










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Colombiana de Biotecnología
versão impressa ISSN 0123-3475
Rev. colomb. biotecnol vol.16 no.1 Bogotá jan./jun. 2014
https://doi.org/10.15446/rev.colomb.biote.v16n1.44226
http://dx.doi.org/10.15446/rev.colomb.biote.v16n1.44226
ARTÍCULO DE INVESTIGACIÓN
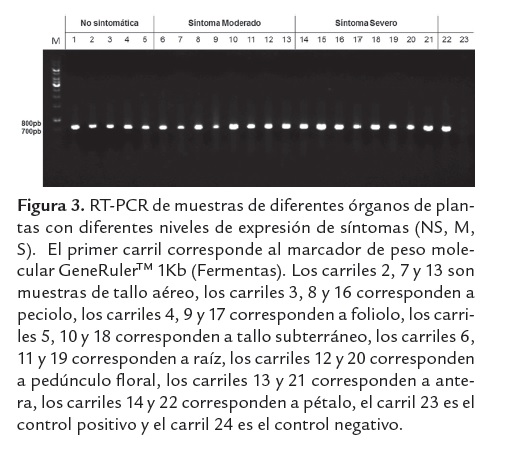
Detección del virus del amarillamiento de las nervaduras de la hoja de la papa en diferentes órganos de Solanum tuberosum grupo Phureja cv Criolla Colombia utilizando RT-PCR convencional y en tiempo real
Potato yellow vein virus detection in different organs of Solanum tuberosum Phureja group cv Criolla Colombia by conventional and real time qRT-PCR
Titulo corto: Detección del virus PYVVV en diferentes órganos de Solanum tuberosum grupo Phureja
Hernández-Guzmán Anngie Katherine1 y Guzmán- Barney M. Mónica2.
1 Bióloga, MSc. Laboratorio de Virus Vegetales, Instituto de Biotecnología (IBUN), Universidad Nacional de Colombia, Sede Bogotá, Colombia.
2 Bióloga, MSc., PhD. Laboratorio de Virus Vegetales, Instituto de Biotecnología (IBUN), Universidad Nacional de Colombia, Sede Bogotá, Colombia. E-mail: mmguzmanb@unal.edu.co.
Recibido: agosto 10 de 2013 Aprobado: mayo 2 de 2014
Resumen
La producción del cultivo de papa en Colombia se puede afectar por infección con diferentes patógenos virales, entre ellos, el Potato yellow vein virus (PYVV) que puede reducir la producción entre el 30 % y 50%. PYVV se ha diagnosticado molecularmente usando RT-PCR convencional en hojas sintomáticas y no sintomáticas. Sin embargo, no hay reportes sobre la detección y distribución viral en diferentes órganos infectados por PYVV en las plantas que expresan síntomas y sin síntomas. El objetivo de esta investigación, fue detectar a PYVV por RT-PCR convencional con cebadores específicos y por qRT-PCR (tiempo real) utilizando Sondas TaqMan® y analizar la distribución viral en plantas de S. tuberosum grupo Phureja cv. Criolla Colombia (papa criolla). Se logró la detección del virus en todos los órganos analizados (foliolo, peciolo, tallo aéreo y subterráneo, pedúnculo floral, pétalo y antera) mediante ambas técnicas, sin embargo, qRT-PCR fue 100 veces más sensible que la técnica convencional. Adicionalmente, se realizó la cuantificación absoluta del gen de la proteína mayor de la cápside de PYVV (CP). Los resultados indican que cuando la planta no expresa síntomas (NS), hay una distribución homogénea del virus, con un promedio del número de copias del gen CP de 4.09×107±2.35×107; mientras que en plantas sintomáticas el título viral es mayor (6.82×108±1.74×108) y la distribución heterogénea en los órganos, con mayor acumulación en órganos de la zona aérea. Este es el primer informe sobre la detección de PYVV en diferentes órganos de papa por medio de tiempo real, incluyendo las anteras y pedúnculo floral. La información debe ser de utilidad para el diagnóstico de PYVV y para adelantar estudios sobre la biología del virus y la relación con el huésped y el vector. La información suministrada debe ser valiosa para agricultores y fitomejoradores, además para programas de indexado de plantas contra PYVV y en la certificación de semilla.
Palabras clave: Sondas TaqMan®, PYVV, Distribución, Crinivirus, Cuantificación absoluta.
Abstract
Potato yield in Colombia could be affected by the infection with different viral pathogens, among which, Potato vein yellow virus (PYVV) could reduce potato production by 30% to 50%. PYVV has been diagnosed molecularly in symptomatic and symptomless leaves samples by conventional RT-PCR. However, the PYVV detection and distribution in different organs of symptomatic and symptomless plants have not been reported until now. The aim of this research was to detect and analyze PYVV distribution in different organs of infected S. tuberosum group Phureja cv. Criolla Colombia (papa criolla) plants using conventional and real time qRT-PCR usindTaqMan® probes. It was achieved to detect the virus in all analyzed organs (leaflets, petiole, peduncle, anther, petals, aerial and underground stem) by both techniques; however, qRT-PCR was 100 times more sensitive than the conventional technique. Additionally, the absolute quantification of coat major protein gene (CP) was determined. The results shown that in non symptomatic plants (NS), PYVV was distributed homogenously with an average CP gene copy number of 4.09 × 107 ± 2.35 × 107, while in symptomatic moderate and severe plants (M) or (S) the viral load was greater (6.82×108±1.74×108) with an heterogeneous distribution regarding the organ and with greater accumulation in the aerial organs. The results presented in this study will be important for PYVV detection and further studies on the virus biology, host and vector relations. The information should be useful to farmers, breeders, indexing and seed certification programs.
Key words: TaqMan® probes, PYVV, distribution, Crinivirus, absolute quantification.
Introducción
PYVV es un virus re-emergente con genoma tripartita ARNss+ con un tamaño de aproximadamente 17kb (Livieratos et al., 2004). Sus partículas virales son de morfología filamentosa que se limitan al floema (Salazar et al., 2000). Pertenece a la familia Closteroviridae, género Crinivirus (Martelli et al., 2002). PYVV es el agente causal de la enfermedad del amarillamiento de las nervaduras de la papa (Potato Yellow Yein Disease, PYVD) (Salazar et al., 2000) y se transmite por medio de la mosca blanca Trialeurodes vaporariorum, Westwood (Hemiptera: Aleyrodidae), de forma semipersistente, también presenta transmisión vegetativa por medio del tubérculo-semilla (Salazar et al., 2000).
PYVD fue reportada por primera vez en Colombia por Alba (1950) en Antioquia. Actualmente, se ha dispersado a países vecinos como Venezuela, Ecuador y Perú debido al incremento de las poblaciones del vector y del transporte indiscriminado de germoplasma (Salazar et al., 2000). PYVV afecta la producción de papa hasta un 50% en Solanum tuberosum grupo Andígena cv Diacol Capiro (Salazar et al., 2000) y más de 25% en S. tuberosum grupo Phureja cv Criolla Colombia (papa criolla) (Guzmán et al., 2012), razón por la que ha sido declarado como patógeno cuarentenario en Europa y Estados Unidos (European and Mediterranean Plant Protection Organization, 1979; United States Department of Agriculture - Animal and Plant Helath Inspection Service, 2000).
La detección de PYVV se ha realizado en muestras de folíolo mediante NASH (Nucleic Acid Spot Hybridization) (Salazar et al., 2000), RT-PCR (Offei et al., 2004; Guzmán et al., 2006- 2012; Guzmán y Rodríguez 2010; Wei et al., 2009), y en brotes de tubérculo mediante RT-PCR). qRT-PCR (López et al., 2006). A diferencia de otras técnicas, qRT-PCR tiene alta sensibilidad, especificidad, reproducibilidad, no requiere de un procesamiento post-PCR para la visualización de los resultados; adicionalmente, permite hacer una estimación precisa del número de copias de un gen viral presente en el tejido analizado. Es útil en diferentes estudios sobre biología de virus, asociación entre la intensidad de síntomas, acumulación viral, evaluación de resistencia, evaluación de la carga viral en la transmisibilidad del virus por el vector y eficiencia de replicación, entre otros. (Bustin, 2002; Heid et al., 1996; Mackay et al., 2002).
Hay poca información de la distribución de virus en plantas de papa infectadas, varios autores han demostrado que algunos virus se distribuyen en la planta de manera heterogénea (Leisner et al., 1992; Singh y Singh, 1996 y 1998; Gosálves et al., 2003; Fox et al., 2005; Sánchez-Navarro et al., 2007; Kogovšek et al., 2011); Kogovšek et al (2011) hacen uso de la técnica de qRT-PCR para analizar el título viral del Potato virus Y (PVY, Potyviridae) en diferentes órganos de plantas de Solanum tuberosum infectadas con el virus, reportando que el PVY presenta mayores títulos virales en tallos y foliolos sintomáticos, en foliolos no sintomáticos y tubérculos hay menores títulos virales, lo que demuestra que su distribución es heterogénea. Para PYVV se ha sugerido una distribución heterogénea del virus dentro del tubérculo (López et al., 2006).
Los ensayos de detección de PYVV se han limitado a material foliar y a brotes de tubérculo, pero no se han analizado otros órganos. Teniendo en cuenta que PYVV se limita al floema de la planta, el objetivo del trabajo fue analizar la distribución y acumulación del virus mediante RT-PCR y qRT-PCR, en diferentes órganos de plantas de papa criolla sintomáticas y NS infectadas con el virus, por transmisión a través del vector natural. Los resultados aportan al conocimiento sobre la distribución y detección del PYVV en papa criolla y son útiles en programas de fitomejoramiento, procesos de indexación de plantas de papa, programas de cuarentena y de certificación de semillas de papa.
Materiales y métodos
Material vegetal y aislados virales
Se tomaron dos plantas de S. tuberosum grupo Phureja cv Criolla Colombia, proveniente del municipio de Chipaque (Cundinamarca) con una edad aproximada de dos meses que expresaban diferentes niveles de amarillamiento intervenal: una de sintomatología moderada (M) y otra de sintomatología severa (S). Estas fueron mantenidas en matera utilizando como sustrato una mezcla de tierra : cascarilla de arroz en relación (3:1). Estas plantas se denominaron plantas donadoras de PYVV.
Adicionalmente, se utilizaron plantas de S. tuberosum grupo Phureja Clon 1 obtenidas a partir de cultivo de meristemos in vitro adquiridas en el Laboratorio de cultivo de tejidos y de biología molecular de plantas del IBUN. Estas plantas se denominaron plantas receptoras de PYVV.
Transmisión de PYVV con vector natural T. vaporariorum
Los ensayos de transmisión se realizaron en el invernadero (no controlado) de la Facultad de Agronomía de la Universidad Nacional de Colombia en jaulas entomológicas selladas con malla anti-áfido (velo suizo que se utiliza de rutina como malla aislante de vectores). Las temperaturas ambientales fluctuaron entre 5 y 27°C
Se estableció una cría de T. vaporariorum en plantas de calabacín (Cucurbita pepo). Aproximadamente 200 adultos avirulíferos de T. vaporariorum (confirmados previamente como negativos para PYVV por RT-PCR del gen CP) fueron liberados en una jaula entomológica que contenía dos plantas madres donadoras de PYVV (S ó M), confirmadas previamente por RT-PCR como positivas para el gen CP de PYVV. Aunque la mosca blanca transmite al virus de una manera semipersistente, es decir en pocos minutos u horas, en el presente estudio, las moscas blancas se mantuvieron en jaula entomológica durante un periodo de 48 horas para garantizar que se alimentaran del material infectado con PYVV y poder obtener individuos virulíferos (confirmados por RT-PCR).
Se liberaron 50 adultos virulíferos de T. vaporariorum en la jaula con un tamaño aproximado de 1m x 1m, donde se mantenían 12 plantas receptoras (confirmadas como negativas por RT-PCR para el gen CP de PYVV), y se dejaron por un periodo de 48 horas para garantizar el proceso de alimentación en la planta donadora y de la transmisión a las plantas receptoras.
Posteriormente, se utilizó el insecticida Karate® 1.5 cc/L para eliminar la mosca blanca de la jaula entomológica donde se mantenían las plantas receptoras. Estas se mantuvieron en jaulas entomológicas en el invernadero de la facultad de Agronomía UN, y se les realizó un seguimiento de expresión de síntomas, desde la aparición de amarillamiento de nervaduras secundarias en el ápice de las hojas. Además, se realizó un seguimiento a la infección del virus mediante la detección del gen CP de PYVV por RT-PCR.
Para evaluar la acumulación del virus en diferentes órganos se seleccionaron tres plantas receptoras cuando completaron 4 meses de edad, las cuales fueron positivas para CP-PYVV mediante RT-PCR, una planta nos sintomática (NS) , una con expresión moderada (M) de síntomas y una con expresión severa (S). Se realizó un muestreo aleatorio de diferentes órganos de cada planta tomando foliolo, pecíolo, tallo aéreo y subterráneo, raíz, pedúnculo floral, pétalo, antera (tabla 1).
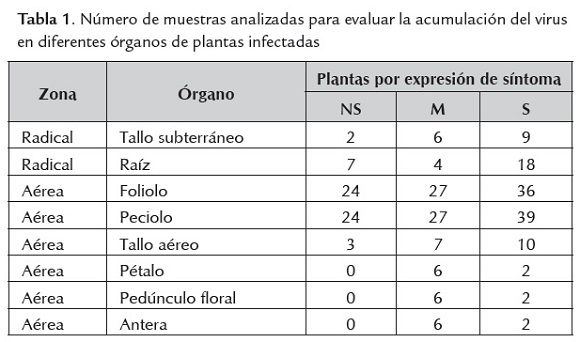
Detección de PYVV por RT-PCR convencional y en tiempo real
La extracción ARN se realizó de material foliar de las plantas donadoras y receptoras para confirmar la presencia/ausencia de PYVV por RT-PCR y de los diferentes órganos de las plantas infectadas descritas en la tabla 1, para lo cual se utilizó el reactivo Trizol® (Invitrogen) de acuerdo a las instrucciones del fabricante. Todos los extractos fueron tratados con 20 U de DNasa I durante 15min a 37°C para descartar la contaminación con DNA.
El gen CP de PYVV se amplificó a partir de los extractos de ARN de los diferentes órganos de las plantas analizadas, utilizando RT-PCR convencional y en tiempo real con los iniciadores y sondas TaqMan® descritos en la tabla 2.
Retrotranscripción
La síntesis de cDNA se llevó a cabo en un volumen final de 10μl adicionando una mezcla de 1X buffer de reacción (Epicentre), 1mM de dNTPs (Bioline), 10mM de DTT (Epicentre), 0.4μM de primer 3´ (tabla 2), 1.6U de RNase inhibitor (Fermentas), 8U de MMLV HP (Epicentre) y 100ng de ARN. Se incubó durante una hora a 42°C seguida de una denaturación a 70°C por 10min. El cDNA se repartió en dos fracciones: una para RT- PCR convencional y otra para la reacción de qRT-PCR.
PCR
Las reacciones de PCR contenían 1.6 μl de cDNA, 1X de buffer NH4 (Bioline), 2.5mM de MgCl2 (Bioline), 0.4 μM de dNTPs (Bioline), 0.4 μM de cada primer (F2/3´) (tabla 2) y 1U de Biolasa (Bioline) en un volumen final de 10μl. La amplificación consistió de una denaturación inicial a 94ºC durante 3min, seguido por 35 ciclos de denaturación a 94 ºC durante 1 min, alineamiento a 55 ºC durante 1 min y extensión a 72 ºC durante 1 min y una extensión final a 72 ºC durante 10 min. Como control positivo se incluyó una muestra foliar de S. tuberosum grupo Phureja cv Criolla Colombia con síntomas de PYVD y como controles negativos una muestra foliar de una planta de Calamondino (Citrofortunella madurensis, Lour) infectada con el virus Citrus tristeza virus (CTV) de la familia Closteroviridae; y una muestra foliar de S. tuberosum grupo Phureja cv Criolla Colombia libre de virus (obtenida por cultivo in vitro de meristemos).
PCR en tiempo real
Las reacciones de qRT-PCR se realizaron con el equipo LightCycler 480® (Roche), usando la química de sondas TaqMan® para la detección de un fragmento de 79 pb del gen CP de PYVV, como control interno de la reacción se amplificó un fragmento de 79 pb del gen Citocromo oxidasa (COX, gen de expresión constitutiva en la planta). Se utilizaron las sondas e iniciadores reportados por López et al. (2006) con algunas modificaciones en el marcaje de las sondas (tabla 2).
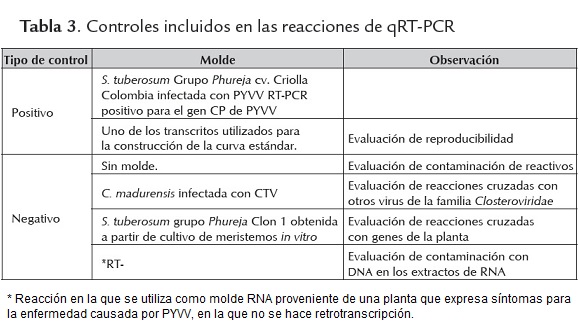
Para la reacción se utilizó 1X de buffer NH4 (Bioline), 5.5 mM de MgCl2 (Bioline), 0.5 mM de dNTPs (Bioline), 0.3 μM de cada primer, 0.24 μM de la sonda (tabla 2), 1U de Biolasa (Bioline), 2 μl de cDNA en un volumen final de 10μl. Los ciclos de amplificación consistieron de una denaturación durante 10 min a 95 ºC, 45 ciclos de amplificación con una denaturación a 95 ºC por 10 s y alineamiento a 55 ºC por 30 s ( siguiendo los protocolos estándar del laboratorio devirus vegetales IBUN). En las reacciones se incluyeron los controles descrito en la tabla 3.
Adicionalmente cada una de las muestras fue amplificada con una pareja de iniciadores para el gen COX con el objetivo de confirmar que las extracciones de ARN fueron realizadas apropiadamente y poder descartar falsos negativos en las amplificaciones del gen CP de PYVV.
Construcción de la curva estándar para la cuantificación absoluta
Para determinar el número de copias del gen CP de PYVV, se construyó una curva estándar utilizando diluciones seriadas de transcritos del gen CP de concentración conocida.
Para la obtención de los transcritos se utilizó un plásmido pGEM-T® (Promega) en el que el inserto era el gen completo de la proteína mayor de la cápside (donado por la estudiante de doctorado en Biotecnología UN, Patricia Rodríguez del laboratorio de Virus Vegetales del IBUN). El plásmido se linearizado con la enzima de restricción PstI (Promega) cuyo sitio de reconocimiento permite que el promotor T7 de la RNA polimerasa quede corriente arriba del inserto; la restricción se realizó de acuerdo a las instrucciones del fabricante. Posteriormente, el plásmido linearizado fue purificado utilizando el kit Wizard® DNA Clean-Up System (Promega). Finalmente, se realizó la transcripción in vitro teniendo en cuenta la siguiente mezcla de reacción: 1 µg de DNA, 0.5 mM rNTPs (Invitrogen), 5X de buffer de reacción (Invitrogen), 50 U de RNase inhibitor (Invitrogen), 10 mM de DTT (Invitrogen) y 30 U de RNA polimerasa T7 (Invitrogen) en un volumen final de 50 µl, el cual fue incubado durante 1 h a 37 °C, según indicaciones de la casa comercial.
Se realizaron diluciones seriadas del transcrito obtenido, las cuales servirían como patrones en la construcción de la curva estándar después de la determinación de su concentración utilizando el kit Quant-iTTM RiboGreen® RNA Assay (casa comercial Invitrogen).
La curva estándar se obtuvo graficando el valor Ct (en inglés threshold cycle) de cada una de las diluciones versus el logaritmo del número de copias del gen CP de PYVV. Cada punto de la curva estándar se corrió por duplicado. Para validar la cuantificación absoluta se corrió en tres ensayos independientes. La estimación del número de copias del gen CP de los estándares utilizados para la construcción de la curva estándar se hizo de acuerdo a la siguiente ecuación:

En donde la cantidad de transcrito de ARN se expresa en mg, el número de avogadro corresponde a 6.02 ×x1023, pb a número de pares de bases del transcrito, 1×x103 es un factor de conversión para convertir de g a mg, 340 es el peso molecular promedio de un nucleótido en una hebra sencilla de ARN. Se determinó el título viral en cada una de las muestras de las plantas por la interpolación del valor Ct en la curva de calibración normalizada.
Análisis estadísticos
Transmisión de PYVV utilizando el vector natural
Se calculó el promedio del número de copias para cada órgano evaluado, la desviación estándar y coeficiente de variación (CV%) inter-ensayo (porcentaje de la desviación estándar en comparación con el promedio del número de copias) utilizando el programa R (R Development Core Team, 2008). La distribución de todos los datos fue evaluada utilizando el test de normalidad Shapiro Wilk. Debido a que los resultados no presentaron una distribución normal se realizaron comparaciones para evaluar si se presentaban diferencias en la carga viral entre los órganos mediante la aplicación de una prueba Mann-Whitney acompañada de la corrección de Bonferroni a nivel de significancia (p < 0.05) para controlar la tasa de error (Yuan et al., 2006).
Resultados
Transmisión de PYVV utilizando el vector natural
Solamente después de 21 días post-transmisión (dpt), PYVV se detectó en las plantas receptoras a través de amplificación del gen CP de PYVV por RT-PCR convencional (figura 1). Los ensayos de detección por RT-PCR incluyen plantas receptoras que no expresaron síntomas (NS), trasmitidas a partir de plantas donadoras M (Carriles 2 al 6); plantas que expresaron síntomas, trasmitidas a partir de plantas donadoras S (Carriles 7 al 13).
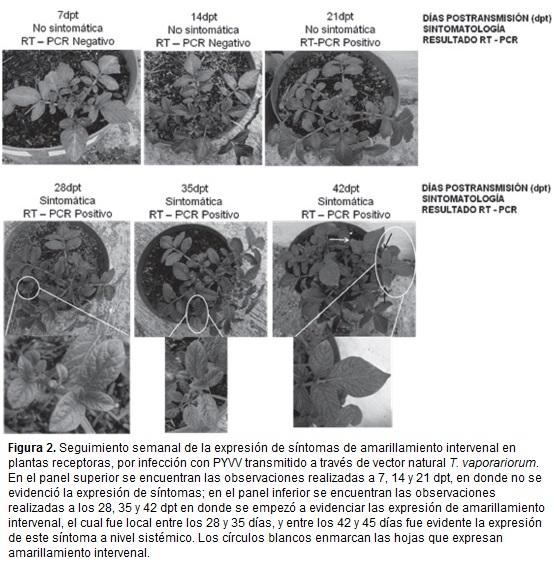
A pesar de la detección del virus en algunas de las muestras a los 21dpt, la expresión de síntomas se observó en algunos foliolos a partir de los 28 a 32 dpt. y a nivel sistémico entre los 42 a 45 dpt (figura 2).
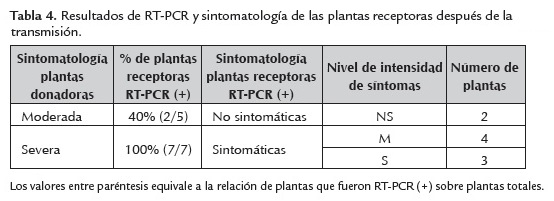
De las doce plantas utilizadas como receptoras, solamente se observó la expresión de amarillamiento intevenal en el 58.3% de las plantas (7/12), en las cuales se detectó el virus mediante RT-PCR. PYVV también fue detectado en dos plantas que no expresaron síntoma de amarillamiento intervenal, lo que indica que el porcentaje de transmisión de PYVV fue del (75%) (9/12). Sin embargo, otros autores como (Gamarra,2002) informa que la eficiencia de trasmisión de PYVV es baja de 6% utilizando Solanum tuberosum Grupo Andígena (variedad Diacol-Capiro). En el presente estudio, el porcentaje de transmisión y la expresión de síntomas en las plantas receptoras después de la transmisión, dependió de la planta donadora de PYVV y de su título viral, es decir, que cuando se utilizó planta donadora S el porcentaje de transmisión fue del 100% (7/7) y todas las plantas receptoras expresaron síntoma de amarillamiento intervenal. Sin embargo, con la planta donadora M el porcentaje de transmisión sólo fue del 40% (2/5) y ninguna de las plantas expresó síntomas (tabla 4).
Detección del gen CP de PYVV por RT-PCR en diferentes órganos de plantas con diferente nivel de expresión de síntomas
A partir del ensayo de transmisión se seleccionaron tres plantas receptoras en las que se había detectado el virus por RT-PCR: una S, una M y una NS; las cuales fueron evaluadas mediante RT-PCR para analizar la distribución del virus en los diferentes órganos (tabla 1). El virus se detectó en todos los órganos evaluados de las tres plantas (figura 3). En la planta S se detectó el virus en el 92.3% (109/118) de las muestras analizadas, en la planta M en el 95.5% (85/89), y en la planta NS sólo se detectó el PYVV en el 46.6% de las muestras (32/60).
qRT-PCR para la detección de PYVV en diferentes órganos de plantas con diferente nivel de expresión de síntomas
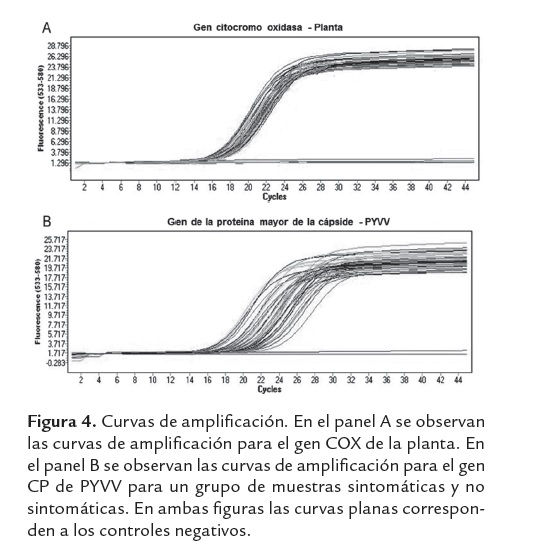
En todas las muestras evaluadas se amplificó el control interno (COX) los valores Ct fueron similares con un promedio de 15.9 ± 0.46 (figura 4A). El coeficiente de variación inter-ensayo no fue mayor al 7% indicando una buena reproducibilidad.
Aunque por RT-PCR convencional el virus se logró detectar en todos los órganos analizados de las tres plantas evaluadas, la detección no se dio en la totalidad de las muestras, lo que si se consiguió en la técnica de qRT-PCR, la cual permitió la detección del virus incluso en las muestras que habían resultado negativas por RT-PCR convencional (41/267), en donde el porcentaje de falsos negativos fue del 15,4%.
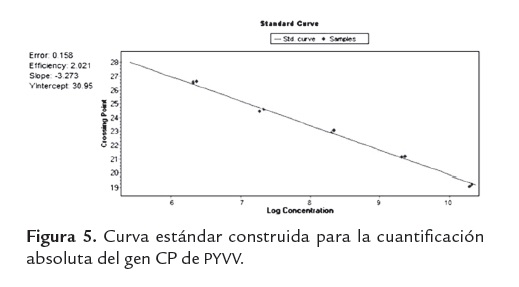
Para poder hacer la cuantificación absoluta del gen CP de PYVV se construyó una curva estándar con diluciones de transcritos de concentración conocida que iban desde 106 hasta 1010 (figura 5).
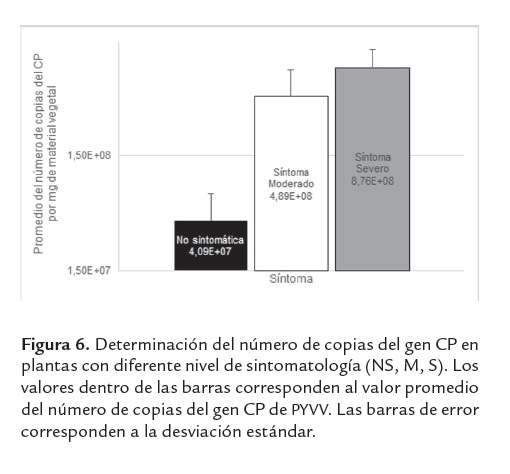
El valor Ct para el gen CP estuvo entre 14.57 y 24.47 (valor promedio de 18.89±2.22) (figura 4B), con un número de copias del gen CP entre 9.17×106 y 4.86×109 (valor promedio de 5.22×108 ± 7.05×107). La gran mayoría de falsos negativos obtenidos en la técnica convencional provenían de plantas NS, las cuales presentaron un promedio del número de copias del gen CP de 4 × 107, el cual fue significativamente menor (Prueba Mann Whitney, p<0.05) en casi un grado de magnitud, que las muestras provenientes de plantas sintomáticas (figura 6, Prueba Mann Whitney, p<0.05). El número de copias del gen CP en plantas con síntomas de amarillamiento intervenal (figura 6) no se vio afectado por la intensidad del síntoma (Prueba Mann Whitney, p=0.184), la diferencia del título viral entre la planta con síntoma severo y la planta con síntoma moderado no fue estadísticamente significativa (Prueba Mann Whitney, p=0.184).
Acumulación de PYVV en diferentes órganos de plantas infectadas
Los resultados de la figura 7 y de la tabla 5 indican que el título viral acumulado en los diferentes órganos de la planta NS, no presentaron diferencias estadísticamente significativas (Prueba de Mann Whitney, p=0.136), lo que sugiere que la distribución del virus en esta planta es homogénea, además, cada órgano de esta planta presenta un título viral significativamente menor que el acumulado en plantas sintomáticas (Prueba Mann Whitney, p<0.05)
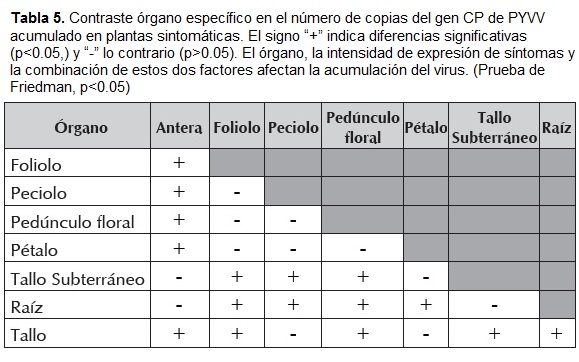
Al comparar el título viral acumulado en diferentes órganos de plantas sintomáticas (figura 7 y tabla 5) se encontró diferencias estadísticamente significativas entre órganos de la zona radical y los de la zona aérea (Prueba Mann Whitney, p>0.05), sugiriendo una acumulación heterogénea, con mayor acumulación en órganos de la zona aérea.
Discusión
El objetivo de este trabajo era evaluar la distribución de PYVV en diferentes órganos de plantas infectadas, para lo cual era necesario contar con plantas que solo estuvieran infectadas por este patógeno, debido a que infecciones mixtas pueden afectar algunos aspectos como la expresión de síntomas (reducción o exacerbación), rango de hospederos (García-Cano et al., 2006), y diferencias de tropismo y del título viral acumulado en diferentes órganos (Wege y Siegmund, 2007); estudios serológicos realizados en muestras de accesiones de bancos de germoplasma del grupo Phureja, han demostrado que algunas plantas mantenidas en campo pueden estar infectadas con uno o varios virus dentro de los que se reportaron: Potato virus S (PVS, Flexiviridae), Potato Leafroll virus (PLRV, Luteoviridae), Potato virus Y (PVY, Potyviridae) y Potato virus X (PVX, Flexiviridae) (Guzmán et al., 2010); a través de estudios de secuenciación profunda, Silvestre et al. (2012) encontraron que PYVV puede estar en coinfección con PVY; para garantizar que las plantas analizadas solo estuvieran infectadas con PYVV, se realizó transmisión a plantas libres de virus utilizando el vector natural que es T. vaporariorum, el cual no podría transmitir PVY porque su vector es Myzus persicae (Hemiptera: Aphidae). (Van Hoof, 1980).
La eficiencia de transmisión (porcentaje de plantas infectadas por el virus durante la transmisión) obtenida en este ensayo fue del 75% (9/12 plantas), aunque no fue del 100%, es posible que en condiciones naturales y climáticas adecuadas la transmisión de PYVV por vector pueda ser más alta. La metodología utilizada en este trabajo permitió la obtención de una eficiencia alta comparada con la obtenida por otros autores, que realizaron ensayos de transmisión de PYVV, Gamarra (2002) reportó un 6%, mientras que Arciniegas et al. (2008) reporta un porcentaje de transmisión del 12%.
Con respecto a la expresión de síntomas, los resultados fueron similares a los obtenidos por Arciniegas et al. (2008) en donde la expresión se dio entre los 28 y 32 días. En este trabajo se encontró que cuando la planta donadora era M, no se observó síntomas en las plantas receptoras (a pesar de esto se confirmó la presencia del virus mediante RT-PCR en algunas plantas que no expresaron síntomas), contrario a lo que sí ocurrió cuando la planta donadora era S. Esto podría indicar que el título viral de la fuente de inóculo está relacionado con la expresión de síntomas en la planta receptora, por lo tanto las plantas donadoras M tendrían un menor título viral que las plantas S, lo que influye sobre todo cuando la transmisión no es persistente o es semipersistente como en PYVV (Salazar et al., 2000; Díaz et al., 1990; Tamayo y Navarro 1984) debido a que no hay replicación del virus dentro del insecto y por lo tanto, durante el periodo de adquisición, el título viral alcanzado por el insecto vector debería ser proporcional al título viral de la planta fuente de inoculo.
En los ensayos de detección del virus en los diferentes órganos de plantas receptoras se encontró que la técnica de RT-PCR en tiempo real , siendo más sensible permitió la detección de PYVV en muestras que habían sido negativas (falsos negativos) por la técnica RT-PCR convencional, las muestras que fueron falsos negativos por la técnica convencional presentaron un título viral menor (4 × 107) que las que lograron ser detectadas mediante esta técnica (6 × 108), lo que expone una de las restricciones de la técnica convencional la cual depende de un límite en el número de copias para lograr un resultado positivo, conllevando a falsos negativos, que en este caso fue del 15,8%.
Los resultados de la técnica de RT-PCR en tiempo real indicaron que en la planta NS, la distribución del virus en los órganos analizados fue homogénea, este patrón de distribución con un bajo título viral podrían estar relacionados con la ausencia de expresión de síntomas en estas plantas, hay que tener en cuenta que estos ensayos fueron realizados en condiciones de laboratorio, pero en condiciones naturales, las plantas NS podrían ser el resultados de eventos de transmisión en donde el inóculo presenta bajo título viral (menor de 4 × 107 ) , o plantas que presentan infecciones tardías que hasta ahora están comenzando el desarrollo de la enfermedad.
Por otro lado, las plantas que expresaron síntomas presentaron una distribución diferencial del virus (distribución heterogénea), aspecto reportado por otros autores; Singh y Singh (1996 y 1998) detectaron el virus Potato virus A (PVA, Potyviridae) en tubérculos y otros tejidos de plantas de papa con una distribución heterogénea; de la misma manera, Kogovšek et al. (2011) reportaron la detección de PVY en diferentes órganos de plantas infectadas, el cual presentó una distribución heterogénea con mayor acumulación en los foliolos; para nuestro caso la mayor concentración se presentó en la zona aérea; esto podría ser una estrategia de dispersión, porque al mantenerlo en esta zona, el virus quedaría más accesible a la mosca blanca, contribuyendo de esta manera con la dispersión del mismo, este aspecto no había sido estudiado previamente en PYVV, pero si en otros virus como por ejemplo Carnation etched ring virus (CERV, Caulimoviridae) y Carnation vein mottle virus (CVMV, Potyviridae), que son transmitidos por miembros de la familia Aphididae, estos presentaron una menor acumulación viral en la zona radical (Sánchez-Navarro et al., 2007), en casos de virus que son transmitidos por hongos que habitan en el suelo o nematodos, se ha descrito que estos se concentran principalmente en la zona radical, como es el caso de los Carmovirus que son transmitidos por hongos del suelo (Gosalvez et al., 2003) y Tobravirus que son transmitidos por nematodos (Schmitt et al., 1998; MacFarlane y Popovich, 2000).
El folíolo fue uno de los órganos que presentó mayor carga viral en plantas sintomáticas, y este es el órgano del que prefiere alimentarse la mosca blanca, sumado a esto PYVV es un patógeno limitado al floema y la mosca blanca se alimenta del floema (Cohen et al., 1996), estos aspectos podrían resultar en un mayor éxito de transmisión. Los hábitos alimenticios de la mosca blanca reforzarían la idea de que el virus se acumula en las plantas de manera que sea muy accesible al vector, apoyando el concepto de un proceso de coevolución entre el vector y el virus (Lovisolo et al., 2003).
Hay que tener en cuenta que este virus también puede ser transmitido vegetativamente a través de tubérculo-semilla, aspecto que no se evaluó en este trabajo porque ya se había abordado previamente; Guzmán-Barney et al. (2012) en un trabajo realizado en material colectado en un cultivo del municipio de Mosquera (Cundinamarca), reportaron que el virus se acumula en los tubérculos de manera heterogénea en la zona radical, tanto en plantas sintomáticas como NS como en plantas sintomáticas, los títulos virales reportados en ese trabajo se encontraron entre 3.42×102 a 6.01×108, lo que podría explicar la falta de expresión de síntomas en algunos individuos de la progenie obtenidos a partir de plantas sintomáticas.
Sería interesante hacer ensayos de detección del virus en semilla sexual, lo que permitiría dilucidar algunos aspectos de transmisión de este virus (transmisión sexual), porque hasta el momento no se ha demostrado si PYVV puede ser transmitido de esta manera, aunque la probabilidad de que PYVV presente este tipo de transmisión es baja, no se puede descartar que algunos eventos de dispersión sean llevados a cabo a través de este mecanismo. No hay muchos reportes sobre la detección de virus fitopatógenos en semilla sexual, Maruthi et al. (2005) reportaron que no se logró detectar Cassava brown streak virus (CBSV, Ipomovirus) en muestras de semilla aunque el virus fue detectado en muestras de flores y frutos, según estos autores posiblemente existe un mecanismo que excluye el virus del embrión y por esta razón este virus no es transmisible sexualmente, en este trabajo el virus se logró detectar en el órgano reproductor masculino (antera), es posible que PYVV tenga un mecanismo similar al de CBSV pero esto estaría por confirmarse.
La detección de plantas infectadas por virus y su eliminación es un paso importante en los programas de manejo y control para poder evitar su dispersión, por lo tanto, la incorporación de técnicas de detección con un alto grado de sensibilidad que permitan la detección del virus tanto en plantas sintomáticas como en no sintomáticas, es de vital importancia en procesos de diagnóstico para la detección de plantas infectadas. Este resultado es útil para el mantenimiento y sanidad de los bancos de germoplasmas, en procesos de fitomejoramiento, indexado de plantas contra PYVV, programas de cuarentena y en certificación de semilla. Además, esta técnica no solamente es útil en procesos de diagnóstico, también ha sido de gran utilidad en estudios de expresión de genes y en estudios que evalúan diversos aspectos de la biología de virus, en este caso se utilizó para evaluar la distribución del virus en plantas infectadas lo que además de aportar información del virus, es una base para la selección de órganos en los procesos de detección.
Conclusiones
Es de resaltar que este es el primer trabajo sobre la detección y distribución de PYVV utilizando tiempo real, en diferentes órganos de plantas de papa criolla que expresaban síntomas de amarillamiento de venas y sin la expresión de los síntomas (no sintomáticas) pero infectadas por el virus. La información generada puede ser de gran importancia para dilucidar algunos aspectos relacionados con la transmisión del virus a partir de semilla-tubérculo como por transmisión por vector y aún con la posibilidad de transmisión sexual pues se detectó en anteras. Faltan trabajos complementarios y más amplios sobre estos aspectos de la biología y distribución viral en la planta. También, los resultados de este trabajo pueden ser útiles en programas de fitomejoramiento, de indexado de plantas de papa contra PYVV, programas de cuarentena y de certificación de semillas de papa libres de virus.
Por primera vez se detecta al virus PYVV en diferentes órganos como peciolo, pedúnculo floral, antera, pétalo y raíz; de plantas de S. tuberosum Grupo Phureja (variedad Criolla Colombia) utilizando las técnicas moleculares de RT-PCR convencional y q-RT-PCR, siendo esta última la más sensible debido pues permitió la detección en muestras de plantas que habían dado resultado negativo por la técnica convencional de RT-PCR. Adicionalmente, es la primera vez que se analiza la distribución del virus PYVV como homogénea o heterogénea en los órganos analizados y en las plantas sintomáticas y no sintomáticas. Las plantas no sintomáticas presentaron títulos virales menores y en distribución más homogénea dentro del órgano que las plantas que expresaban síntomas. Por lo tanto se pueden realizar otros estudios que aporten a la comprensión de la biología de PYVV dentro de la planta, puesto que anteriormente solo se había reportado el uso de RT-PCR con fines del diagnóstico viral. La técnica de qRT-PCR permitió la detección viral en un mayor porcentaje de muestras (mayor al 90%) y una mayor sensibilidad para la detección viral en todos los órganos evaluados comparado con la técnica de RT-PCR convencional (15.8% de falsos negativos).
Agradecimientos
Esta investigación se realizó gracias al apoyo económico y técnico del Ministerio de Agricultura y Desarrollo Rural de Colombia y a Colciencias (201010016538). Al Instituto de Biotecnología de la Universidad Nacional de Colombia. A Carlos Barragán por el apoyo en los ensayos de transmisión, a Patricia Rodríguez por su apoyo en la construcción de la curva estándar y a Ángela Villamil, por los valiosos aportes realizados.
Referencias bibliográficas
1 Alba, V. 1950. Viropatógenos. Memorias de la Conferencia Latinoamericana de Especialistas en Papa (Bogotá, Colombia). pp. 52-58. [ Links ]
2 Arciniegas, N., Guzmán, M. y Ñustez, C. 2008. Metodología de evaluación de resistencia al virus del amarillamiento de las venas de la papa (PYVV) en genotipos de la colección central colombiana de Solanum Phureja. Memorias XXIV Congreso de la Asociación Colombiana de Fitopatología ASCOLFI. [ Links ]
3 Bustin, S. 2002. Quantification of mRNA using real-time reverse transcription PCR (RT - PCR): Trends and problems. Journal of Molecular Endocrinology. 29: 23-39. [ Links ]
4 Cohen, A., Henneberry, T. and Chu, C. 1996. Microstructure of feeding in nymphal Bemisia argentifolii in cotton and cantaloupe. Entomologia. Experimentalis Applicata. 78: 135-142 [ Links ]
5 Díaz M., Pulgarín J. y Saldarriaga A. 1990. Relaciones insecto-patógeno en el problema del amarillamiento de las venas de la papa. Revista Colombiana de Entomología. 16: 3-14. [ Links ]
6 European and Mediterranean Plant Protection Organization. 1979. Data sheets on quarantine organisms No. 29, Potato vein yellowing virus. Bulletin OEPP/EPPO Bulletin. 9 (2). [http://www.eppo.org/QUARANTINE/virus/Potato_yellow_vein_virus/PYVV00_ds.pdf] [ Links ].
7 Fox, A., Evans, F. and Browning, I. 2005. Direct tuber testing for Potato Y potyvirus by real-time RT-PCR and ELISA: reliable options for post-harvest testing?. EPPO Bulletin. 35: 93-97. [ Links ]
8 Gamarra, H. 2002. Transmisión del virus del amarillamiento de las venas de la papa en variedades y/o clones de Solanum tuberosum Linneo, 1753. Universidad Ricardo Palma. Facultad de Ciencias Biológicas. Lima, Perú [ Links ].
9 García-Cano, E., Resende, R, Fernández-Muñoz, R. and Moriones, E. 2006. Synergistic interaction between Tomato chlorosis virus and Tomato spotted wilt virus results in breakdown of resistance in tomato. Phytopathology. 96: 1263-1269. [ Links ]
10 Gosalvez, B., Navarro, J., Lorca, A., Botella, F., Sánchez-Pina, M. and Pallas, V., 2003. Detection of Melon necrotic spot virus in water samples and melon plants by molecular methods. Journal of Virological Methods. 113: 87-93. [ Links ]
11 Guzmán, M., Ruiz, E., Arciniegas, N. and Coutts, R. 2006. Occurrence and variability of Potato yellow vein virus in three departments of Colombia. Phytopathology. 154: 748-750. [ Links ]
12 Guzmán, M. y Rodríguez, P. 2010. Susceptibilidad de Solanum phureja (Juz. et Buk.) al virus del amarillamiento de las venas. Agronomía Colombiana. 28: 219-224. [ Links ]
13 Guzmán, M., Román, V., Franco, L. y Rodríguez, P. 2010. Evaluación serológica de cuatro virus en accesiones colombianas de papa (Solanum spp.). Agronomía Colombiana. 28(2): 225-234. [ Links ]
14 Guzmán,-Barney, M., Franco-Lara, L., Rodríguez, D., Vargas, L., Fierro, J. 2012. Yield losses in Solanum tuberosum group Phureja cultivar Criolla Colombia in plants with symptoms of PYVV in field trials. American Journal of Potato Research. 89: 438-447. [ Links ]
15 Guzmán,-Barney, M., Hernández, A. and Franco-Lara, L. 2012. Tracking foliar symptoms caused by tuber-borne Potato yellow vein virus (PYVV) in Solanum Phueja (Juz et Buk) Cultivar "Criolla Colombia". American Journal of Potato Research. 90(3): 284-293. [ Links ]
16 Heid, C., Stevens, J., Livak, K. and Williams, M. 1996. Real time quantitative PCR. Genome Methods. 6: 986-994. [ Links ]
17 KogovŠek, P., Kladnik, A., Mlakar, J., TuŠek Šnidariè, M., Dermastia, M., Ravnikar, M. and Pompe-Novak, M. 2011. Distribution of Potato virus Y in potato plants, organs, tissues, and cells. Phytopathology. 101(11): 1292-1300. [ Links ]
18 Leisner, S., Turgeon, R. and Howell, S. 1992. Long Distance Movement of Cauliflower Mosaic Virus in Infected Turnip Plants. Molecular Plant-Microbe Interactions. 5: 41-47. [ Links ]
19 Livieratos, I., Eliasco, E., Müller, G., Olsthoorn, R., Salazar, L., Pleij, C. and Coutts, R. 2004. Analysis of the RNA of Potato yellow vein virus: Evidence for a tripartite genome and conserved 3'-terminal structures among members of the genus Crinivirus. Journal of General Virology. 85: 2065-2075. [ Links ]
20 López, R., Asencio, C., Guzmán, M. and Boonham, N. 2006. Development of real-time and conventional RT-PCR assays for the detection of Potato yellow vein virus (PYVV). Journal of Virological Methods. 136: 24-29. [ Links ]
21 Lovisolo, O., Hull, R. and Rösler, O. 2003. Coevolution of viruses with hosts and vectors and possible paleontology. Advances in Virus Research. 62: 325-379. [ Links ]
22 MacFarlane, S. and Popovich, A. 2000. Efficient expression of foreign proteins in roots from tobravirus vectors. Virology. 267: 29-35. [ Links ]
23 Mackay, I., Arden, K. and Nitsche, A. 2002. Real time PCR in Virology. Nucleic Acids Research. 30: 1292-1305. [ Links ]
24 Martelli, G., Agranovsky, A., Bar-Joseph, M., Boscia, D., Candresse, T., Coutts, R., Dolja, V., Falk, B., Gonsalves, D., Jelkmann, W., Karasev, A., Minafra, A., Namba, S., Vetten, H., Wisler, G. and Yoshikawa, N. 2002. The family Closteroviridae revised. Archives of Virology. 147: 2039-2044. [ Links ]
25 Maruthi, M, Hillocks, R., Mtunda, K., Raya, M. Muhanna, M., Kiozia, H., Rekha, A., Colvin, J and Thresh, J. 2005. Transmission of Cassava brown streak virus by Bemisia tabaci (Gennadius). Journal of Phytopathology. 153: 307-312. [ Links ]
26 Offei, S., Arciniegas, N., Müller, G., Guzmán, M., Salazar, L. and Coutts, R. 2004. Molecular variation of Potato yellow vein virus isolates. Archives of Virology. 149: 821-827. [ Links ]
27 R Development Core Team. 2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. [ Links ]
28 Rodríguez, P., Cháves, G., Franco, L. and Guzmán, M. 2009. Low molecular variability of Potato yellow vein virus (PYVV) isolated from Solanum phureja and Solanum phureja in Colombia. Ponencia en Joint Meeting of the Florida Phytopathological Society and the American Phytopathological Society Caribbean Division. Florida. [ Links ]
29 Salazar, L., Müller, G., Querci, M., Zapata, J. and Owens, R. 2000. Potato yellow vein virus. Its host range, distribution in South America and identification as a Crinivirus transmitted by Trialeurodes vaporariorum. Annals of Applied Biology. 137: 007-019. [ Links ]
30 Sánchez-Navarro, J., Cañizares, M., Cano, E. and Pallás, V. 2007. Plant tissue distribution and chemical inactivation of six Carnation viruses. Crop Protection. 26: 1049-1054. [ Links ]
31 Silvestre, R., Villamil, A., Guzmán, M., Cuellar, W. and Kreuze, J. 2012. Genetic diversity of PVY isolates infecting Solanum spp. In the Andes by siRNA sequencing. Memorias del XX v Congreso de la Asociación Latinoamericana de la Papa. [ Links ]
32 Singh, M. and Singh, R. 1996. Factors affecting detection of PVY in dormant tubers by reverse transcription polymerase chain and nucleic acid spot hybridization. Journal of Virological Methods. 60: 47-57. [ Links ]
33 Singh, M. and Singh, R. 1998. Specific detection of Potato virus A in dormant tubers by reverse-transcription polymerase chain reaction. Plant Disease. 82: 230-234. [ Links ]
34 Schmitt, C., Mueller, A., Mooney, A., Brown, D. and MacFarlane, S. 1998. Immunological detection and mutational analysis of the RNA2-encoded nematode transmission proteins of Pea early browning virus. Journal of General Virology. 79: 1281-1288. [ Links ]
35 Tamayo, P. y Navarro, N. 1984. Aumenta la incidencia del virus del amarillamiento de venas de la papa en Antioquia. ASCOLFI INFORMA (Bogotá). 19: 40-42. [ Links ]
36 United States Department of Agriculture - Animal and Plant Helath Inspection Service. 2000. Regulated plant pest list. [http://www.aphis.usda.gov/import_export/plants/plant_imports/downloads/RegulatedPestList.pdf] [ Links ].
37 Van Hoof, H. 1980. Aphid vectors of potato virus Y. Netherlands Journal of Plant Pathology. 86: 159-162. [ Links ]
38 Wege, C. and Siegmund, D. 2007. Synergism of a DNA and an RNA virus: Enhanced tissue infiltration of the begomovirus Abutilon mosaic virus (AbMV) mediated by Cucumber mosaic virus (CMV). Virology. 357: 10-28. [ Links ]
39 Wei, T., Lu, G. and Clover, R. 2009. A multiplex RT-PCR for the detection of Potato yellow vein virus, Tobacco rattle virus and Tomato infectious chlorosis virus in potato with a plant intenal amplification control. Plant Pathology. 58: 203-209. [ Links ]
40 Yuan, J., Reed, A., Chen, F. and Stewart, C. 2006. Statistical analysis of real time PCR data. BMC Bioinformatics. 7(85). [ Links ]
