










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol vol.15 no.1 Bogotá jan./jun. 2013
ARTÍCULO DE INVESTIGACIÓN
Bacterias celulolíticas aisladas del intestino de termitas (Nasutitermes nigriceps) con características probióticas y potencial en la degradación del pasto
Título corto: Bacterias celulolíticas aisladas del intestino de termitas (Nasutitermes nigriceps)
Isolated cellulolytic bacteria in the gut termites (Nasutitermes nigriceps) with probiotic characteristics and potential pasture degradation
Cecilia Lara Mantilla1 y Roberto Carlos Acosta Pineda2.
1Química; MSc; Ph. D. Línea de Investigación en Biotecnología. Directora e investigadora GRUBIODEQ. Investigadora principal proyecto. Directora Tesis Maestría. Universidad de Córdoba. lara_mantilla_cecilia@hotmail.com; clara@correo.unicordoba.edu.co
2 Biólogo. Magister en Biotecnología. Investigadora GRUBIODEQ. Universidad de Córdoba.
Recibido: octubre 10 de 2012 Aprobado: junio 15 de 2013
Resumen
El objetivo de la presente investigación fue aislar bacterias celulolíticas del intestino de termitas (Nasutitermes nigriceps) para determinar sus propiedades probióticas in vitro y su potencial en la degradación de pasto. Las termitas fueron tratadas con detergente antibacterial, separando y macerando luego, el intestino de las mismas en agua peptonada estéril. Diluciones de esta mezcla fueron inoculadas en cajas Petri con medio Luria Bertani (LB) y Carboximetilcelulosa (CMC) al 2%, incubando a 37º C por 24 horas, para luego revelar con Rojo Congo al 1%. Las bacterias que presentaron mayores halos de degradación fueron sometidas a tinción de Gram y a pruebas probióticas de temperatura, pH, salinidad y presencia de sales biliares, así como también a pruebas de antagonismo y degradación de pasto. Los resultados revelaron la presencia de 9 bacilos celulolíticos Gram (-) de los cuales, los bacilos BTN7 y BTN8, presentaron los mejores halos de degradación, 12 y 14 mm de diámetro respectivamente, creciendo adecuadamente en las diferentes pruebas probióticas con densidades entre 106 y 108 UFC/ml; el porcentaje de degradación de materia seca fue de 39.73% y 36.10% en 48 y 72 horas respectivamente. Las pruebas bioquímicas API E (bioMérieux SA) revelaron que los bacilos BTN7 y BTN8 pertenecen al género Enterobacter sp. Los anteriores resultados abren la posibilidad de emplear, estos microorganismos como aditivos en la alimentación de rumiantes, a fin de contribuir con un mayor aprovechamiento de pastos, y otros sustratos vegetales lignocelulósicos.
Palabras clave: Probióticos, Maralfalfa (Pennisetum sp.), microorganismos ruminales, lignocelulosa, Enterobacter.
Abstract
The objective of this research was to isolate cellulolytic bacteria in the gut of termites (Nasutitermes nigriceps) to determine their probiotic properties in vitro and its potential degradation of grass. Termites were treated with antibacterial detergent, separating and then macerating the intestine of the same in sterile peptone water. Dilutions of this mixture were inoculated in Petri dishes with Luria Bertani (LB) and carboxymethylcellulose (CMC) 2%, incubated at 37 ° C for 24 hours, and then revealed with 1% Congo Red. Bacteria that had higher degradation halos were subjected to tests and Gram stain probiotic temperature, pH, salinity and presence of bile salts, as well as antagonism tests and degradation of grass. The results revealed the presence of 9 bacilli cellulolytic Gram (-) of which the bacilli BTN7 and BTN8, showed the best degradation halos, 12 and 14 mm in diameter respectively, growing well in the different tests probiotic with densities between 106 and 108 CFU / ml, the degradation percentages of dry matter was 39.73% and 36.10% within 48 and 72 hours respectively. Biochemical tests revealed that API E (bioMérieux SA) bacilli BTN8 BTN7 and belong to the genus Enterobacter sp. The above results open the possibility of using these organisms as additives in feed for ruminants, in order to contribute to a better use of pasture, and other lignocellulosic plant substrates.
Key words: Probiotics, Maralfalfa (Pennisetum sp.), ruminal microbes, lignocellulose, Enterobacter.
Introducción
Las termitas han sido consideradas tradicionalmente como plagas que destruyen la madera, pero también poseen una importante función en la naturaleza como descomponedoras de materia orgánica rica en lignocelulosa (Ramírez y Lafranco, 2001). En la región Sucreña Colombiana, una práctica tradicional entre los pequeños ganaderos es la utilización de termiteros arbóreos como aditivo para la alimentación de bovinos afirmando que su uso favorece la ganancia de peso en estos animales. Aunque no se han encontrado reportes científicos específicos relacionados con este aspecto, el efecto puede ser considerado como una consecuencia de la incorporación en el rumen de microorganismos lignocelulolíticos (Warnecke, et al., 2007) que habitan en el intestino de las termitas o en la estructura del termitero, y que pueden contribuir con el mejoramiento de la degradación de pastos y forrajes.
Existen algunas investigaciones que argumentan la presencia de bacterias con capacidad celulolítica en el intestino de las termitas y otros estudios que indican, que la actividad metabólica de esos microorganismos es similar a la realizada por las bacterias ruminales. Breznak y Switzer, citados por Ibarra (2006), plantean que las termitas digieren sustratos lignocelulósicos gracias a microorganismos simbiontes como bacterias, protozoos y hongos que habitan en sus intestinos, transformándolos en acetato como principal fuente de energía y precursor biosintético. Méndez y Equihua (2001), afirman que las termitas obreras son capaces de degradar la celulosa de los vegetales con la ayuda de protozoarios, bacterias y levaduras que viven dentro de su tracto intestinal y que facilitan las enzimas para este fin. Ohkuma (2008), refiere que las termitas inferiores tienen una porción dilatada en el intestino grueso densamente poblada por microorganismos simbiontes que contribuyen en la digestión de sustancias lignocelulósicas. Warnecke et al., (2007), llevaron a cabo un análisis de metagenómica en la comunidad de bacterias que residen en el intestino grueso de termitas Nasutitermes revelando presencia de Treponema y Fibrobacter. Sakar et al., (1988), en estudios realizados en Odontotermes, encontraron que Micrococcus luteus y M. roseus degradaron varios tipos de celulosa mediante la producción de celulasas endógenas y exógenas in vitro.
Dentro de las bacterias con capacidad celulolítica encontradas en el intestino de las termitas pueden existir algunas con propiedades probióticas las cuales pueden contribuir a mejorar la degradación de material fibroso; los microorganismos probióticos deben poseen característica importantes como son, la resistencia al ácido, pH, temperatura, sales biliares y, otros, que les permite superar las barreras ácida y biliar en el tracto digestivo de los animales, manteniéndose así en estado viable y concentraciones adecuadas (Tuomola et al., 2001; Rondón et al., 2008). Los microorganismos probioticos revisten una gran importancia por los efectos benéficos que aportan en la nutrición animal.
El objetivo del presente trabajo fue aislar bacterias celulolíticas del intestino de termitas arbóreas (Nasutitermes nigriceps), recolectadas en la Ciénaga de la Caimanera del municipio de Coveñas - Sucre, para evaluar sus características probióticas in vitro y determinar su potencial en la degradación del pasto Maralfalfa (Pennisetum sp).
Materiales y métodos
- Recolección de las termitas. Se colectaron manualmente termiteros arbóreos del género Nasutitermes ubicados en la Ciénaga de la Caimanera del municipio de Coveñas en el departamento de Sucre y se extrajeron las termitas mediante flotación en solución salina 20% p/v. Las termitas aisladas fueron lavadas con detergente antibacterial y posteriormente, se procedió a separar y macerar el intestino en agua peptonada estéril 0.1% p/v, llevando la mezcla a un tubo de ensayo y agitándola en un vortex para liberar las bacterias del intestino. Se realizaron diluciones de esta mezcla y se inocularon en cajas Petri con medio Luria-Bertani (LB) suplementado con Carboximetilcelulosa (CMC) 2% p/v (Ortiz et al., 2010); las cajas fueron incubadas a 30º C, durante 48 horas en condiciones anoxigénicas; finalizado el tiempo, se vertió sobre la superficie de la caja, una solución de Rojo Congo al 1% durante 10 minutos. Pasado ese tiempo se descartó el exceso de colorante y se lavó tres veces con solución salina; seguidamente se seleccionaron y aislaron los microorganismos que presentaron halos claros de hidrólisis sobre el fondo rojo con un diámetro no menor de 5 mm y se purificaron con repiques sucesivos (Gaitán y Pérez, 2007).
- Evaluación de las características probióticas in vitro. Las bacterias que presentaron halos con mayores dimensiones fueron sometidas a las siguientes pruebas probióticas empleando una concentración conocida de inoculo (10 6 UFC/ml): a) tolerancia a temperatura de 30º y 42º C (Ávila, et al., 2010); b) tolerancia a diferentes concentraciones de sales, biliares, 0.0, 0.05, 0.15, y 0.3% p/v (Guilliland et al., 1990) y de NaCl, 0, 2, 5 y 10% p/v (Rondón et al., 2008); d) evaluación a diferentes valores de pH, 5, 6 y 7 (Ortiz y Reuto, 2007); e) pruebas de antagonismo empleando bacterias ruminales y E.coli para observar la formación de halos de inhibición de crecimiento en placa (Gaitán y Pérez, 2007). En todos los casos de evaluó la población bacteriana por conteo en placa en UFC/ml.
- Prueba de degradación de sustratos vegetales lignocelulósicos. El experimento se realizó in vitro utilizando hojas de pasto Maralfalfa (Pennisetum sp), las cuales fueron lavadas, secadas y molidas hasta conseguir trozos de 1 a 2 mm de longitud. Seguidamente se esterilizó el pasto en autoclave y se vertió 1 g de éste, en varios tubos de ensayo con medio líquido LB; los tubos fueron inoculados con las bacterias seleccionadas en una concentración de 106 UFC y luego incubados por 24, 48 y 72 horas; pasado el tiempo se extrajo el sobrenadante y se determinó la concentración de azucares reductores mediante la prueba del ácido 3,5-dinitrosalicílico (DNS) utilizando un Espectrofotómetro y una longitud onda de 540 nm; se utilizó una muestra control en donde se determinó la concentración inicial de azúcares reductores. Todos los ensayos se llevaron acabo por triplicado (Gaitán y Pérez, 2007; Sosa et al., 2006).
Para establecer la digestibilidad del pasto por parte de las bacterias seleccionadas se extrajo el medio líquido de cada tubo de ensayo con una pipeta y el precipitado se llevó a un horno a 60ºC por 1 hora. Pasado este tiempo se pesó el material vegetal residual y se restó a la cantidad inicial de pasto en cada tubo de ensayo para calcular el porcentaje de digestibilidad de acuerdo a la siguiente ecuación (Sosa et al., 2006):
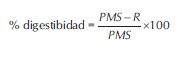
PMS: Peso inicial de la materia seca.
R: Residuo o peso final de la materia seca. - Identificación de los microorganismos. Las bacterias que presentaron los mejores resultados en las pruebas probióticas y degradación de pasto fueron sometidas a pruebas bioquímicas rutinarias y a través del Kit comercial API E (bioMérieux SA) para identificar el género y posible especie.
- Análisis estadístico. Los ensayos fueron organizados en un diseño completamente al azar, los datos obtenidos se sistematizaron y analizaron mediante una ANAVA y en el caso de la existencia de diferencias significativas se aplicó una prueba de comparaciones múltiples a través de la Diferencia Mínima Significativa (DMS) en el programa SPSS.
Resultados y discusión
Las termitas halladas y recolectadas en los árboles de mangle del estuario de la Ciénaga de la Caimanera en el municipio de Coveñas, departamento de Sucre pertenecen al género Nasutitermes especie nigriceps (Haldeman, 1853; Abadía y Arcila, 2009), el cual se agrupa dentro de la familia Termitidae (Matsui, et al., 2009). Esta familia de termitas tiene una dieta basada principalmente en hojarasca, madera y pastos (Fernández, et al., 2008), para lo cual, poseen bacterias intestinales con actividad celulolítica que facilitan la digestión de estos materiales. En este sentido, para el caso de las termitas Nasutitermes, según Wenzel, et al., (2002), se ha demostrado que las bacterias que habitan en sus intestinos contribuyen con la digestión de la celulosa.
- Características de las bacterias celulolíticas aisladas. El revelado con Rojo Congo permitió aislar 9 microorganismos con actividad celulolíticas de un total de 30 bacterias; la observación microscópica con tinción de Gram permitió observar bacilos Gram (-) de diferente morfología, los cuales fueron codificados como BTN1, BTN2, BTN3, BTN4, BTN5, BTN6, BTN7, BTN8 y BTN9. Asimismo, se encontró que los bacilos BTN2, BTN6, BTN7 y BTN8 presentaron los mayores halos de hidrólisis de CMC, 7, 10, 14 y 12 cm respectivamente, siendo un indicador de la actividad celulolítica de estas bacterias (tabla 1). Por lo cual, fueron seleccionadas para el desarrollo de las pruebas probióticas y de degradación de pasto.
La presencia de los halos en el medio sólido con CMC es un indicador de que las bacterias poseen enzimas que digieren celulosa. Los valores obtenidos en la prueba son similares a los reportados por Ortíz, et al., (2010), quienes obtuvieron halos de degradación entre 3 y 15 mm de diámetro con cepas de bacterias aisladas de nejayote de maíz (mezcla de agua, cal y residuos de cascarilla y endospermo de maíz). - Características probióticas evaluadas. Dentro de los criterios de selección para considerar microorganismos con potencial probiótico se encuentran, la capacidad de sobrevivir en el tracto gastrointestinal, resistir a las secreciones digestivas (gástricas, bilis, etc.) y poseer actividad antimicrobiana contra patógenos (Guillot, 2003). Los resultados obtenidos se analizan a continuación.
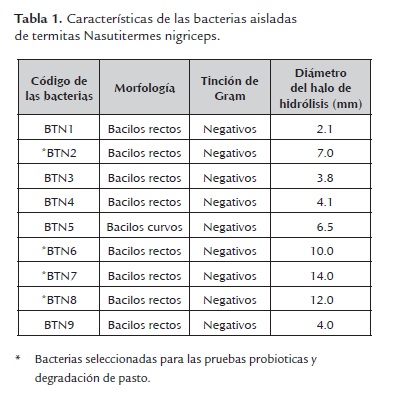
a) Tolerancia a cambios de temperatura. Los bacilos BTN2, BTN7 Y BTN8 mantuvieron su crecimiento en concentraciones de 108 UFC/ml después de 24 horas de incubación a temperatura de 30º y 42º C, indicando con ello que estas bacterias no son susceptibles a las variaciones de temperatura en este rango. Sin embargo, el bacilo BTN6 presentó un aumento significativo en su crecimiento a 30º C, pasando de una concentración de 107 a 108 UFC/ml. De igual manera, se observó que esta bacteria no fue capaz de crecer a 42º C, lo cual revela que es muy susceptible a las variaciones de temperatura (tabla 2).
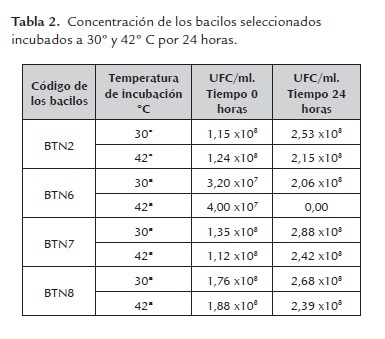
Estadísticamente se observó, que el P-valor (0.067) fue mayor que alfa (0.05); la prueba DMS para comparaciones múltiples permitió establecer que el crecimiento del bacilo BTN6 fue estadísticamente diferente de los bacilos BTN7 y BTN8; no se observó diferencias significativas entre los bacilos BTN2, BTN7 y BTN8, los cuales demostraron capacidad de crecer en cantidades adecuadas (10 8 UFC/ml), a temperaturas relativamente altas como las encontradas en el rumen (38° y 42° C) (Van Lier, 2009). Este dato es importante si se tiene en cuenta que las concentraciones más habituales de un microorganismo probiótico oscilan entre 108 y 1010 UFC/ml, (Acedo y Rico 1998).
b) Tolerancia a sales biliares. Se observó que los bacilos BTN2, BTN7 y BTN8 presentaron incrementos significativos en la concentración de UFC/ml después de 24 horas de incubación, en presencia de sales biliares en rangos de 0.05% a 0.30%, pasando, en todos casos, de una densidad de 107 a 108 UFC/ml. El bacilo BTN6 no tuvo incrementos poblacionales significativos en ninguna de las concentraciones (tabla 3). El crecimiento de los bacilos no se afectó por las diferentes concentraciones de las sales biliares; éste resultado es deseable en los microorganismos candidatos a probióticos puesto que aseguran su paso a través del tracto gastrointestinal del animal hospedero pudiendo desarrollar sus actividades metabólicas sin verse inhibidas.
Estudios realizados por Ávila, et al., (2010), revelaron buenos crecimientos de cepas de Lactobacillus aislados del tracto intestinal de animales de granja en presencia de sales biliares al 0.15%; Campos et al., (2009), reportaron crecimientos en placa de bacterias lácticas de 1.7x107 UFC/ml en medios con 0.30% de sales biliares, los cuales son inferiores a los obtenidos en esta prueba con los bacilos nativos BTN2, BTN6 y BTN8, pero similares a los obtenidos con el bacilo BTN7. Sin embargo, en todos estos ensayos se alcanzaron densidades poblacionales de 107 y 108 UFC/ml, encontrándose dentro de los valores habituales establecidos para microorganismos probióticos (Acedo y Rico, 1998).
Las variaciones observadas revelan un comportamiento inversamente proporcional entre el crecimiento de las bacterias y las concentraciones de sales biliares empleadas. El análisis de varianza para esta prueba demostró la existencia de diferencias significativas en el ensayo, debido a que el P-valor (0.017) fue menor que el alfa de 0.05; la prueba de DMS para comparaciones múltiples permitió establecer que existen diferencias entre los crecimientos obtenidos en ausencia y presencia de sales biliares para cada uno de los bacilos estudiados,. Asimismo, se determinó que no existen diferencias significativas entre los crecimientos obtenidos en concentraciones de sales biliares de 0.05%, 0.15% y 0.03%, para cada uno de los bacilos.
c) Tolerancia a NaCl. En esta prueba se observó que el bacilo BTN2 presentó un incremento en la magnitud de su densidad poblacional inicial después de 24 horas de incubación en presencia de NaCl al 2%, pasando de 107 a 108 UFC/ml, mientras que los bacilos BTN6, BTN7 y BTN8 se mantuvieron. De igual forma, se observó que para concentraciones de NaCl al 5%, los bacilos BTN2, BTN7 y BTN8 mantuvieron sus magnitudes de crecimiento, entre tanto que el bacilo BTN6 tuvo una disminución sustancial pasando de 108 a 106 UFC/ml; los datos revelan que todos los bacilos en estudio presentaron disminución de la magnitud de su densidad poblacional inicial en presencia de NaCl al 10% (tabla 4). Asimismo, se observó que el crecimiento de las bacterias seleccionadas es inversamente proporcional a la concentración de NaCl y el bacilo BTN6 es el más susceptible a los incrementos de salinidad, mientras que las bacterias BTN2, BTN7 y BTN8 son las tolerantes frente a este factor. A pesar de estas diferencias observables, el análisis de varianza para esta prueba revela que no hay diferencias significativas entre los crecimientos de los bacilos estudiados, en cada una de las concentraciones de NaCl ensayadas, ya que el P-valor (0.347) fue mayor que el alfa del 0.05.
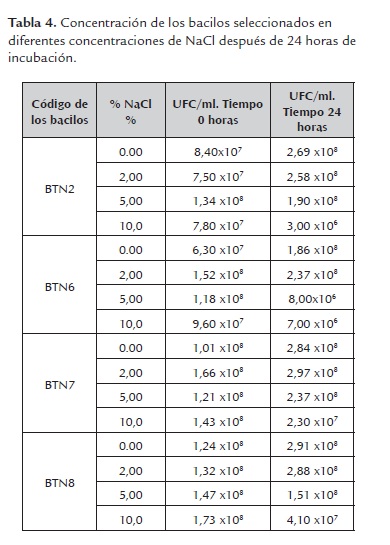
En consecuencia, teniendo en cuenta los resultados obtenidos en esta prueba, se puede decir que las bacterias seleccionadas en este estudio tienen capacidad para adaptarse fácilmente a condiciones de salinidad relativamente altas, ya que demostraron que pueden crecer en densidades de 106 hasta 108 en concentraciones de NaCl entre 2% y 10% con una osmolaridad de 684 mOsm y 3400 mOsm respectivamente. Al respecto, según Van Lier y Regueiro, (2008), la osmolaridad del líquido ruminal oscila entre 260 y 340 mOsm, pudiendo llegar a 400 mOsm con el consumo de concentrado. Por lo que este factor no afectaría significativamente a los bacilos estudiados a nivel de este órgano.
d) Tolerancia a pH. Se observó que ninguna de las bacterias estudiadas creció a pH 4 después de 24 horas de incubación; el bacilo BTN6 a diferencia de los bacilos BTN2, BTN7 y BTN8, presentó un incremento en la magnitud de su densidad poblacional pasando de 107 a 108 UFC/ml en medios con pH 5 y 7 (tabla 5). Marrero, et al., (2010), reportó crecimientos de bacterias celulolíticas de 1.7x108 UFC/ml y 7.87x108 UFC/ml a pH 6.6 a nivel del rumen, siendo muy similar a los resultados obtenidos en esta prueba para cada una de los bacilos estudiados. De igual forma, Hernández y Cobos, (2001) reportaron crecimientos de bacterias celulolíticas extraídas del colon distal de conejos de 4x102 UFC/ml a pH 6.3 y del apéndice cecal de 9x107 a pH 7.3, inferiores en la mayoría de los casos a los obtenidos en este ensayo.
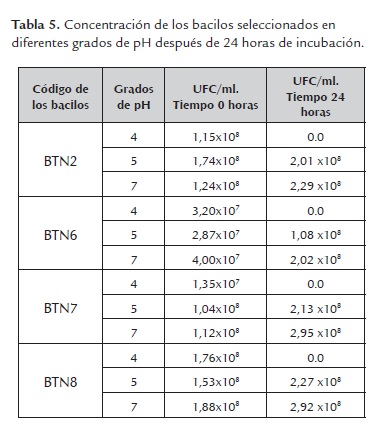
El análisis de varianza para esta prueba reveló que no existen diferencias significativas entre los crecimientos obtenidos de los bacilos seleccionados en los diferentes grados de pH, debido a que el P-valor (0.380) fue mayor que el alfa del 0.05.
Con respecto a esta prueba, según Calsamiglia y Ferret, (2002), el pH normal óptimo en el rumen oscila entre 6.2 y 7.0, por lo que se puede decir que las bacterias estudiadas pueden crecer en cantidades adecuadas a nivel del rumen sin afectarse significativamente por variaciones entre pH 5 y pH 7. Van Lier y Regueiro, (2008), refiere que el pH ruminal puede variar entre 5.8 y 7.0, pero puede disminuir aun más luego de la ingesta de concentrados debido a la rápida fermentación producida que genera un medio ácido; también, plantean que las bacterias celulolíticas del rumen se inhiben a pH menor de 6.0. No obstante, las bacterias celulolíticas empleadas en este estudio crecieron a pH 5 en cantidades adecuadas (108 UFC/ml), lo que indica que pueden adaptarse fácilmente al pH ruminal.
e) Prueba de antagonismo. En este ensayo se observó que las colonias de bacterias de las termitas no inhibieron significativamente el crecimiento de las bacterias ruminales utilizadas, ya que, en el caso de las bacterias BTN2 y BTN7, presentaron halos alrededor de los discos de papel filtro que no superaron los 2 mm de ancho, mientras que las bacterias BTN6 y BTN8 no formaron halos de inhibición. En presencia de bacterias E. Coli aisladas del líquido ruminal se pudo observar que las bacterias BTN2 y BTN8 formaron halos de 2 mm de ancho alrededor de los discos de papel filtro, en comparación con las bacterias BTN6 y BTN7 que formaron halos de inhibición de E. coli de 3 mm cada una, siendo diferencias poco relevantes. Gaitán y Pérez, (2007), reportaron tamaños de halos de inhibición muy superiores (5mm y 7.5 mm) a los de este ensayo, formados por interacción entre bacterias celulolíticas aisladas de compost generados en cultivos de crisantemo. Zamudio y Zavaleta, (2003) reportaron halos de inhibición de E.coli y Listeria monocytogenes entre 2 y 8 mm de diámetro, superiores a los hallados en este ensayo. Mientras tanto, Barrera y Charry, (2008), obtuvieron cepas aisladas de compostaje de residuos de lechuga que no presentaron efectos antagónicos.
Este aspecto se considera uno de los más importantes, debido a que se requiere que las bacterias aisladas del abdomen de Nasutitermes nigriceps al incorporarse al rumen no afecten el desarrollo de la microbiota normal que realiza funciones importantes en el metabolismo de los alimentos que consumen los bovinos. Sin embargo, el control de las poblaciones de Coliformes en el animal es deseable ya que estas pueden causar enfermedades a nivel intestinal. Por lo tanto, las bacterias en estudio cumplen con este requisito pudiendo ser incorporadas al rumen sin afectar negativamente su ecosistema.
- Prueba de degradación in vitro de sustratos vegetales lignocelulósicos. En este ensayo se observó que durante las primeras 48 horas las bacterias BTN2, BTN6 y BTN8 produjeron burbujas de gas en los medios, siendo más evidente este fenómeno a las 24 horas y cesando por completo a las 72 horas; la bacteria BTN7 no produjo burbujeo durante el tiempo de observación del experimento. En relación con lo anterior, se han reportado bacterias productoras de gas metano e hidrógeno (Ibarra, 2006) aisladas del intestino de termitas, por lo que es posible que las burbujas observadas correspondan a algunos de estos gases, que se producen principalmente durante la fermentación de la glucosa (Van Lier y Regueiro, 2008).
En cuanto a la producción de azúcares reductores, se observó que las bacterias BTN6 y BTN8 presentaron una tendencia similar, notándose un incremento en la concentración de estos azúcares en las primeras 48 horas, siendo más eficiente la bacteria BTN8, con un descenso posterior en dichas concentraciones a las 72 horas de incubación. Entre tanto, las bacterias BTN2 y BTN7 presentaron un comportamiento similar con aumento progresivo en los valores de la concentración de azúcares reductores durante las 72 horas de incubación, destacándose la bacteria BTN7 que obtuvo la mayor concentración de estos azúcares en este periodo de tiempo (tabla 6). La eficiencia las bacterias para producir azúcares reductores a partir del pasto se debe principalmente al ataque de enzimas microbianas específicas sobre la celulosa y la hemicelulosa que conforman la fibra vegetal (Lynd et al., 2002). Por lo que se puede decir que las bacterias BTN7 y BTN8 presentaron la mayor actividad celulolítica en el ensayo.
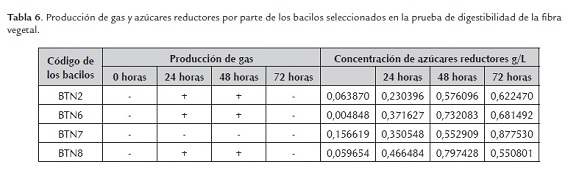
Para el propósito de la presente investigación la bacteria BTN8 presentó los mejores resultados en esta prueba, ya que se requiere que la bacteria al momento de incorporarse al rumen degrade la mayor cantidad fibra en el menor tiempo posible, liberando azúcares reductores que posteriormente sean convertidos en ácidos grasos volátiles (AGV) fundamentales para el rumiante. Si bien la bacteria BTN7 obtuvo el mejor resultado a las 72 horas de incubación, no se observó que utilizara estos carbohidratos, ya que la concentración de los mismos se mantuvo en aumento.
No obstante, a pesar de las variaciones observadas con respecto a la producción de azúcares reductores por parte de los bacilos seleccionados, el análisis de varianza reveló que no existen diferencias significativas en este aspecto, ya que se obtuvo un P-valor de 0.956 mayor que el alfa de 0.05.
En cuanto a la digestibilidad del pasto se observó que las bacterias BTN6 y BTN8 presentaron un comportamiento similar, incrementando el porcentaje de digestibilidad del pasto entre las 24 y 48 horas, siendo la bacteria BTN8 la más eficiente (36.10%). Asimismo se observó que estas bacterias disminuyeron su actividad degradadora a las 72 horas de incubación. En contraste, las bacterias BTN2 y BTN7 tuvieron una tendencia parecida incrementando progresivamente el porcentaje de digestibilidad del pasto durante las 72 horas de incubación, destacándose la bacteria BTN7 que presentó el mayor porcentaje de digestibilidad durante este periodo de tiempo (39.73%).
Los datos obtenidos en este experimento son superiores a los reportados por Grilli et al., (2011) quienes obtuvieron porcentajes de digestibilidad de celulosa en alfalfa (Medicago sativa) de 19,10% ± 4,721% con una cepa de Fibrobacter succionógenes, pero inferiores a los reportados por Clavero y Razz, (2009), donde la digestibilidad in vitro de Pennisetum sp. alcanzó el 52.1% y 62.45% utilizando bacterias ruminales. Igualmente fueron inferiores a los reportados por Heredia y Paladines, (2007), en los cuales el porcentaje de digestibilidad in situ de este pasto fue de 76.7% en promedio. Asimismo, fueron inferiores a los reportados por Soto, (2007) morera (Morus alba) de 64.4% y en madero negro (Gliricidia sepium) de 54%, en condiciones in vitro utilizando líquido ruminal. De igual forma, estuvieron dentro de los rangos obtenidos por Kato et al., (1998), quienes encontraron que bacterias aisladas del intestino de termitas Nasutitermes takasagoensis fueron capaces de degradar, en condiciones in vitro, compuestos lignocelulósicos presentes en el medio de cultivo con porcentajes variables de 28%, 60% y 95%. Cabe resaltar, que en las pruebas de digestibilidad de fibra donde se emplea líquido ruminal la degradación del material vegetal es mas compleja y es producida no solo por bacterias, sino, también, por protozoarios y hongos que habitan en el rumen.
El análisis de varianza para esta prueba reveló que no existen diferencias significativas entre las bacterias con respecto a los porcentajes de digestibilidad del pasto Maralfalfa (Pennisetum sp.) empleado en el ensayo, ya que el P-valor (0.862) fue mayor que el alfa de 0.05.
Después de analizar las pruebas realizadas a las diferentes bacterias aisladas del intestino de termitas Nasutitermes nigriceps, se puede observar que la bacteria BTN7, presentó los mejores resultados, seguida de la bacteria BTN8. Las pruebas bioquímicas API 20E permitieron identificar que estas bacterias pertenecen al género Enterobacter aerogenes. Este tipo de bacteria ya había sido reportado por Ramin et al., (2008) en el intestino de termitas Coptotermes curvignathus demostrando que eran microorganismos anaerobios facultativos Gram (-), mientras que Kuhnigk et al., (1994) aisló bacterias Enterobacter aerogenes con capacidad para degradar monómeros de lignina de termitas Mastotermes darwiniensis y Nasutitermes nigriceps. Asimismo, Dechamps, citado por Ramin, et al., (2008) reveló que bacterias el género Enterobacter tienen capacidad para asimilar compuestos fenólicos.
Los resultados sugieren que las bacterias BTN7 y BTN8 tienen capacidad para crecer adecuadamente en condiciones relativamente difíciles de temperatura, salinidad, pH y presencia de sales biliares, con buen porcentaje de digestibilidad de fibra vegetal y ausencia de antagonismo frente a microorganismos ruminales, lo cual indica que la acción de las bacterias celulolíticas que habitan en el intestino de las termitas (Nasutitermes nigriceps) puede ser una de las causas que favorece la ganancia de peso de los bovinos alimentados con termiteros en la región Sucreña. Siendo, por lo tanto, candidatas para utilizarse como probióticos en la alimentación de rumiantes.
Teniendo en cuenta la poca información que existe con respecto al efecto positivo producido por el consumo de termitas arbóreas sobre la ganancia en peso de bovinos sucreños y, con el ánimo de contribuir en el conocimiento en éste aspecto, se llevo a cabo el presente trabajo; sin embargo, es necesario continuar con el desarrollo de más investigaciones al respecto.
Conclusión
Los resultados de la presente investigación permiten concluir lo siguiente:
- Las termitas Nasutitermes nigriceps recolectadas en la Ciénaga de la Caimanera en el municipio de Coveñas, contienen en el interior de su intestino bacilos celulolíticos Gram (-) identificados como Enterobacter aerógenes.
- Los bacilos Enterobacter aerógenes son capaces de sobrevivir en condiciones ambientales relativamente difíciles manteniendo concentraciones adecuadas, sin inhibir el crecimiento de bacterias ruminales, pero afectando significativamente el crecimiento de bacterias E. Coli.
- Este tipo de bacterias presentan una actividad celulolítica que les permite degradar la fibra vegetal del pasto a nivel in vitro en porcentajes significativos.
- Las capacidades de supervivencia y digestión de la fibra vegetal que poseen los bacilos Enterobacter aerogenes, sugiere que pueden adaptarse a las condiciones ambientales del rumen y favorecer la digestibilidad del pasto a nivel de este órgano, pudiendo repercutir en la nutrición de bovinos alimentados con termiteros.
Agradecimientos
Los autores expresan sus agradecimientos a la Universidad de Córdoba y a la Universidad de Sucre, por su colaboración.
Referencias bibliográficas
1 Abadía, J. y Arcila, A. 2009. Termitas en cultivos de limón en los departamentos del Atlántico y Magdalena, Colombia. Boletín del museo de entomología de la Universidad del Valle. 10(2): 36-46. [ Links ]
2 Acedo, J. y Rico, G. 1998. Utilización de aditivos en piensos para rumiantes: minerales forma orgánica, levaduras, enzimas, ionóforos y otros. Disponible en: www.produccion-animal.com.ar. [ Links ]
3 Ávila, J., Ávila, M., Tovar, B., Brizuela, M., Perazzo, Y. y Hernández, H. 2010. Capacidad probiótica de cepas del género Lactobacillus extraídas del tracto intestinal de animales de granja. Revista Científica, 20 (2): 161-169. [ Links ]
4 Barrera, L. y Charry, N. 2008. Producción y evaluación de un inoculante microbiano con capacidad amilolítica a partir de un proceso de compostaje de residuos de lechuga. Trabajo de grado. Pontificia Universidad Javeriana. p. 43-44. [ Links ]
5 Calsamiglia, S. y Ferret, A. 2002. Fisiología ruminal relacionada con la patología digestiva: acidosis y meteorismo. XVIII Curso de Especializacion FEDNA, Barcelona, España. Disponible en: http://www1.etsia.upm.es/fedna/capitulos/2002CAP_VI.pdf. [ Links ]
6 Clavero, T. y Razz, R. 2009. Valor nutritivo del pasto maralfalfa (Pennisetum purpureum y Pennisetum glaucum) en condiciones de defoliación. Revista Facultad de Agronomía (Luz). 26:78-87. [ Links ]
7 Fernández, M., De Lima, H. y de Aquino, A. 2008. Asociaciones entre termitas y microorganismos. Documentos 254. Seropédica Embrapa Agrobiología. P. 1-20. [ Links ]
8 Gaitan, D. y Pérez, L. 2007. Aislamiento y evaluación de microorganismos celulolíticas a partir de residuos vegetales frescos y en composta generado en un cultivo de crisantemo. Trabajo de grado. Pontificia Universidad Javeriana. P. 29-30. [ Links ]
9 Grilli D, Paez S, Egea V, Cerón M, Cobos E, Allegretti L, y Arenas N. 2011 Determinación in vitro de la Digestibilidad de la Celulosa contenida en pasturas autóctonas por una Cepa de Fibrobacter succinogenes aislada de Cabras Biotipo Criollo. Revista bioanálisis, P. 35. [ Links ]
10 Guilliland, S. y Walker, D. 1990. Factos to consider when selectin a culture of Lactobacillus acidophilus as a dietary adjunct to produce a hipocolesterolemic effect in humans. Dairy science. 73: 905-911 [ Links ]
11 Guillot, J.F. 2003. Probiotic feed additives. Journal of Veterinary Pharmacology and Therapeutics 26 (Suppl. 1): 52-55. [ Links ]
12 Heredia, N. y Paladines, O. 2007. Respuesta del pasto maralfalfa (Pennisetum violaceum) a la fertilización nitrogenada con dos distancias de siembra. Cayambe, Pichincha. Revista RUMIPAMBA XXI(1) : 2-10 [ Links ]
13 Hernández, D. y Cobos, M. 2001. Digestibilidad in vitro, población de bacterias celulolíticas y totales del apéndice cecal, ciego y colon del conejo. Revista técnica pecuaria en México. 39( 3) : 229-236. [ Links ]
14 Ibarra, J. 2006. Aislamiento de bacterias anaerobias hidrogenotróficas en el tracto gastrointestinal del avestruz (Struthio camelos). Tesis de maestría, Universidad de Colima. México. P. 22-24. [ Links ]
15 Kato, K., Kozaki, S., Sakuranaga, M. 1998. Degradation of Lignin Compounds by Bacteria from Termite Guts Source: Biotechnology Letters, 20( 5): 459-462. [ Links ]
16 Kuhnigk, T., Borst, A., Ritter, P., Kampfer, A., Graf, A., Hertel, H and Koning, H. 1994. Degradation of lignin monomers by the hindgut flora of xylophagous termites. Journal Systematic and Applied Microbiology. 17: 76-85. [ Links ]
17 Lynd, L., Weimer, P., Van Zyl, W. y Pretorius, I. 2002. Microbial Cellulose Utilization: Fundamentals and Biotechnology. Microbiology and Molecular Biology Reviews 3: 506-577. [ Links ]
18 Marrero, Y., Martín, E., Rodríguez, D. y Galindo, J. 2010. Efecto de la inclusión de fracciones del cultivo de Saccharomyces cerevisiae en la fermentación ruminal in vitro de pasto estrella (Cynodon nlemfuensis). Revista Cubana de Ciencia Agrícola, 44( 2) : 161-168. [ Links ]
19 Matsui T., Tokuda G. y Shinzato, N. 2009. Termites as Functional Gene Resources. Recent Patents on Biotechnology. 3( 1) : 12. [ Links ]
20 Méndez, J y Equihua, A. 2001. Diversidad y Manejo de los Termes de México (Hexápoda, Isóptera). Acta Zoológica Mexicana. Número especial 1:173-187. [ Links ]
21 Ohkuma, M. 2008. Symbioses of flagellates and prokaryotes in the gut of lower termites. [ Links ]
22 Ortiz, A y Reuto, J. 2007. Evaluación de la capacidad probiótica in vitro de una cepa nativa de Saccharomyces cereviciae. Trabajo de grado. Pontificia Universidad Javeriana. [ Links ]
23 Ortíz, M., Muñoz, L., Ballinas, L., Calvillo, C., Rascón, Q. 2010. Caracterización de bacterias degradadoras de celulosa. Memorias del VII congreso del noreste y III nacional de ciencias alimentarias y biotecnología. [ Links ]
24 Ramin, M., Alimon, A., Abdullah, N., Panandam, J y Sijam, K. 2008. Isolation and identification of three species of bacteria from the termite Coptotermes curvignathus (Holmgren) present in tre vicinity of University Putra Malaysia. Research Journal of Microbiology. 3 ( 4) : 288-292. [ Links ]
25 Ramírez, J.C. Y Lanfranco, D. 2001. Descripción de la biología, daño y control de las termitas: especies existentes en Chile. Bosque. 22 ( 2) : 77-84. [ Links ]
26 Rondón, A., Samaniego, L., Bocourt, R., Rodríguez, S., Milian, G., Ranilla, M., Laurencio, M y Pérez, M. 2008. Aislamiento, identificación y caracterización parcial de las propiedades probióticas de cepas de Lactobacillus sp. Procedentes del tracto gastrointestinal de pollos de ceba. Revista Ciencia y Tecnología alimentaria, 6(1) : 56-63. [ Links ]
27 Sarkar, A., Varma, A. y Sarkar, A. 1988. Influence of cellulolytic organisms associated with a termite, Odontotermes obesus, on carbon mobility in a semiarid ecosistema. Arid Land Research and Management. 2 ( 2) : 75 - 84. [ Links ]
28 Sosa, D., Larco, C., Falconí, R., Toledo, D., y Suárez, G. 2006. Digestibilidad de maralfalfa (Pennisetum sp.) en cabras. Boletín Técnico 5, Serie Zoológica Volumen 2. Sangolquí - Ecuador. P. 72. [ Links ]
29 Van Lier y Regueiro, 2008. Digestión en reticulo-rumen. Curso de anatomía y fisiología animal. Facultad de agronomía universidad de la república. Montevideo, Uruguay. P. 8. [ Links ]
30 Van Lier, 2009. Ecosistema ruminal. Disponible en http://cursoafa2009.webs.com/Ecosistema-Reticulo-Ruminal-2009.pdf. [ Links ]
31 Warnecke, F., Luginbühl, P., Ivanova, N., Ghassemian, M., Richardson, T., Stege, J., Cayouette, M., Mchardy, A., Djordjevic, G., Aboushadi, N., Sorek, R., Tringe, S., Podar, M., Garcia, H., Kunin, V., Dalevi, D., Madejska, J., Kirton, E., Platt, D., Szeto, E. Salamov, A., Barry, K., Mikhailova, N., Kyrpidesk, N., Matson, E., Ottesen, E., Zhang, X., Hernández, M., Murillo, C., Acosta, L., Rigoutsos, I., Tamayo, G., Green, B., Chang, C., Rubin, E., Mathur, E., Robertson, D., HUGENHOLTZ, P. Y LEADBETTER, J. 2007. Metagenomic and functional analysis of hindgut microbiota of a wood-feeding higher termite. Revista nature. Vol. 450. [ Links ]
32 Wenzel, M., I. Schonig, M., Berchtold, P., Kampfer y Konig, H.o 2002. Aerobic and facultatively anaerobic cellulolytic bacteria from the gut of the termite Zootermopsis angusticollis. Journal of Applied Microbiology. 92 :32-40. [ Links ]
33 Zamudio, K y Zavaleta, A. 2003. Estudio del potencial probiótico de Lactobacilos aislados de fuentes naturales. Revista Ciencia e Investigación. 6 ( 1) : 30-35. [ Links ]
