










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista Colombiana de Biotecnología
versión impresa ISSN 0123-3475
Rev. colomb. biotecnol vol.16 no.2 Bogotá jul./dic. 2014
https://doi.org/10.15446/rev.colomb.biote.v16n2.47245
http://dx.doi.org/10.15446/rev.colomb.biote.v16n2.47245
ARTÍCULO DE INVESTIGACIÓN
Cultivo por lote de Wickerhamomyces anomaìus en un biorreactor a escala laboratorio para la producción de una poligalacturonasa
Batch culture of Wickerhamomyces anomaìus in a lab scale bioreactor for poligalacturonase production
Maria Alicia Martos1, Ana Paula Butiuk1, Natalia Lorena Rojas2, Roque Alberto Hours2
1 Facultad de Ciencias Exactas, Químicas y Naturales, Universidad Nacional de Misiones, Misiones, Argentina. amartos@fceqyn.unam.edu.ar.
2 Centro de Investigación y Desarrollo en Fermentaciones Industriales (CINDEFI), Universidad Nacional de La Plata - CONICET, La Plata, Argentina.
Recibido: febrero 5 de 2014 Aprobado: octubre 20 de 2014
Resumen
Wickerhamomyces anomalus, una levadura aislada de frutas cítricas en la provincia de Misiones, Argentina, produce una poligalacturonasa (endo-PG) con capacidad macerante de tejidos vegetales. El objetivo del presente trabajo fue determinai los parámetros cinéticos y estequiométricos del crecimiento de W. anomalus y la producción de la enzima poligalacturonasa en un medio de cultivo sintético, operado en sistema por lote, en un biorreactor a escala laboratorio. Los cultivos se realizaron en un biorreactor de 4 l que contenía 3 l de un medio sintético compuesto por glucosa, pectina de citrus, vitaminas, aminoácidos, sulfato de amonio y sales, y se incubaron con agitación y aireación, a 30 °C durante 12 h. El transcurso de proceso fermentativo se siguió por medidas de biomasa, glucosa residual, actividad poligalacturonasa y contenido de O2 y CO2 de los gases a la salida del reactor. La velocidad específica de crecimiento máxima (|im) de W. anomalus fue de 0,337 h-1 y el rendimiento de biomasa producida (Yx/s) de 0,401 gx/gs. Al finalizar el cultivo, la actividad PG en el sobrenadante fue de PG de ~ 83,7 UE/ml. La actividad específica y la productividad obtenidas fueron de ~ 1,91. 104 UE/gx y ~ 9.301 UE/l.h, respectivamente. El cociente respiratorio fue cercano a 1 durante el proceso fermentativo. No se formó ningún otro producto, además de biomasa y CO 2 . El cultivo por lote resultó ser una buena alternativa para la producción de PG a partir de W. anomalus, obteniéndose un extracto con elevada actividad enzimática, en un medio de cultivo sintético y de bajo costo.
Palabras clave: Wickerhamomyces anomalus, poligalacturonasa, cultivo por lote, parámetros estequiométricos, parámetros cinéticos.
Abstract
Wickerhamomyces anomalus, a yeast isolated from citrus fruit peels in the province of Misiones, Argentina, produces a polygalacturonase (endo-PG) with maceration activity of vegetable tissues. The objective of the present work was to determine kinetic and stoichiometric parameters of W. anomalus growth and polygalacturonase production in a synthetic culture medium, operating in a batch-type bioreactor at laboratory scale. Cultures were performed in a bioreactor of 4 l, containing 3 l of a synthetic medium composed of glucose, citrus pectin, vitamins, amino acids, ammonium sulfate and salts, and were incubated with agitation (450 rpm) and aeration at 30 °C, during 12 h. The course of the fermentation process was followed by measuring biomass, residual glucose, polygalacturonase activity and O2 and CO2 content of outlet gases from the reactor. The maximum specific growth rate (Um) of W. anomalus was 0.337 h-1 and the biomass yield (Yx/s) was 0.40 gx/gs. At the end of the culture, PG activity in the supernatant was ~84 UE/ml. The specific activity and the productivity obtained were ~1.91 104 UE/gx and ~9,301 UE/l.h, respectively. Respiratory quotient was approximately 1.0 throughout the fermentation process. No other product different from biomass and CO2 was detected. Batch culture could be an adequate alternative for the production of polygalacturonase from W. anomalus and an extract with high enzymatic activity using a synthetic and economic culture medium could be obtained.
Key words: Wickerhamomyces anomalus, polygalacturonase, batch culture, kinetic parameters, stoichiometric parameters.
Introducción
Las enzimas pécticas son las responsables de la degradación de las pectinas presentes en la laminilla media y en la pared celular primaria de las plantas superiores. Según su modo de acción se clasifican en: poligalacturonasa (PG), pectinesterasa (PE), pectinliasa (PL) y pectatoliasa (PAL). Algunas pectinasas tienen la capacidad de hidrolizar restringidamente la protopectina presente en los tejidos vegetales, liberando pectina soluble con la consiguiente separación de las células sin producir mayores daños, proceso denominado maceración. De este modo las células conservan muchos de sus compuestos intracelulares, entre los cuales se cuentan carotenoides, vitaminas (C, B6, B9 y E) y flavonoides, compuestos que poseen reconocidos beneficios sobre la salud. El proceso de maceración mediante enzimas pécticas es empleado en la industria alimenticia para la obtención de alimentos destinados a niños y adultos mayores (Jayani et al., 2005; Tari et al., 2007).
Las enzimas pécticas son producidas por una gran variedad de microorganismos como hongos filamentosos, levaduras y bacterias. La producción de pectinasas de levaduras tiene algunas ventajas en comparación con la producción a partir de hongos filamentosos. Las levaduras son organismos unicelulares por lo que su crecimiento es relativamente simple y fácil de cambiar de escala y a muchas de ellas se las considera como microorganismos GRAS (Generally Recognized as Safe) (Souza et al., 2003; da Silva et al., 2005).
Teniendo en cuenta que el costo de los medios de fermentación es uno de los factores determinantes en la producción de enzimas pécticas, es importante formular medios de cultivos con el menor costo posible y que al mismo tiempo provean los requerimientos nutricionales necesarios para el crecimiento adecuado del microorganismo y la producción enzimàtica (Nighojkar et al., 2006). Los medios de cultivos definidos son convenientes porque brindan no solo reproducibilidad al sistema, sino también pureza para las proteínas que son secretadas al medio.
Una levadura pectinolítica fue aislada a partir de cascaras de frutas cítricas en la Provincia de Misiones, Argentina, la cual fue identificada como Wickerhamomyces anomalus, reclasificación de la especie Pichia anomala (Passoth et al., 2011). W. anomalus, al crecer en un medio de cultivo compuesto por Base Nitrogenada para Levaduras (Yeast Nitrogen Base, YNB), glucosa como fuente de carbono y energía (FCE) y pectina de citrus como inductor, excreta al medio una enzima con actividad poligalacturonasa (endo-PG, EC 3.2.1.15). Esta enzima es de gran importancia industrial ya que posee actividad protopectinasas (PPasa) o capacidad macerante de tejidos vegetales (papa, mandioca, zanahoria, etc.) (Martos et al., 2013a, Martos et al., 2013b). La potencial aplicación industrial de esta enzima justifica la realización de estudios relacionados con su producción a mayor escala.
El presente trabajo tuvo como objetivo estudiar el crecimiento de W. anomalus y la expresión de la enzima poligalacturonasa en un medio de cultivo sintético y de bajo costo en un biorreactor a escala laboratorio, operado en sistema por lote.
Materiales y métodos
Microorganismo
Wickerhamomyces anomalus, aislada a partir de frutas cítricas en la provincia de Misiones, Argentina (Martos et al., 2013a).
Medios de cultivos
a) Medio de conservación (medio YM): extracto de levadura (Sigma), 5 g/l; triptona (Difco-Becton Dickinson), 5 g/l; glucosa bacteriológica (Britania), 10 g/l; agar-agar (Britania) 18 g/l, a pH 5,0.
b) Medio de fermentación: glucosa bacteriológica (Britania), 10 g/l; pectina de citrus (Parafarm), 5 g/l; (NH4)2SO4, 3 g/l; KH2PO4, 1 g/l; MgSO4, 0,5 g/l; CaCl2, 0,1 g/l; solución de vitaminas (1000 x), 1 ml/l; solución de aminoácidos (100 x), 10 ml/l; solución de microelementos 1000 x, 1 ml/l; pH 5,0.Solución (1000 x) de vitaminas (Sigma) (|Jg/l): biotina, 2; pantotenato de Ca, 400; ácido fólico, 2; inositol, 2000; niacina, 400; ácido p-aminobenzoico, 200; piridoxina, 400; riboflavina, 200; tiamina, 400. Solución (100 x) de aminoácidos (Sigma) (mg/l): histidina, 10; metionina, 20 y triptófano, 20.
Solución de microelementos 1000 x (|g/l): H3BO3, 500; CuSO4-5H2O, 40; Kl, 100; FeCl3-6 H2O; 200; MnSO4-H2O, 400; NaMoO4-2H2O, 200; ZnSO4-7H2O, 400.
La FCE (glucosa y pectina), el sulfato de amonio, los fosfatos, el resto de las sales (MgSO4, CaCl2, solución de microelementos) y los aminoácidos, fueron esteri lizados por separado, en autoclave 15 min a 121 °C, previo ajuste del pH a 5,0. La solución de vitaminas se esterilizó por filtración utilizando membranas de nitrato de celulosa de 0,22 Mm de tamaño de poro (Sartorius). Antes de su utilización, se mezclaron las cantidades correspondientes de cada una de las soluciones en forma aséptica y se midió el pH del medio completo.
Crecimiento de W. anomalus en un biorreactor a escala laboratorio
Inóculo: se inocularon tres frascos Erlenmeyers de 500 ml que contenían 95 ml del medio de fermentación, con 5 mi de inoculo (DO620 = 0,96) proveniente de un cultivo joven (24 h) de W. anomalus en medio YM. Los mismos se incubaron a 30 °C con agitación (180 rpm), durante 20 h. Al cabo de este tiempo, los cultivos (300 mi) se cosecharon por centrifugación a 4068 x g a 5 °C, durante 10 min. El sobrenadante se descartó y las células se suspendieron en 100 ml de agua destilada estéril. Fermentación: Se utilizó un biorreactor LKB (Bromma, Suecia) de 4 l de capacidad. El mismo fue esterilizado en autoclave (121 °C, 15 min) con la solución de glucosa y pectina a la que se le agregó 500 |i de aceite de silicona como antiespumante (Sigma). El resto de las soluciones de nutrientes (aminoácidos, vitaminas, sales) y la suspensión de células se mezclaron, antes de su utilización, en un frasco Erlenmeyer de 1000 ml con salida lateral y se utilizó inmediatamente para inocular el biorreactor e iniciar el proceso fermentativo, obteniéndose un volumen final de 3 l. El cultivo se agitó a 450 rpm y se le suministró aire estéril con un caudal de 2,82 l/min. Luego de 5 min se comenzaron a tomar muestras; las mismas fueron centrifugadas a 6505 x g, durante 5 min a 5 °C. Las células decantadas se usaron para medir biomasa por peso seco y el sobrenandante se utilizó para medir pH, glucosa residual y actividad PG. Los cultivos se realizaron por duplicado y se tomaron para los cálculos los valores promedios.
Medida de gases: Las medidas del contenido en O2 y en CO2 de los gases de salida del reactor fueron realizadas mediante un detector de tipo paramagnético (Servomex, UK) y un detector infrarrojo (Horiba Pir 2000, Japón), respectivamente. Con estos datos y el valor del caudal de aire se determinaron las velocidades de consumo de O2 y producción de CO2, en condiciones normales de presión y temperatura, mediante las siguientes ecuaciones que surgen de balances de materia en fase gaseosa y líquida, planteadas por Cooney et al. (1977) (ec. 1 y ec. 2):

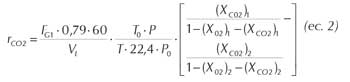
Los subíndices 1 y 2 representan valores a la entrada y salida del reactor respectivamente.
Siendo:
ro2:velocidad de consumo de O2 (moles/l.h).
rCO2: velocidad de producción de CO2 (moles/l.h).
FG: caudal de aire (l/min).
0,79: fracción molar de nitrógeno en el aire.
60: factor que convierte minutos a hora.
VL: volumen de medio (l).
T0 y T: 273 °K y temperatura de trabajo respectivamente.
Po y P: 760 mmHg y presión de trabajo respectivamente.
22,4: volumen (l) ocupado por un mol de gas en condicio nes normales de temperatura y presión.
Xo2 y fracción molar de oxígeno y anhídrido carbónico, XCO2: respectivamente.
El coeficiente respiratorio (CR) se calculó como la relación entre la velocidad de formación de CO2 (rCO2) y la velocidad de consumo de O2 (rO2).
La cantidad total de O2 consumido y de CO2 producido durante el cultivo se calculó como la integral de las respectivas velocidades en función del tiempo, empleando el programa Sigma Plot:
Determinaciones analíticas
Actividad poligalacturonasa: la actividad PG se midió
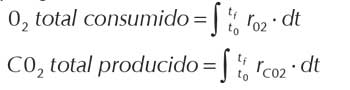
utilizando como sustrato 2 g/l de ácido poligalacturónico (APG, Sigma), en buffer acetato de sodio/ácido acético 0,2 M, pH 5,0. La reacción se efectuó a 37 °C, 10 min y se determinaron los grupos reductores liberados por el método del DNS (Miller, 1959). Como referencia se utilizó ácido galacturónico (Sigma). Una unidad enzimática (UE) se definió como la cantidad de enzima que libera 1 mmol del monómero correspondiente por min en las condiciones de ensayo.
Determinación de biomasa: el crecimiento microbiano (g/l) se determinó por medidas de peso seco. Luego del proceso de centrifugación, las células fueron lavadas con agua destilada, centrifugadas y colocadas en estufa a 80 °C, hasta peso constante (Cavalitto et al., 2000).
Glucosa residual: se utilizó el método de glucosa oxidasa-peroxidasa (Glicemia, Wiener, Argentina). Las determinaciones se realizaron por triplicado y se tomaron para los cálculos los valores promedios.
Resultados y discusión
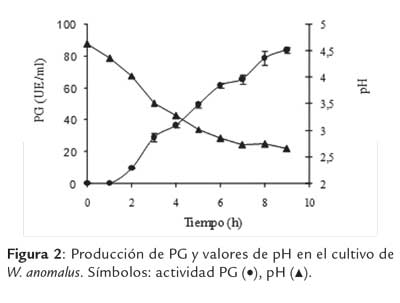
En la figura 1 se presentan los perfiles de crecimiento y concentración remanente de la FCE (glucosa) y en la figura 2 los valores de actividad PG y pH, durante el cultivo de W. anomalus en el biorreactor, operado en sistema por lote.
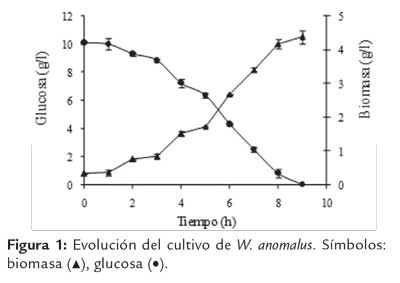
En la figura 1 se observa que a las 8 h de cultivo se consume la mayor parte de la FCE y finaliza el crecimiento exponencial. La biomasa total producida fue de 4,013 ± 0,26 gx/l, obteniéndose un rendimiento (Yx/s) de 0,401 gx/gs (yx/s = 0,466 C-molx/C-mols) en base a glucosa. De la curva de crecimiento microbiano se determinó un valor de Um de 0,337 h-1 (R2: 0,920), similar al reportado en trabajos anteriores para el cultivo de W. anomalus en frascos agitados, usando el medio YNB, adicionado con glucosa y pectina de citrus (μm: 0,346 h-1) (Martos et al., 2013a).
Los valores de μm y rendimiento obtenidos en el presente estudio son similares a los informados para el crecimiento de la mayoría de las levaduras sobre monosacáridos. Así, Geotrichum klebahnii, al crecer en un medio sintético, presentó un rendimiento (Yx/s) de 0,49 gx/gs, y un valor de Um de 0,47 h-1 (Cavalitto et al., 2000), mientras que una cepa de S. cerevisiae Narince 3, aislada de uvas presentó un valor de μm de 0.535 h-1 al crecer en un medio de cultivo que contenía glucosa, extracto de levadura y sales (Karasu Yalgn & Özbas, 2004).
En la figura 2 se observa que la producción detectable de PG comenzó a las 2 h, obteniéndose al finalizar el cultivo un valor de actividad PG de 83,7 ± 1,79 UE/ml. La síntesis de la enzima estuvo directamente asociada al crecimiento.
En este cultivo se utilizó (NH4)2SO4 como fuente de nitrógeno. Si bien el uso de una sal de amonio simplifica el proceso de esterilización (puede hacerse en autoclave conjuntamente con el resto del medio), es sabido que el consumo de amonio implica el incre mento equivalente en la concentración de protones en el medio de cultivo, con el consiguiente descenso continuo en el pH, resultado que efectivamente se observó considerando que el mismo disminuyó en el transcurso de la fermentación desde un valor inicial de 4,72 hasta valores cercanos a 2,6 al finalizar el cultivo (Fig. 2). Estos bajos valores de pH alcanzados, podrían, de prolongarse el cultivo, afectar la estabilidad de la enzima PG, ya que en estudios previos se determinó que la enzima permanece estable en un rango de pH entre 3,0 a 6,0 (Martos et al., 2013b).
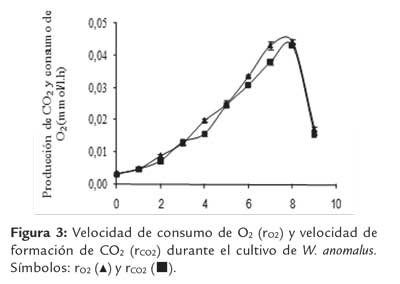
En la figura 3 se presentan las respectivas velocidades de consumo de O2 (rO2) y formación de CO2 (rCO2), en el transcurso del cultivo. Como puede observarse, los picos de las velocidades de consumo de O2 y producción de CO2 se obtuvieron alrededor de las 8 h y luego disminuyeron al consumirse totalmente la glucosa. Las curvas de ro2 y rCO2 corrieron paralelas, indicando que el CR fue cercano a 1 durante todo el tiempo de fermentación, valor razonable para un metabolismo completamente respiratorio.
Estudios previos indicaron que la levadura W. anomalus es incapaz de utilizar pectina, ácido poligalacturónico o sus productos de hidrólisis (ácido galacturónico) como FCE pero los necesita para inducir la síntesis de la enzima (Martos et al., 2003a). Por lo tanto, el crecimiento de W. anomalus en el medio sintético, utilizando glucosa como FCE, amonio como fuente de nitrógeno y asumiendo biomasa estándar (CH1,8O0,5N0,2), puede ser representado mediante la siguiente ecuación estequiométrica (expresada en C-mol):
CH2O + a NH3 + bO2 → yx/s CH1,8O0,5N0,2 + yco2/s CO2 + yp/sCHaObNc + WH2O
Siendo:
a: moles de nitrógeno consumido/C-mol de FCE
b: moles totales de O2 consumido/C-mol de FCE
yx/s: C-mol de biomasa producida /C-mol de FCE (rendi miento de biomasa)
yco2/s: moles totales de CO2 producidos/C-mol de FCE.
yP/s: C-moles de producto/C-mol de FCE.
Aplicando balances de carbono y grado de reducción a la ecuación de crecimiento microbiano se obtienen las siguientes ecuaciones:
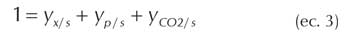
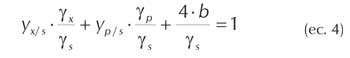
Siendo ys grado de reducción del sustrato; y< grado de reducción de la biomasa, yp grado de reducción del producto.
La tabla 1 muestra los valores de los parámetros estequiométricos del cultivo y los resultados del balance de carbono (ec. 3) y balance de grado de reducción (ec. 4), los que permiten determinar si se formó algún producto y si hubo consistencia interna en los resultados experimentales obtenidos. Los valores de grado de reducción se determinaron considerando NH3 como nivel de referencia.
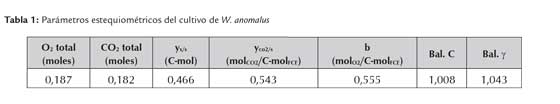
Los resultados obtenidos en los balances de carbono (ec. 3) y grado de reducción (ec. 4) indican que, en las condiciones de crecimiento, no hubo generación de ningún producto carbonado diferente a la biomasa y CO2. Según fue informado por otros autores, P. anomala (W. anomalus) es producida de manera eficiente considerando que es una levadura que presenta efecto Crabtree negativo, es decir que respira y produce biomasa en presencia de O2, independientemente de la concentración de azúcar presente en el medio (Passoth et al., 2011).
La actividad específica obtenida fue de 1,91. 104 ± 290 UE/gx y la productividad de 9.301±140 UE/l.h. Al hacer una estimación de costos, se obtuvo un valor de ~ US$ 2, para 4 l de medio de cultivo utilizado en el presente estudio (medio de fermentación). Considerando que la síntesis de la enzima estuvo directamente asociada al crecimiento, los datos cinéticos obtenidos en el presente estudio se podrían utilizar para el diseño de un cultivo por lote alimentado. En dichos cultivos, la alimentación restricta del nutriente limitante permite que las células crezcan a una alta densidad, lo que podría potenciar la expresión de la enzima, aumentando la productividad del proceso.
Conclusiones
El crecimiento de Wickerhamomyces anomalus en un biorreactor de 4 l, operado mediante sistema tipo por lote y usando un medio de cultivo a base de glucosa como FCE, pectina Cooney como inductor y sulfato de amonio como FN, se desarrolló con una velocidad de crecimiento máxima de 0,337 h-1. Se alcanzó un rendimiento de biomasa de 0,401 gx/gs y una actividad enzimática de ~ 84 UE/ml. Además, se obtuvo una actividad específica de ~1,91.104 UE/gx y una productividad de ~9.301 UE/l.h. Bajo estas condiciones de operación no se presentó formación de productos carbonados diferentes a biomasa y CO2.
El cultivo en sistema por lote resultó ser una buena alternativa para la producción de PG por W. anomalus, obteniéndose un extracto enzimático con elevada actividad enzimática en un medio de cultivo sintético y de bajo costo.
Referencias bibliográficas
1. Cavalitto, S.F.; Hours, R.A.; Mignone, C.F. 2000. Growth and protopectinase production of Geotrichum klebahnii in batch and continuous cultures with synthetic media. J. Ind. Microbiol Biotechnol. 25: 260-265. [ Links ]
2. Cooney, C.; Wang, H.; Wang, D. 1977. Computer-aided material balancing for prediction of fermentation parameters. Biotechnology and Bioengineering. 19: 55-67. [ Links ]
3. Da Silva, E.G.; de Fátima Borges, M.; Medina, C.; Piccoli, R.H.; Schwan, R. F. 2005. Pectinolytic enzymes secreted by yeasts from tropical fruits. FEMS Yeast Res. 5(9): 859-865. [ Links ]
4. Jayani, R.S.; Saxena, S.; Gupta, R. 2005. Microbial pectinolytic enzymes: A review. Process Biochem. 40(9): 2931-2944. [ Links ]
5. Karasu, Yalcin S.; Ózbas, Z.Y. 2004. Effects of different substrates on growth and glycerol production kinetics of a wine yeast strain Saccharomyces cerevisiae Narince 3. Process Biochem. 39(10): 1285-1291. [ Links ]
6. Martos, M.A.; Zubreski, E.R.; Combina, M.; Garro, O.A.; Hours, R.A. 2013a. Isolation of a yeast strain able to produce a polygalacturonase with maceration activity of cassava tissues. Food Sci Technol. 33 (2): 332-338. [ Links ]
7. Martos, M.A.; Zubreski, E.R.; Garro, O.A.; Hours, R.A. 2013b. Production of Pectinolytic Enzymes by the Yeast Wickerhanomyces anomalus Isolated from Citrus Fruits Peels. Biotechnol Res Int. 1: 1-7. [ Links ]
8. Miller, G.L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem. 31(3): 426-428. [ Links ]
9. Nighojkar, S.; Phanse, Y.; Sinha, D.; Nighojkar, A.; Kumar, A. 2006. Production of polygalacturonase by immobilized cells of Aspergillus niger using orange peel as inducer. Process Biochem. 41(5):1136-1140. [ Links ]
10. Passoth, V.; Olstorpe, M.; Schnürer, J. 2011. Past, present and future research directions with Pichia anomala. Antonie Van Leeuwenhoek. 99(1): 121-125. [ Links ]
11. Souza, J.V.B.; Silva, E.S.; Maia, M.L.S.; Teixeira, M.F. 2003. Screening of fungal strains for pectinolytic activity: endopolygalacturona se production by Paecilomyces clavisporus 2A.UMIDA.1. Process Biochem. 39:455-458. [ Links ]
12. Tari, C.; Gògus, N.; Tokatli, F. 2007. Optimization of biomass, pellet size and polygalacturonase production by Aspergillus sojae ATCC 20235 using response surface methodology. Enzyme Microbial Technol. 40(5): 1108-1116. [ Links ]
