Trends in Managing and Using Rice Genetic Resources. Asian Biotechnology and Development Review. E.U. p. 49-66. [ Links ]
55 Repellin A., Båga M., Jauhar P. & Chibbar R. 2001. Genetic enrichment of cereal crops via alien gene transfer: New challenges. Plant Cell, Tissue and Organ Culture. 64: p. 159-183 [ Links ]
56 Riaz N. 2006. Development of Indica Basmati rice harboring two insecticidal genes for sustainable resistance against lepidopteran insects. South African Journal of Botany. 72(2): p. 217-223 [ Links ]
57 Roy M., Jain R., Rohila J: & Wu R. 2000. Production of agronomically superior transgenic rice plants using Agrobacterium transformation methods: Present status and future perspectives. Current Science. 79 (7): p. 954-960 [ Links ]
58 Roy M. & Wu R. 2001. Arginine decarboxylase transgene expression and analysis of environmental stress tolerance in transgenic rice. Plant Science. 160(5): p. 869-875 [ Links ]
59 Saharam V., Yadav R., Yadav N. & Ram K. 2004A. Studies on improved Agrobacterium-mediated trasnformation in two Indica rice (Oryza sativa l.). African Journal of Biotechnology. 3 (11): p. 572-575 [ Links ]
60 Sanford J. 2000. The Development Of The Biolistic Process. in vitro Cell. Dev. Biol. Plant. 36: p. 303-308 [ Links ]
61 Sang T. & Ge S. 2007. Genetics and phylogenetics of rice domestication. Current Opinion in Genetics & Development. 17: p. 533-538 [ Links ]
62 Sauka D. & Benintende G.2008. Bacillus thuringiensis: generalidades. Un acercamiento a su empleo en el biocontrol de insectos lepidópteros que son plagas agrícolas. Revista Argentina de Microbiología. 40: p. 124-140 [ Links ]
63 Schaub P., Al-Babili S. & Drake R. 2005. Why Is Golden Rice Golden (Yellow) Instead of Red? Plant Physiology. 138: p. 441-450 [ Links ]
64 Shimamoto K. & Kyozuka J. 2002. Rice as a model for comparative genomics of plants. Annual Reviews. 53: p. 399-419 [ Links ]
65 Shrawat A. & Lörz H. 2006. Agrobacterium-mediated transformation of cereals: a promising approach crossing barriers. Plant Biotechnology Journal. 4: p. 575- 603 [ Links ]
66 SMIA. 2006. Perspectivas Alimentarías: Análisis de mercados. Sistema Mundial de Información y Alerta sobre la agricultura y alimentación. Argentina. p 35-39 [ Links ]
67 Soberón M., Pardo-López L., López I., Gómez I., Tabashnik B. & Alejandra Bravo A. 2007. Engineering Modified Bt Toxins to Counter Insect Resistance. 318: p. 1640-1642 [ Links ]
68 Sreenivasulu N., Sopory S. & Kavi-Kishor P. 2007. Deciphering the regulatory mechanisms of abiotic stress tolerance in plants by genomic approaches. Gene. 388: p. 1-13 [ Links ]
69 Stein A., Sachdev H. & Gaim M. 2008. Genetic Engineering for the Poor: Golden Rice and Public Health in India. World Development. 36(1): p. 144-158 [ Links ]
70 Tang L., Zou X., Achoundong G., Potgieter C., Second G., Zhang D. & Ge S. 2010. Phylogeny and biogeography of the rice tribe (Oryzeae): Evidence from combined analysis of 20 chloroplast fragments. Molecular Phylogenetics and Evolution. 54: 266-277 [ Links ]
71 Toriyama K, Heong K & Hardy B. 2005. Rice is life: scientific perspectives for the 21st century. Proceedings of the World Rice Research Conference held in Tokyo and Tsukuba, Japan, 4-7 November 2004. Los Baños (Philippines): International Rice Research Institute, and Tsukuba (Japan): Japan International Research Center for Agricultural Sciences. Filipinas. p. 590 [ Links ]
72 Tyagi A. & Mohanty A. 2000. Rice transformation for crop improvement and functional genomics. Plant Science. 158: p. 1-18 [ Links ]
73 Vasil I. 2005. The story of transgenic cereals: the challenge, the debate, and the solution - a historical perspective. in vitro Cell. Dev. Biol. Plant. 41: p. 577-583 [ Links ]
74 Vaughan D., Morishima H. & Kadowaki K. 2003. Diversity in the Oryza genus. Current Opinion in Plant Biology. 6: p. 139-146 [ Links ]
75 Villalba A. 2009. Resistance to Herbicides. Glyphosate. Exact and Naturales Sciencies. 39: p. 169-186 [ Links ]
76 Welch R. & Graham R. 2004. Breeding for micronutrients in staple food crops from a human nutrition perspective. Journal of Experimental Botany. 55 (396): p. 353-364 [ Links ]
77 WHO & FAO. 2004. Vitamin and mineral requirements in human nutrition: report of a joint FAO/WHO expert consultation. World Health Organization and Food and Agriculture Organization of the United Nations. p 17-37. Disponible en línea en: http://whqlibdoc.who.int/publications/2004/9241546123.pdf [ Links ]
78 Xu S., Li T., Deng Z., Chong K., Xue Y. & Wang T. 2008. Dynamic Proteomic Analysis Reveals a Switch between Central Carbon Metabolism and Alcoholic Fermentation in Rice Filling Grains. Plant Physiology. 148: p 908-925 [ Links ]
79 Xu Y., McCouch S. & Zhang Q. 2005. How can we use genomics to improve cereals with rice as a reference genome?. Plant Molecular Biology. 59: p. 7-26 [ Links ]
80 Ye X., Al-Babili S., Kloeti A., Zhang J., Lucca P., Beyer P. & Potrykus I. 2000. Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science. 287 (5451): p. 303-305 [ Links ]
81 Yu J., Wang J., Lin W., Li S., Li H., et al. 2005. The genomes of Oryza sativa: A history of duplications. PLoS Biol 3(2): p. 266-281 [ Links ]
