











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
Metabolites are responsible for many essential functions and phenomena for plant growth and development, as well as plants' interaction with their environment. They include compounds of great importance for the growth and survival of plants, such as phenols, proteins, proline and carbohydrates (Petti and Scully, 2009; Yooyongwech et al. 2012; Mulekar and Huq, 2014; Van den Ende, 2014).
Phenolic compounds are secondary metabolites with a complex nature in plants. They are related to the processes of maturation, defense mechanisms, and response to biotic and abiotic stress. In turn, they have a great influence on the organoleptic properties of food products derived from plants (Kubo and Matsumoto, 1984; Cimato et al. 1990; Niemenak et al. 2006). While proteins, proline and carbohydrates are primary metabolites that, like phenols, may be involved in resistance mechanisms to biotic and abiotic stress, as is the case of proteins related to pathogenesis (PR) (Martinez-Esteso et al. 2009), proline is related to the response to drought in some plants (Wagner et al. 2003). The carbohydrates on the leaf surface have an important role in the early colonization and growth of fungus, as well as sucrose which facilitates the formation of biofilms of bacteria in the culture medium (Lindow and Brandl, 2003; Tomihama et al. 2006).
Theobroma cacao L. (T. cacao L.) is cultivated throughout the tropical areas of the world (Elwers et al. 2009), although it is a native plant of the tropical region of the Americas (Rusconi and Conti, 2010). It is used in popular medicine as an antiseptic, diuretic and parasiticide (Edwards et al. 2005), and these properties are associated with the varied metabolites it contains. The same metabolites can be found in different plant organs (leaves, stems, roots, flowers, seeds and trichomes) according to their biosynthetic route and their biological function (Ávalos and Pérez-Urria, 2009). Currently, there are studies with the aim to analyze the quality and flavor of chocolate through the characterization of changes in the composition of proteins and phenols during the maturity of the cacao fruits, the fermentation of the seeds and the key steps in the industrial production of chocolate (Pirovani et al. 2008; Guehi et al. 2010; Afoakwa et al. 2012a; Afoakwa et al. 2012b).
Plants with higher concentrations of proteins attract great interest because of their importance for the plants' development, as well as for human benefit. Phenols are also known for their benefits for human health in diverse activities, which include, regulation of fat absorption, anti-inflammatory effect, reduction of cardiovascular disease by reducing the synthesis of lipids, and as antioxidants (Guerrero et al. 2013; Gu et al. 2014; Sarria et al. 2014). They are also known for their antimicrobial activity against pathogens that affect plants and humans (Godard et al. 2009; Li et al. 2009; Vivot et al. 2009; Mishra et al. 2010; Pandey and Mishra 2010).
The leaf surface in the majority of plants is covered by modified cells of the epidermis that are called trichomes (Peter and Shanower, 1998). The compounds found in the trichomes of the leaf surface may be essentially involved in defense against different light conditions and the attack of pathogens and insects (Wagner, 1991; Peter and Shanower, 1998; Wagner et al. 2004; Ishida et al. 2008). In cacao plants, it was found that the glandular trichomes may provide the point of entry to Trichoderma species in the stem, contributing to systematic colonization of the tissue and establishment of an endophytic association (Bailey et al. 2009).
Therefore, and as far as we have knowledge, there is a lack of information in terms of the biochemical similarities and differences, as well as the relationship that exists between the trichomes and the chemical composition of the clones of T. cacao L. that are found in the germplasm bank of Padrón, INIA-Miranda, Venezuela. Therefore, the objective of this research was to study the existing relationship between the biochemical indicators (content of phenolic compounds, soluble proteins, free proline, total carbohydrates and lignins) and the number of trichomes to characterize the behavior in 26 clones of said bank.
Materials and Methods
Plant Material
The samples were collected in the Germplasm Bank of Estación Experimental de Padrón, INIA-Miranda, Miranda State, Venezuela (Table 1).
Table 1 Classification of the 26 clones of T. cacao collected in the Germplasm Bank of Estación Experimental de Padrón, INIA-Miranda, Miranda State, Venezuela. Types: Forastero (F) and Trinitario (T).
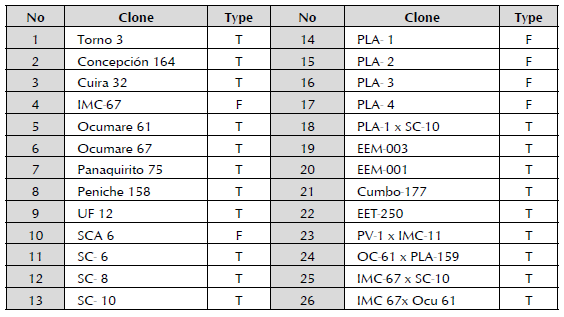
The samples of T. cacao were collected in the Germ-plasm Bank of Estación Experimental de Padrón, IN-IA-Miranda, Miranda State, Venezuela. Located at 41 m.a.s.l., at 10°13'14.1" N and 66°17'57.5" W, the sampled plants have been planted for 12 years in tropical humid forest in clay loam soil with pH 5.3, with flat topography and without irrigation. The density of the plantation was 1,600 plants per ha and it was fertilized with 100 g NPK fertilizer (12-12-17) + 50 g of urea. The humidity oscillated between 72-89% with an average rainfall of 2,441 mm.
For the biochemical analysis and count of trichomes, the four youngest green leaves were taken from plagiotropic branches of a medium age according to each evaluated clone of T. cacao L. They were immediately placed in liquid nitrogen and stored at -80 °C until the extractions were made.
Phenolic Compounds
The phenols were extracted and determined according to the method of Friend (1992) with modifications. To do this, the leaves (leaf blades without a petiole or central nerve) were macerated in liquid nitrogen into a fine powder (three replicas of 0.1 g of the mix of leaves from each plant material), and they were extracted three times with methanol for a final volume of 1 ml (soluble phenols). The residues were dried and hydrolyzed with 0.25 ml of 2 mol/l NaOH for 16 hours at 70 °C. Later, 0.25 ml of 2 mol/l HCl was added. The mixture was centrifugated at 15,000 g for 5 minutes and the supernatant was collected (wall-bound phenols). The residue was washed with 7 ml of distilled water and adjusted to pH 7.5 (with NaOH/HCl) and it was diluted to 10 ml with distilled water (lignins).
The soluble wall-bound phenols were determined with the Folin-Ciocalteu reagent. An aliquot of each extract was diluted with distilled water to 1 ml, and 0.1 ml of Folin-Ciocalteu reagent was added. After 5 minutes, 0.6 ml of saturated Na2CO3 was added to 1 mol/l NaOH was added. It was incubated for 60 minutes in the dark. The staining measurement used was in a spectrophotometer at 725 nm. The phenol concentration was expressed in milligrams per gram of fresh mass (FM) equivalent to a standard curve of chloro-genic acid. The total phenols were calculated with the sum of the soluble and wall-bound phenols.
For the quantification of lignins, 4 ml of 0.1 mol/l NaOH (pH 12) was added to 1 ml of each extract and 4 ml of phosphate buffer (pH 7) to another 1 ml. The absorbency was measured at 350 nm of extract at pH 12 against pH 7. The results were expressed as absorbance units (AU) for 0.1 g FM.
Soluble Proteins
The macerated leaves were used for the extraction of the soluble proteins just as was done for the phenols. One milliliter of extraction buffer was added (Urea: 6 mmol/l, EDTA: 10 mmol/l, NaH2PO4: 0.1 mol/l, β-mercaptoethanol: 14 mmol/l, Tween-20 (0.05%), Tris HCl: 10 mmol/l pH = 8) to 0.1 g of plant material (three replicas of the mix of leaves from each plant material). The mix was homogenized and centrifugated at a cold temperature of 4 °C for 10 minutes. The supernatant was collected for quantification. The concentration of TSP was determined according to Bradford, (1976) and the staining developed was measured in a spectrophotometer at 595 nm. The concentration was expressed in mg/g FM, equivalent to a standard curve of bovine blood serum albumin.
Total Carbohydrates and Free Proline
The extraction of carbohydrates and free proline was carried out on macerated leaves with the phenol procedure. Ten milliliters of methanol: 80% water (v:v) was added to 1 g of plant material (three replicas of the mix of leaves from each plant material). The mixture was shaken with a vortex and centrifugated at room temperature at 4,000 grams for 8 minutes. The extraction was repeated twice. The supernatants were collected, joined and filtered using filter paper. The ethanol phase was eliminated by vacuum in a rotary evaporator. The aqueous phase was collected for the quantifications.
The concentration of carbohydrates was determined according to the method of Dubois et al. (1956). To 1 ml of extract, 0.5 ml of phenol mixture: 5% water (v:v) and 2.5 ml of carefully concentrated H2SO4, was added. It was incubated at room temperature for 30 minutes. The staining measurement used was in a spectrophotometer at 492 nm. The concentration of total carbohydrates was expressed in μg/g FM, equivalent to a standard glucose curve.
The concentration of free proline was determined according to Bates et al. (1973). To 0.5 ml of plant extract, 0.5 ml of ninhydrin and 0.5 ml of acetic acid were added. Subsequently, it was shaken and placed in a bain-marie at 100 °C for 1 hour. Then, 1 ml of toluene was added and it was shaken strongly. The absorbency was determined in the organic phase at 520 nm. The concentration was expressed in μg/g FM, equivalent to a standard L-proline curve.
Trichome Count
The number and type of trichomes per visual field was directly counted in an optical microscope from the epidermal sheet of the abaxial side of the 26 clones' leaves (six replicas of the total leaves of each plant material). The evaluations were made in a LEICA DM 3000 microscope with a 20x lens.
Statistical Analysis
The statistical processing of the results was carried out with the Statistical Package for Social Sciences (SPSS) (Version 20 for Windows, SPSS Inc.). Parametric statistics were carried out (one-factor and two-factor ANOVA, and Tukey's range test). To study the relationship between the number of trichomes and the chemical composition of the leaves, the principal component analysis (PCA, principal component analysis) was used. This analysis was carried out with InfoStat (2011 Version, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina). The details of the statistical treatment appear in each figure or table of the results and discussion.
Results and Discussion
The concentration of soluble phenols, wall-bound phenols, lignin, soluble proteins and free proline in 26 clones of T. cacao is shown in Table 2.
Table 2 Concentration of soluble phenols (PHs), wall-bound phenols (PHwb), total phenols (PHt), lignin (Lg), soluble proteins (Ps), total carbohydrates (Ct) and free proline (Pf) in 26 clones of T. cacao L.
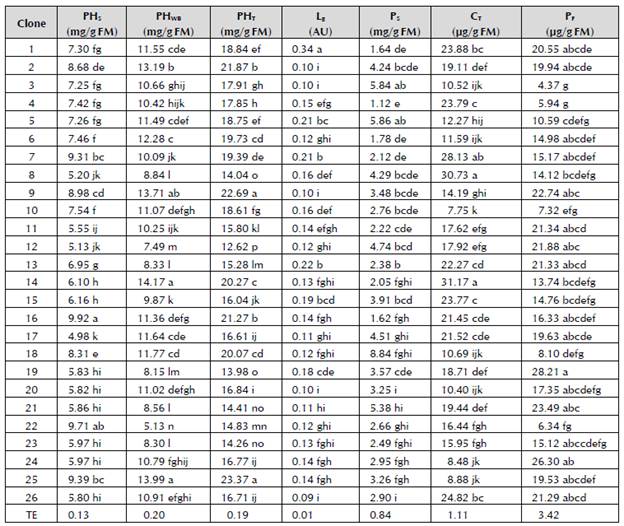
* Values in each column represent the mean of each treatment. Means with different letters differ significantly according to parametric statistics of simple ANOVA, Tukey's test, p ≤ 0.05, typical error (TE).
For the soluble phenols, Clones 16 and 22 showed the highest values without significant differences with 9.92 and 9.71 mg/g FM, respectively, and Clone 17 showed the lowest value with 4.98 mg/g FM. While in Clones 9, 14 and 25, the greatest concentrations of wall-bound phenols was found with 13.71, 14.17 and 13.99 mg/g FM, respectively and without significant differences. The lowest value was found in Clone 22. When analyzing the total phenols, Clones 9 and 25 were the highest without significant differences between them and with concentrations of 22.69 and 23.37 mg/g FM, followed by Clones 2 and 16 with 21.87 and 21.27 mg/g FM, and subsequently, Clones 6, 14 and 18 with 19.73, 20.27 and 20.07, respectively.
It can also be appreciated that for lignins, Clone 1 with 0.35 AU had the highest content of lignins, and the lowest values were found in clones: 2, 3, 9, 20 and 26, with 0.10, 0.10, 0.10, 0.10 and 0.09 AU, respectively, and without significant differences. These results indicate a greater concentration of wall-bound phenolic compounds than the soluble phenols and do not coincide with the clones that have a greater concentration of both kinds of phenols. Phenolic compounds are an important part of plants' active defense. These compounds can be synthesized and mobilized in stress conditions and are sometimes esterified to form part of the cell wall and of lignin (Fry, 1987; Nicholson and Hammerschmidt, 1992; Cabrera-Soto et al. 2009). In this case, all the plants were exposed to similar cultivation conditions without symptoms of disease, and therefore, it cannot be confirmed that the differences in the composition of phenols are associated with the response to a specific kind of stress.
It can be appreciated that in the case of soluble proteins, Clones 3, 5 and 18 showed the highest concentrations with 5.84, 5.86 and 8.84 mg/g FM, and the lowest value was found in Clone 4 with 1.12 mg/g FM. In the total carbohydrates, the highest concentrations were found in Clones 7, 8 and 14 with 28.13, 30.73 and 31.17 Mg/g FM, respectively, while the lowest concentration was found in Clone 10 with 7.75 Mg/g FM.
When analyzing the free proline, Clones 19 and 24 had higher concentrations with 28.22 and 26.29 Mg/g FM, while Clones 3, 4 and 22 had the lowest concentrations with 4.37, 5.94 and 6.34 Mg/g FM.
Regarding the number and type of trichomes, in Figure 1, it can be observed that glandular trichomes (indicated in Figure 1 a, g, h and i), non-glandular stellate trichomes (indicated in Figure 1 a, b, c and i) and non-glandular simple trichomes (indicated in Figure 1 d, e and f) were found.
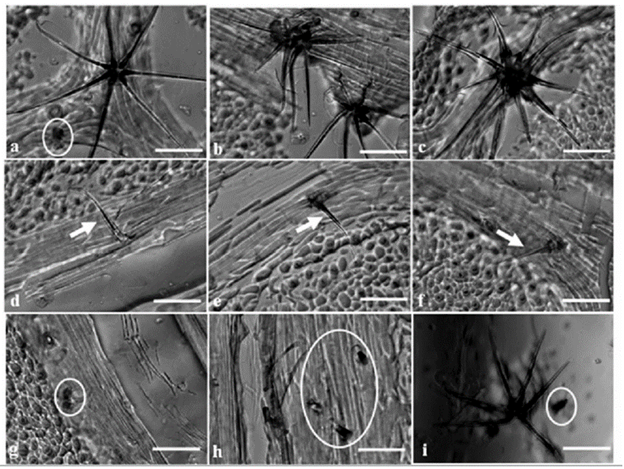
Figure 1 Types of trichome identified in the surface of adult plants of T. cacao L, Clone 5 (Ocumare 61). Complex, non-glandular stellate trichomes (a, b, c, i). Simple non-glandular trichomes (d, e, f). Glandular trichomes (a, g, h, i). Scale bar: 50 pm.
There were no differences in the type of trichome for the 26 assessed clones. Similar behavior was observed in all cases with glandular trichomes, non-glandular stellate trichomes and non-glandular simple trichomes, although the most frequent type was the non-glandular stellate trichome.
As can be observed in Figure 1 (a-i), the cacao tri-chomes showed diversity in terms of the number and type, as well as in the ramification of the non-glandular stellate trichomes. Additionally, the accumulation of colored compounds was found in them, which may be phenolic or other metabolites of interest, such as terpenes, among others (Tattini et al. 2000, McDowell et al. 2011). However, to corroborate the kind of compounds present, it is necessary to use other kinds of microscopy.
The number of trichomes varied depending on the clone, although the majority of the clones did not show significant differences. The largest amount of trichomes was found in Clones 5, 23 and 26 with 45, 40 and 40 trichomes per square millimeter, without significant differences, and the smallest amount in Clone 10 with 15 trichomes per square millimeter. In potato and tomato leaves, the trichomes contain phenols and phenol oxidizing enzymes (polyphenol oxidases), which can kill insects, including aphids (Peter and Shanower, 1998). That is why it is of great interest to group the clones by their majority compounds, as well as assessing whether there is a relationship between the number of trichomes and the chemical compounds of the T. cacao leaves.
Figure 2 shows the analysis of the relationship between the 26 studied clones of T. cacao with their content of free proline, soluble proteins, total carbohydrates, lignins, soluble phenols and wall-bound phenols, and the number of trichomes.
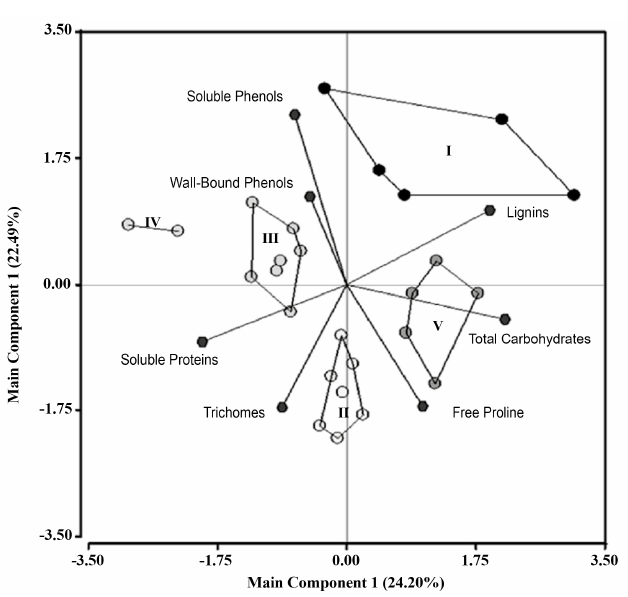
Figure 2 Relationship between the clones of 7. cacao L. Groups comprised by Main Component 1 in free proline, soluble proteins, total carbohydrates, lignins, soluble phenols, wall-bound phenols and number of trichomes. Groups: I (Clones 4, 16, 7, 10 and 1), II (Clones 17, 12, 15, 26, 21, 23 and 24), III (Clones 2, 5, 6, 9, 20, 22 and 25), IV (Clones 3 and 18) and V (Clones 14, 8, 13, 11 and 19).
The results show that in the bidimensional space, which explains 46.68% of the total variability, five homogenous groups were defined. Group 1 (I) comprised of Clones 4, 16, 7, 10 and 1, associated with the maximum concentration of lignins. Group 2 (II) comprised of Clones 17, 12, 15, 26, 21, 23 and 24, associated with plants with leaves presenting the highest number of trichomes. Groups 3 (III) and 4 (IV) comprised of Clones 2, 5, 6, 9, 20, 22 and 25; and 3 and 18, respectively, associated with high contents of soluble proteins, soluble phenols and wall-bound phenols. Finally, Group 5 (V) comprised of Clones 14, 8, 13, 11 and 19, associated with the gradient of maximum content of free proline and total carbohydrates. When comparing these two kinds of cacao (Forastero and Trinitario) in terms of the concentration of total phenols, lignins, soluble proteins and free proline (Table 3), it was found that the Trinitario clones have a higher content of soluble proteins and free proline than the Forastero clones. While the latter exceed the Trinitario variety in total phenols (soluble phenols + wall-bound phenols) and carbohydrates.
Table 3 Concentration of total phenols, lignin, soluble proteins, carbohydrates and free proline in Trinitario (T) and Forastero (F) clones of T. cacao L.
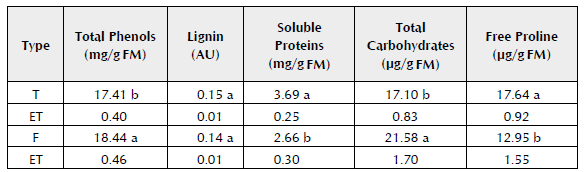
Values in each column represent the mean of each treatment. Means with different letters differ significantly according to parametric statistics of two-factor ANOVA, Tukey's test, p ≤ 0.05, and typical error (TE).
It is important to note that the substances produced by the aerial parts of the plants may be carbohydrates, organic amino acids, phenols and other compounds. The phenols include tannins, which are generally toxins because of their capacity to join to proteins, and they act as food repellents against many animals that could be harmful to the plants. However, they may have a positive effect on human health (Ávalos and Pérez-Urria, 2009). In the case of the total phenols (Table 3), their greater concentration in Forastero clones may be related to the fact that this kind of cacao is more resistant and of lower quality for the industry because of its high phenol content, which includes the tannins with a greater concentration in these clones (Figueira et al.1997).
The quantitative differences found in the phenol content (Table 2) between the clones are largely related to the genotype, because the plants had the same age and were grown in similar conditions. This coincides with the proposal of Zapata et al. (2000), who demonstrated in tobacco that the phenols depend on the age, the stage of development and the genotype. Niemenak et al. (2006) found that the total phenols, catechin, epicatechin and anthocyanin in fresh and fermented seeds of T. cacao L. depended on the genotype, but qualitative differences were not observed in the content of polyphenols regarding their genetic origin. The differences were more closely related to the growth conditions. However, in this work, differences were found in the concentration of phenols, carbohydrates, proteins and proline related to the genotype and also to the genetic origin, because in the Forastero clones, the concentration of total phenols and total carbohydrates was higher than in the Trinitario clones, as long as the Trinitario clones were in their majority soluble proteins and free proline (Table 4). Furthermore, all the evaluated indicators varied between the different clones (Table 3).
Coinciding with Chen et al. (2005), they propose that the soluble proteins, ribulose-1.5-bisphosphate carboxylase/oxygenase (RuBisCo) and its activase content, as well as the activity of the sucrose-phosphate synthase, decrease significantly, while the content of soluble sugars, inorganic phosphate, chlorophyll and Complex II of the photosynthesis increased in rice leaves acclimatized with air enriched with CO2. Currently, it is known that proteins are found in a low concentration in plant tissue compared to animals and bacteria. Furthermore, they are more difficult to extract in mature plants because of the thickness of the cell wall (Islam et al. 2004). In addition, the phenolic compounds combine with the proteins by covalent condensations (Carpentier et al. 2005), and proteins such as RuBisCo are very abundant, which may hinder the detection of those that are not so abundant (Neilson et al. 2010; Balbuena et al. 2011).
Matt et al. (2002) transformed tobacco plants with an antisense RuBisCo to study the consequences of damage to the metabolism of photosynthetic carbon on the metabolism of nitrogen and secondary metabolism. They demonstrated that the inhibition of photosynthesis and the decrease in the sugar content lead to the inhibition of the metabolism of nitrogen and dramatic changes in the composition of secondary metabolites of the leaves, such as chlorogenic acid and nicotine, which have a structure rich in carbon and nitrogen, respectively. They also demonstrated that the changes in the amino acid to sugar ratio were related to the nicotine to chlorogenic acid ratio. These results correspond with those presented in Table 4, where the group that presented the greatest concentration of phenols (Forastero clones), also had a greater concentration of carbohydrates.
On the other hand, proteins, carbohydrates and proline play very important roles in response to stress, and it has been demonstrated that in leaves infected by C. perniciosa, the soluble proteins decrease (Frias et al. 1991). It has also been found that in the leaves of T. grandiflorum plants infected by fungus, the chlorophyll content is affected and therefore, photosynthesis and carbohydrate synthesis are also affected, as well as variations occurring in the phenol concentrations (Oliveira et al. 1997).
As can be observed, the groups that are formed with PCA are not related to the type of cacao, as the Forastero and Trinitario clones are mixed within the different groups (Figure 2). In Table 3, the group of Forastero clones showed a greater concentration of carbohydrates and phenols, the Trinitario clones showed a greater concentration of proline and proteins, and there were no differences between the two groups for lignins. However, in the PCA, the clones were grouped in a different way. Five clones were associated because of their high lignin content (three Forastero clones and two Trinitario clones), another five because of their high concentration of proline and total carbohydrates (two Forastero and three Trinitario clones), nine because of their high protein and phenol content (all Trinitario clones), and seven with a higher number of trichomes (one Forastero clone and six Trinitario clones). Unexpectedly, in the PCA, the greatest number of accumulation structures of metabolites (trichomes) was more closely related to the greater concentration of soluble proteins and the free proline than to the rest of the measured parameters.
In the case of the trichomes, it is known that the combination of glandular and non-glandular trichomes confers resistance to different pests, because the glandular trichomes contain or secrete mixtures of chemical compounds and the non-glandular trichomes serve as a mechanical barrier, above all against insects (Peter and Shanower, 1998). The clones with a larger number of phenols were not associated with the greater trichome content. This does not discard the fact there are kinds of phenols that are found in the trichomes or that are secreted by them and are of importance because of their biological activity, which makes them interesting for pharmacy, pathogen control and obtaining pigments or aromatic substances. However, to select clones for the production of compounds in the trichomes, not only those of a greater number would have to be assessed, but also the profile of the metabolites present in the trichomes of each clone and the value of them because of their activity.
In the plants of the Solanum genus, there are six types of trichome, and in all of those of type 1, 4 and 6, genes were found for the biosynthesis of metabolites such as flavonoids, acetylated sugars and terpenoids (McDowell et al. 2011). To date, it has not been described what kind of compound may be in the trichomes in cacao, but it is known that there are glandular and non-glandular trichomes (Bailey et al. 2009), and that there is also a high phenol content. Therefore, there could also be synthesis and secretion of similar compounds to those described by Solanum.
Finally, the clones analyzed in this research were not visibly infected by any pathogen. However, within the 26 existing clones, there are some with a greater resistance to pathogens than others. Therefore, the concentrations of the analyzed compounds may indicate a greater amount of them in the resistant clones than in those susceptible to fungal diseases. For example, in the previous research carried out in the germplasm bank from which the evaluated material was selected, Clone 3 (high protein and phenol content) is resistant to P. palmivora and to C. frimbriata; Clone 8 (high carbohydrate and proline content) is moderately resistant to P. palmivora; Clone 4 (high lignin content) is resistant to C. frimbriata and has a low incidence of M. roreri; and Clone 24 (greater number of trichomes) is resistant to P. palmivora (Reyes et al. 1973; González et al. 2009; González et al. 2011; Morillo et al. 2012). Furthermore, when Clone 19 (high carbohydrate and proline content) was used as the mother clone in two crosses (19 x 4 and 19 x 22), the descendant of both showed a low incidence of P. palmivora in harvested fruits and a low incidence of M. perniciosa as plant brushes and in floral cushions. These crosses presented low rates of productivity, almond rate > 1.3, and corn rate < 22 (González et al. 2011). This corroborates that each group obtained offers the possibility of managing the clones depending on their chemical composition and future use.
Conclusions
This study demonstrated that there are five groups according to the existing relationship between the concentration of biochemical compounds and the trichomes of leaves of 26 clones of T. cacao. Five clones were associated because of their high lignin content; another five because of their high concentration of proline and total carbohydrates; nine because of their high protein and phenol content; and seven due to the greater number of trichomes according to the PCA, which explains 46.68% of the total variability. Furthermore, the group of Forastero clones showed the greatest concentration of total phenols and carbohydrates, while the Trinitario clones presented a greater concentration of proteins and proline. The results obtained may serve as a basis for the selection of clones, taking into account the content of the different evaluated parameters, depending on the interest of future research.

