










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.17 no.2 Bogotá July/Dec. 2015 Epub Aug 10, 2021
https://doi.org/10.15446/rev.colomb.biote.v17n2.45326
Artículos cortos
Efecto del anegamiento en la actividad de la enzima alcohol deshidrogenasa en raíces de maracuyá amarillo Passiflora Edulis var. Flavicarpa
* Bióloga, Universidad del Valle, Apartado Aéreo 25360, Cali, Colombia. keren.giraldo@correounivalle.edu.co
** MSc., Universidad del Valle, Apartado Aéreo 25360, Cali, Colombia.
*** Ph.D., Universidad del Valle, Apartado Aéreo 25360, Cali, Colombia. enrique.bravo@correounivalle.edu.co
**** Ph.D., Universidad del Valle, Apartado Aéreo 25360, Cali, Colombia. jaime.restrepo@correounivalle.edu.co
Bajo entornos naturales, las plantas de maracuyá amarillo frecuentemente se enfrentan a condiciones de anegamiento, un factor limitante para la producción generalizada de su cultivo, especialmente en tierras bajas inundables. El presente estudio se encargó de identificar la activación de la enzima alcohol deshidrogenasa (ADH) durante la inundación, como un posible mecanismo de sobrevivencia de plántulas de maracuyá amarillo. Se evaluó la actividad de la enzima ADH durante 0, 1, 3, 7, 9 y 14 días de tratamiento con inundación y sin inundación, en raíces de plántulas de maracuyá amarillo de tres meses y medio de germinadas. En las raíces de plántulas de maracuyá amarillo en condiciones de inundación, la actividad de la enzima ADH presentó un aumento significativo respecto a las plántulas en condiciones normales de riego (sin inundación). Esto sugiere que la actividad de la enzima alcohol deshidrogenasa, implicada en el metabolismo anaeróbico, es un posible mecanismo de supervivencia al anegamiento de plántulas de maracuyá amarillo en periodos cortos de inundación.
Palabras clave: actividad enzimática; ADH; fermentación alcohólica; inundación; metabolismo anaerobio
Under natural environments, yellow passion fruit plants often face flooded conditions as a limiting factor for the widespread production of the crop, especially in flood-prone lowlands. This study was carried out to identify the activation of the enzyme alcohol dehydrogenase (ADH) during the flood, as a possible mechanism for the survival of yellow passion fruit seedlings. The activity of the ADH enzyme was assessed on days 0, 1, 3, 7, 9 and 14, with and without flooding, on rooted yellow passion fruit seedlings germinated during three and a half months. In the roots of the yellow passion fruit seedlings in flood conditions, the activity of the ADH enzyme showed a significant increase compared to seedlings under normal irrigation (no flooding). These results suggest that the activity of the alcohol dehydrogenase enzyme involved in anaerobic metabolism, is a possible mechanism for survival of yellow passion fruit seedlings waterlogged in short periods of flooding.
Key words: enzymatic activity; ADH; alcoholic fermentation; flooding; anaerobic metabolism
Introducción
Durante la inundación o anegamiento, la transferencia de oxígeno y otros compuestos entre el suelo y la atmósfera se bloquea, y el poco O2 disuelto disponible en el agua es consumido por las plantas, fauna del suelo y microorganismos hasta su total agotamiento (Joshi & Kumar, 2012). Ante la baja disponibilidad de O2 (hipoxia) o ausencia de oxígeno (anoxia), se afecta el funcionamiento normal de las plantas, principalmente por la disminución de la síntesis de ATP, causando marchitamiento en las plantas anegadas (Sairam et al., 2008). A pesar de la crisis energética que sufren las plantas ante la deficiencia de oxígeno por efecto de inundaciones, se han encontrado en diferentes especies de cultivo adaptaciones que les permiten tolerar las condiciones anaeróbicas provocadas por el anegamiento (Nishiuchi et al., 2012). Entre estas adaptaciones encontramos cambios anatómicos como la formación de raíces adventicias con interconexiones longitudinales de espacios de gas llamados aerénquima, que permiten la aireación interna entre brotes y raíces (Voesenek et al., 2006), o la formación de barreras para evitar la pérdida radial de O2 a la rizósfera (Armstrong et al., 2000). También se producen cambios metabólicos que incluyen disminución de la absorción y de la asimilación de fuentes de nitrógeno, activación de la respiración anaerobia, mantenimiento del suministro de carbohidratos para la respiración anaeróbica, control de la acidificación citoplasmática y desarrollo del sistema de defensa antioxidante (Chen & Qualls, 2003; Kumutha et al., 2008; Oliveira et al., 2013; Roberts et al., 1984), los cuales permiten a las plantas seguir creciendo y sobrevivir en entornos de poca disponibilidad de oxígeno (Medina et al., 2009).
El oxígeno es vital para la producción de energía de la célula, y su presencia es determinante para las actividades metabólicas y la producción de ATP, al ser el aceptor de electrones en la vía de la fosforilación oxidativa. Es por esto que muchas de las investigaciones en plantas se han enfocado en la presencia y función de vías de fermentación como mecanismo de rescate metabólico cuando la respiración se detiene debido a la anoxia por el anegamiento del suelo (Dennis et al., 2000). En plantas, las vías de fermentación no se presentan bajo condiciones normales de oxígeno, pero su rápida inducción por un bajo suministro de oxígeno sugiere un mecanismo de supervivencia ante la poca oxigenación. El sistema de raíces es el objetivo principal ante la deficiencia de oxígeno por el anegamiento, debido a los diversos trastornos metabólicos que ocurren en el tejido de la raíz, que en última instancia afecta al rendimiento global de la planta (Kulkarni & Chavan, 2013). Estudios de la respuesta de raíces de plántulas de maíz al tratamiento anaeróbico han revelado que la síntesis normal de proteínas solubles aeróbicas se reduce, mientras que un conjunto de 20 proteínas anaeróbicas (ANPs) se sintetizan selectivamente, siendo estas identificadas como enzimas que intervienen en la glucólisis y en el metabolismo fermentativo (Sachs et al., 1980). De este grupo de proteínas anaeróbicas, la enzima que cataliza la fermentación alcohólica, la alcohol deshidrogenasa (ADH), es la más estudiada, y su mayor inducción por efecto de la anaerobiosis se ha demostrado en especies de importancia económica como Zea mays, Glycine max, Euterpe oleracea, Oryza sativa, Hordeum vulgare, Pennisetum glaucum, Dios-corea cayenensis, Ipomoea batatas y Echinochloa crus-galli (Borella et al., 2014; Fukao et al., 2003; Gaut & Clegg, 1991; Huang et al., 2002; Johnson et al., 1994; Menezes et al., 1995; Oluoha, 1995; Wignarajah et al., 1976). Este aumento significativo de la actividad de la enzima ADH, es una característica considerada de tolerancia al estrés por exceso de agua, razón por la cual es importante la exploración de mecanismos de resistencia a nivel metabólico en especies de importancia económica como el maracuyá amarillo, cuyo atributo podría ser utilizado en programas de mejoramiento del cultivo (Roorkiwal & Chand, 2012).
El maracuyá amarillo (Passiflora edulis var. flavicarpa Deg), es uno de los frutales de cultivo más apetecidos para el consumo en muchos países del mundo, encontrándose cultivos de gran producción en Colombia, Venezuela, Ecuador, Perú, Brasil, Hawai, Taiwán, India, Sud-África, Srilanka, Kenia, Nueva Guinea y Australia (Potosí et al., 2008). En Colombia, en el Valle del Cauca se tienen cultivos de maracuyá amarillo en áreas de siembra de 919 hectáreas, de las cuales 619 hectáreas (~67%) se encuentran al norte del departamento en los municipios de Roldanillo, La Unión y Toro, zona denominada distrito RUT. Esta zona del Valle del Cauca, correspondiente a una parte del valle geográfico del río Cauca, con una alta capacidad para producción agropecuaria, de tierras planas y fértiles, presenta ciertas limitaciones para su labrado debido a las variaciones del clima, con épocas de sequías e inundaciones, siendo estas últimas ocasionadas por el desbordamiento del río Cauca y sus tributarios (Sierra, 2008). Dichas inundaciones en el país, especialmente las presentadas en la segunda mitad del año 2010, y repetida en abril de 2011 por el denominado fenómeno de La Niña, generaron pérdidas parciales y totales de los cultivos de maracuyá amarillo en la zona (CORPOICA, 2011), pasando de tener el mejor rendimiento de cultivo de maracuyá del país en el 2010 con 35 ton/ha, a 8,6 ton/ ha para el año 2011 (DANE, 2012). Teniendo en cuenta el efecto de las inundaciones sobre el rendimiento del cultivo de maracuyá, es de gran importancia intensificar el estudio direccionado al diseño de estrategias alternativas para mantener el cultivo en condiciones ambientales adversas con objeto de evitar la pérdida total de la producción de maracuyá.
Uno de los pilares importantes para desarrollar este tipo de estrategias es el entendimiento de los mecanismos que presentan las plantas para responder a determinado estrés, siendo la activación del metabolismo anaerobio una de las respuestas ante el estrés por anegamiento. En el presente estudio se comparó la actividad de la enzima ADH de raíces de plántulas de maracuyá amarillo en condiciones de anegamiento y en condiciones normales de riego. Los resultados demuestran que la actividad de la enzima ADH en raíces de plántulas de maracuyá amarillo en condiciones de anegamiento difiere significativamente de las raíces en condiciones normales de riego. Este aumento significativo de la actividad de la enzima ADH en raíces de plántulas de maracuyá amarillo sometidas a condiciones de inundación, demuestran la presencia de una posible estrategia de sobrevivencia de las plántulas de maracuyá ante el anegamiento a corto plazo.
Materiales y métodos
Material vegetal y condiciones de crecimiento
Semillas de cultivo de maracuyá amarillo (Passiflora edulis var. flavicarpa) de la zona norte del departamento del Valle del Cauca fueron seleccionadas y proporcionadas por los agricultores del distrito de riego Roldanillo, La Unión y Toro (distrito RUT), en el valle geográfico del río Cauca. Estas se plantaron en bolsas de polietileno de 9 x 12" con sustrato de tierra enriquecido, tratado con fungicida TECTO 500 SC, y durante dos meses se mantuvieron con suficiente suministro de agua en el invernadero del Laboratorio de Suelos y Aguas (LASA) de la Universidad del Valle sede Meléndez. Después de los dos meses, se trasplantaron plántulas con altura entre 15 y 20 cm, en cubetas plásticas individuales de 12 litros, con las mismas condiciones de suelo y riego descritas anteriormente. Para el inicio del experimento se tomaron 18 plántulas de tres meses y medio desde que germinaron, y se seleccionaron aleatoriamente 9 de los contenedores para el tratamiento con inundación, condición que se logró mediante inundación del contenedor manteniendo la lámina de agua 2 cm por encima del nivel del suelo, y los otros 9 contenedores se mantuvieron en condiciones normales de riego. El experimento tuvo una duración de catorce días en condiciones semicontroladas en el invernadero del LASA, y para cada recipiente en condiciones de anegamiento se realizó el monitoreo del contenido de oxígeno disuelto (O2D) y temperatura (°C) del agua en cada unidad de tiempo, utilizando un oxímetro YSI 550A.
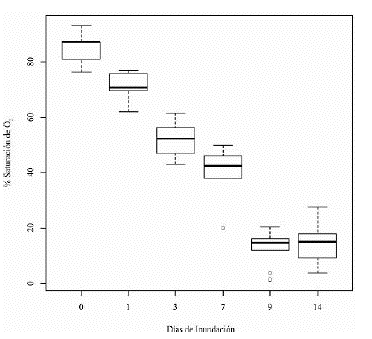
A partir de estas dos variables se determinó el porcentaje de saturación de O2 en el agua de los suelos del tratamiento de inundación, el cual mostró una disminución a lo largo del periodo del experimento (figura 1).
Toma de muestras
Se tomaron muestras de 2 cm de la zona apical de las raíces en los días 0, 1, 3, 7, 9 y 14 para ambos tratamientos. Las muestras de raíz de plántulas con tratamiento de inundación al día 14 fueron tomadas de raíces adventicias, menos de las plántulas designadas con los números 4 y 7, las cuales presentaron poca emergencia de dichas raíces (aproximadamente 0,3 cm de largo en promedio para cada plántula), en comparación con las demás que presentaron raíces adventicias de hasta 2 cm de largo. Las muestras fueron procesadas inmediatamente para el análisis bioquímico. Las muestras se tomaron de la zona apical de las raíces debido a su importancia en el metabolismo de la raíz y por su sensibilidad a la anoxia (Chen & Qualls, 2003).
Extracción y ensayo de la alcohol deshidrogenasa (ADH)
La extracción y determinación de la actividad enzimática de la ADH (Alcohol: NAD oxidorreductasa, E.C.1.1.1.1) se realizó como se describe en Fukao et al. (2006). La proteína cruda se extrajo a partir de aproximadamente 100 mg de tejido fresco de cada muestra, con un mortero pre-enfriado sobre hielo y 0,4 ml de buffer de extracción frío que contenía Tris-HCl (pH 9,0) 100 mM, MgCl2 20 mM y mercaptoetanol 0,1 % (v/v). Los extractos fueron centrifugados por 15 min a 13500 rpm y posteriormente la fracción sobrenadante se analizó para la actividad enzimática de la ADH. La mezcla de la reacción (1,2 ml) incluía 60 |i de extracto, Tris-HCl (pH 9,0) 50 mM y NAD 0,10 mM. El etanol (60 se añadió para iniciar la reacción, y la actividad de la ADH se determinó espectrofotométricamente a 340 nm por la reducción del NAD+ a temperatura ambiente por 2 min. Adicionalmente, se determinó la concentración de proteínas en los extractos crudos por el método de Bradford (1976), con BSA como estándar.
Diseño del estudio y análisis estadístico
El experimento realizado fue exploratorio, con el ánimo de evaluar las diferencias de la actividad de la enzima ADH, de raíces de tres plántulas de maracuyá amarillo, en condiciones de inundación en comparación con tres plántulas en condiciones normales (sin inundación). El experimento fue realizado por triplicado y se realizó un análisis descriptivo para observar el comportamiento de los datos de actividad enzimática de las plántulas en el tratamiento con inundación y sin inundación y a través del tiempo de duración del experimento (unidad de tiempo: día 0, 1, 3, 7, 9 y 14). Teniendo en cuenta las medidas de las variables de respuesta (actividad enzimática y actividad enzimática específica de la ADH) en diferentes unidades de tiempo del experimento sobre cada unidad experimental (plántula) para ambos tratamientos, el análisis se planteó como un diseño de medidas repetidas para así poder controlar el efecto de tiempos desiguales sobre el desarrollo de la respuesta en cada plántula para cada tratamiento. Para efectos del análisis estadístico, no se tomaron en cuenta las variaciones de las repeticiones del experimento ya que no presentaron diferencias significativas.
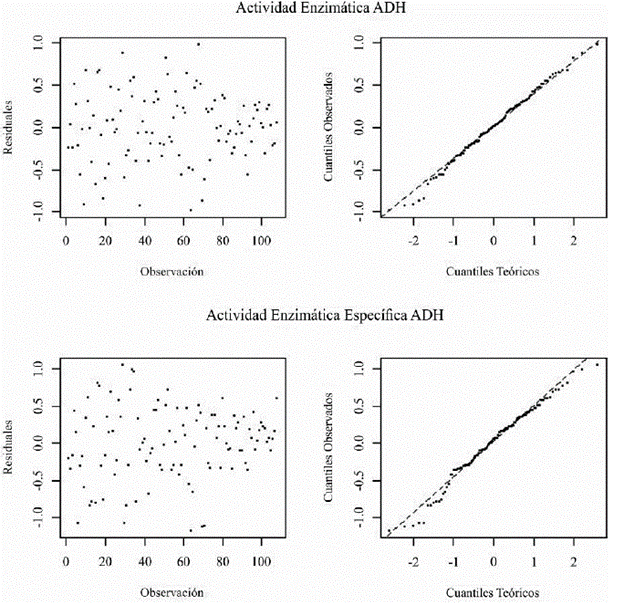
Los datos fueron transformados por logaritmo natural (LN) para ajustarlos a un modelo lineal mixto de coeficientes aleatorios, cuyos residuales tienen una distribución normal multivariada y existe una homogeneidad en la varianza para ambas variables de respuesta (figura 2). A partir de los efectos fijos de las mediciones repetidas se estimó el modelo lineal para ambas variables de respuesta a lo largo del tiempo. También, se realizó un análisis de contraste entre mediciones repetidas para determinar la significancia en las diferencias encontradas entre los tratamientos, las unidades de tiempo y la interacción para los datos de actividad enzimática y actividad enzimática específica de la ADH. Por último, se determinaron las diferencias entre los dos tratamientos en cada día mediante comparación de efectos principales, permitiendo precisar el significado del efecto de la interacción tiempo-tratamiento (Arnau & Bono, 2008). Para estos análisis estadísticos se utilizó el PROC MIXED de SAS 9.3 bajo la licencia de la escuela de estadística de la Universidad del Valle.
Resultados
La actividad enzimática de la ADH en raíces de plántulas de maracuyá amarillo aumentó rápidamente después del primer día de inundación hasta el noveno día. Al día 14, la respuesta de la actividad de la ADH disminuyó un poco. Sin embargo, esta seguía siendo mayor, en comparación con la estabilidad de la actividad en las plántulas en condiciones normales, sin inundación (figura 3).
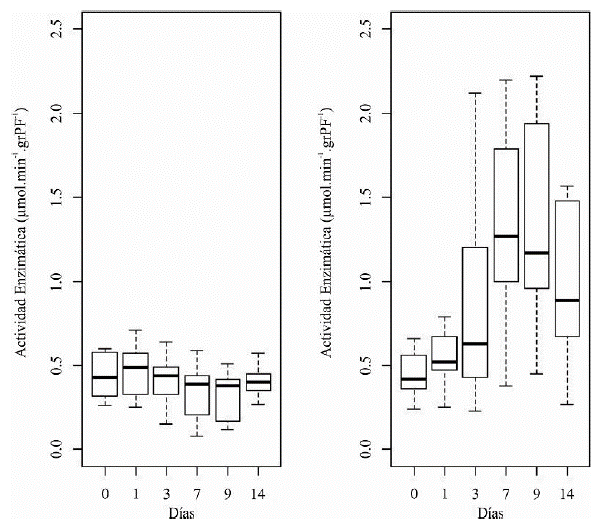
Figura 3 Diagrama de cajas de la actividad de la enzima ADH en las diferentes unidades de tiempo: Tratamiento sin inundación (izq.); Tratamiento con inundación (der.). grPF= gramo de Peso Fresco.
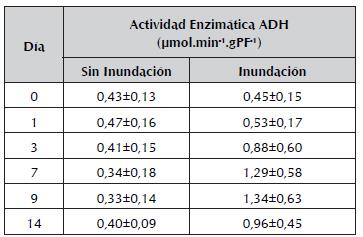
Después de 9 días de inundación, la actividad de la enzima ADH mostró el máximo incremento, de 3 veces respecto al tiempo 0 de inundación, y una alta variación de los datos. Por el contrario, en las plántulas sin inundar, la actividad de la enzima se mantuvo estable y con poca variación de los datos a lo largo del tiempo (tabla 1).
Tabla 1 Medias y desviación estándar de la actividad enzimática de la ADH de raíces de plántulas de maracuyá amarillo en cada tratamiento (Sin Inundación vs Inundación) por unidad de tiempo (día 0, 1, 3, 7, 9 y 14). Media ± desviación estándar (DS).
El análisis estadístico de los datos descritos en la figura 3 mostró que las diferencias encontradas en la respuesta de la enzima ADH a los tratamientos de inundación y sin inundación son significativas (P<0,1), al igual de las diferencias entre las unidades de tiempo y la interacción tratamiento-tiempo de la actividad enzimática (tabla 2).
Tabla 2 Comparación de los efectos fijos y su interacción para las variables actividad enzimática y actividad enzimática específica de la ADH de raíces de plántulas de maracuyá amarillo.
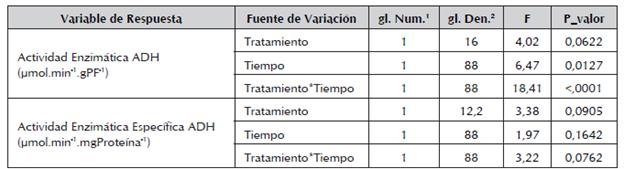
1Grados de Libertad del Numerador 2Grados de Libertad del Denominador
Al comparar los valores de la actividad enzimática entre los tratamientos en cada unidad de tiempo se demostró que las diferencias significativas (P<0,1) se dan a partir del día 3 hasta el día 14 de finalización del experimento (tabla 3).
Tabla 3 Diferencias entre los tratamientos con inundación y sin inundación en cada unidad de tiempo para las variables de respuesta actividad enzimática y actividad enzimática específica de la ADH de raíces de plántulas de maracuyá.
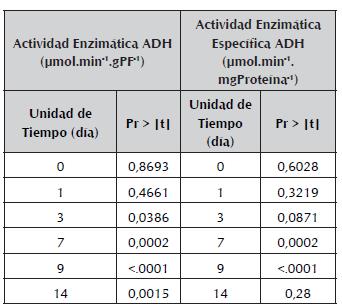
Teniendo en cuenta la respuesta de la actividad de la enzima en cada una de las plántulas del experimento, se encontró una respuesta variada a lo largo del tiempo en cada uno de los individuos de la población, y es de destacar el gran aumento de la actividad de la enzima en las raíces de las plántulas inundadas en comparación con las no inundadas, y por otro lado, la poca variación de la actividad de la enzima en las plántulas No. 4 y 7, que además presentaron diferencias de formación de raíces adventicias mostrando una velocidad de emergencia menor (menos de 0,05 cm/ día) con respecto al resto de las plántulas (aprox. 0,3 cm/día) en condiciones de inundación. La tendencia de la actividad de la enzima en la planta No. 7 es similar a la de las plantas en condiciones normales (sin inundación), y en la planta No. 4 tiende a aumentar la actividad de la enzima pero en menor proporción respecto al resto de plantas en condiciones de inundación (figura 4).
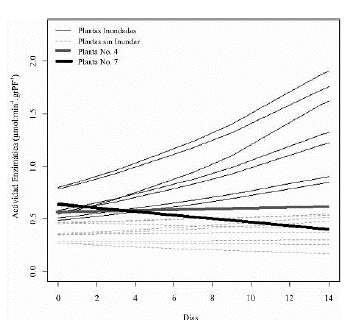
Figura 4 Tendencia de la actividad enzimática de la ADH de cada plántula de maracuyá amarillo a lo largo del tiempo.
El modelo encontrado finalmente para la actividad de la enzima alcohol deshidrogenasa a lo largo del tiempo en cada tratamiento (figura 5) muestra un patrón creciente en la respuesta de la actividad de la enzima en las plántulas de maracuyá amarillo por efecto del tratamiento inundación y del tiempo de exposición a la inundación, en contraste con el tratamiento sin inundación que no muestra una tendencia a aumentar sino a mantenerse a lo largo del tiempo.
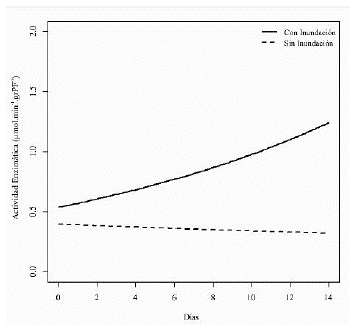
Figura 5 Estimación del modelo para la variable de respuesta actividad de la enzima alcohol deshidrogenasa en los tratamientos inundación y sin inundación a lo largo del tiempo.
Por otro lado, tenemos que la actividad enzimática específica de la ADH muestra una tendencia a ser mayor en plántulas sometidas a inundación en comparación con las que se encuentran en condiciones normales, pero no se visualiza un efecto marcado a lo largo del tiempo (figura 6).
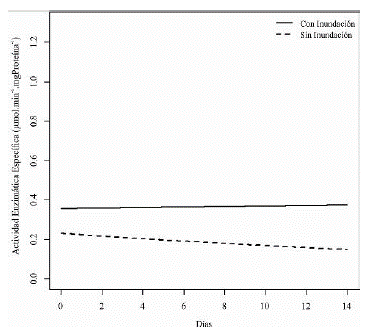
Figura 6 Estimación del modelo para la variable de respuesta actividad enzimática específica alcohol deshidrogenasa en los tratamientos inundación y sin inundación a lo largo del tiempo.
A pesar de que el contraste entre las mediciones muestra estas diferencias en el tratamiento pero no en el tiempo para la actividad específica, se encontró que hay un efecto de la interacción entre el tratamiento y la unidad de tiempo (tabla 2). El análisis de las diferencias de la interacción tratamiento-tiempo se observan en la tabla 3, donde se demuestra que existen diferencias significativas entre los tratamiento con inundación y sin inundación a partir del día 3 hasta el día 9 del experimento.
Discusión
La baja disponibilidad de oxígeno es un componente muy importante en la respuesta metabólica de las plantas ante el estrés por anegamiento. Estudios sobre el efecto de la inundación en la respuesta de la actividad de las enzimas del metabolismo anaerobio como la ADH sugieren esta respuesta como un mecanismo de tolerancia al estrés debido al déficit de oxígeno (Agarwal et al., 2007). La tendencia de aumento encontrada en la actividad de la enzima ADH de las raíces de plántulas de maracuyá amarillo ante el anegamiento, comparado con la actividad en condiciones normales, proporciona la evidencia de que la fermentación alcohólica en estas plantas se activa, como un mecanismo para la producción de energía en condiciones de estrés por la ausencia del oxígeno. La razón por la cual la fermentación alcohólica es de gran importancia durante la inundación es que en este proceso ocurre la regeneración continua de NADH en el citoplasma (Kimmerer, 1987). Por tanto el proceso de glicólisis continúa y se genera energía en forma de ATP para el mantenimiento del metabolismo de las raíces para la supervivencia de las plantas, ya que en ausencia de oxígeno (en condiciones anóxicas), el ciclo de Krebs y la fosforilación oxidativa dependiente de la disponibilidad de oxígeno se bloquean (Hossain & Nasir, 2011).
A pesar de que se muestra una mayor actividad de la enzima ADH de las raíces de las plántulas de maracuyá en condiciones de anegamiento respecto a las condiciones normales de riego, se encontró que a lo largo del tiempo la actividad de la enzima aumenta solamente después del primer día hasta los 9 días de inundación y luego disminuye. La poca actividad enzimática durante el primer día de inundación posiblemente se debió a la disponibilidad de oxígeno aún presente durante ese periodo de tiempo, ya que al tener en cuenta los valores del porcentaje de saturación de O2 del agua de inundación (figura 1), se encontró que: según el estatuto de calidad del agua introducido por Dunnette desde 1979, el porcentaje de saturación de O2 para el día 0 indica un buen entorno (75-89%), y para el día 1 indica una limitada disponibilidad de oxígeno (60-74%), mostrando que es menor respecto al día 0 pero aún está disponible para la planta. A partir del día 3 se empieza a tener un medio con escasa saturación de oxígeno (<60%) hasta el final del experimento al día 14, lo cual corresponde a la rápida inducción de la actividad de la enzima a partir de este día (3) hasta el día noveno. Se encontraron diferentes posibles causas por las cuales la actividad de la enzima en el día 14 muestra una menor actividad respecto al noveno día. El hecho de que las muestras de este día hayan sido tomadas de raíces adventicias y no de raíces subterráneas influye, ya que las raíces adventicias se encuentran cerca de la superficie del agua donde hay mayor disponibilidad de oxígeno disuelto, teniendo en cuenta que la cantidad de oxígeno disuelto en el agua es inversamente proporcional a la profundidad (Cifuentes & Torres, 2003). Por otro lado, existe la posibilidad, de que estas raíces adventicias generadas por las plántulas presenten espacios aéreos (aerénquima) que según el estudio de Escobar (2013) se generan en las raíces adventicias de maracuyá amarillo en condiciones de anegamiento, permitiendo de esta manera hacer uso del oxígeno disponible tanto de la superficie del agua como del oxígeno almacenado en los espacios aéreos para activar su metabolismo normal y disminuir la dependencia de obtención de energía por la vía fermentativa. Otra de las posibles razones por las cuales la actividad de la enzima fue menor al día 14, es el tiempo de exposición a las condiciones de anegamiento, donde estudios en Arabidopsis han mostrado que la actividad de la ADH es necesaria para la resistencia al anegamiento a corto plazo (Ellis et al., 1999), es decir que su activación es importante solamente los primeros días de inundación. Comparando estos estudios con la respuesta de la actividad enzimática del ADH en maracuyá, se puede demostrar que este mecanismo de rescate por medio de la activación del metabolismo anaeróbico es de gran importancia para su supervivencia durante los primeros 9 días de inundación. Posiblemente, la inducción de la ADH para periodos de inundación mayores a 9 días no sea por sí sola la causa de supervivencia de las plántulas de maracuyá durante el anegamiento.
La respuesta de la actividad de la enzima ADH presentó una gran variación entre los individuos en el tratamiento de inundación, encontrando tanto plántulas que aumentaron significativamente su actividad enzimática, así como plántulas "como las numeradas 4 y 7" que presentaron la peculiaridad de manifestar una poca activación de la enzima respecto al resto de las plantas inundadas, y que de hecho no generaron raíces adventicias a la misma velocidad que el resto de las plántulas del experimento con tratamiento de inundación. Estas variaciones encontradas dentro de la población podrían sugerir una variabilidad genética que esté generando este comportamiento de la actividad de la enzima ADH en los diferentes individuos. Por ejemplo, la característica observada en las plántulas No. 4 y 7 podrían relacionarse con la variación de un factor de inducción que afecta ambos atributos en las plantas (la actividad de la enzima ADH y la formación de raíces adventicias), donde la expresión de un gen puede influenciar y finalmente modular la respuesta a la inundación, en diferentes características incluyendo la respuesta de la fermentación etanólica (Fukao et al., 2006), siendo esto una aproximación teórica de lo que podría estar sucediendo a nivel molecular para que se presenten las variaciones observadas en las plántulas de maracuyá amarillo en condiciones de inundación.
Este estudio provee evidencias de un posible mecanismo de sobrevivencia de plántulas de maracuyá amarillo a las condiciones de hipoxia o anoxia a corto plazo, consecuencia de la inundación, al activar la enzima ADH implicada en el metabolismo anaerobio, donde las plántulas anegadas presentaron una actividad enzimática de la ADH hasta tres veces mayor en comparación a las plántulas en condiciones normales de riego. Se propone la exploración de otros rasgos como son la presencia de barreras para evitar la pérdida radial de oxígeno y mecanismos de regulación y señalización a nivel molecular responsables de desencadenar respuestas consideradas de tolerancia a la inundación, lo cual será de relevancia para el conocimiento del comportamiento del cultivo de maracuyá en condiciones de anegamiento.
Agradecimientos
Especialmente a la profesora Neyla Benítez directora del Laboratorio de Investigaciones Microbiológicas (LIM), por facilitar el espacio y material para la realización de los experimentos, al estadístico Diego Tovar por su valiosa ayuda en el análisis estadístico, y a los cultivadores del distrito RUT quienes proporcionaron las semillas. El presente trabajo se logró realizar en el marco del proyecto "Evaluación de los cambios en la Relación Agua-Suelo-Planta-Atmosfera (RASPA), en condiciones de anegamiento en la IFAN. Aplicado al caso Maracuyá - Passiflora edulis, post inundaciones 2010". Agradecimiento a la Universidad del Valle que financió este proyecto y brindó infraestructura para desarrollar los diferentes procesos de este trabajo.
REFERENCIAS
Agarwal, S., A. Kapoor, O. S. Lakshmi & Grover, A. (2007). Production and phenotypic analysis of rice transgenics with altered levels of pyruvate decarboxylase and alcohol dehydrogenase proteins. Plant Physiology and Biochemistry, 45, 637-646. [ Links ]
Armstrong, W., D. Cousins, J. Armstrong, D. W. Turner & Beckett, P. M. (2000). Oxygen distribution in wetland plant roots and permeability barriers to gas-exchange with the rhizosphere: a microelectrode and modelling study with Phragmites australis. Annals of Botany, 86, 687-703. [ Links ]
Arnau, J. & Bono, R. (2008). Estudios longitudinales de medidas repetidas: Modelos de diseño y análisis. Escritos de Psicología, 2(1), 32-41. [ Links ]
Borella, J., Amarante, L., Colares, D., Barneche, A. C. & Bolacel, E. J. (2014). Waterlogging-induced changes in fermentative metabolism in roots and nodules of soybean genotypes. Sci. Agric ., 71(6), 499-508. [ Links ]
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 72, 248-254. [ Links ]
Chen, H. & Qualls, R. G. (2003). Anaerobic metabolism in the roots of seedlings of the invasive exotic Lepidium latifolium. Environmental and Experimental Botany, 50, 29-40. [ Links ]
Cifuentes, J. & Torres, M. (2003). El océano y sus recursos II. Las ciencias del mar. México: Fondo de Cultura Económica. [ Links ]
CORPOICA (Corporación Colombiana de Investigación Agropecuaria). (2011). Estrategias de respuesta de CORPOICA para mitigar el impacto de las inundaciones sobre la agricultura colombiana. Rehabilitación de la capacidad productiva de los suelos afectados por las inundaciones, en el Valle del Cauca. Ministerio de Agricultura y Desarrollo Rural. Recuperado de http://www.palmira.gov.co/potpaigr/02.%20INFORMACION%20REGIONAL/Documentos%20consulta/Corpoica_Norte_del_Valle.pdf [ Links ]
DANE (Departamento Administrativo Nacional de Estadística). (2012). Insumos y factores asociados a la producción agropecuaria. Pasifloráceas en Colombia. Ministerio de Agricultura y Desarrollo Rural. República de Colombia. Recuperado de https://www.dane.gov.co/files/investigaciones/agropecuario/sipsa/insumos_factores_de_produccion_noviembre_2012.pdf [ Links ]
Dennis, E. S., Dolferus, R., Ellis, M., Rahman, M., Wu, Y., Hoeren, F. U., Grover, A., Ismond, K. P., Good, A. G. & Peacock, W.J . (2000). Molecular strategies for improving waterlogging tolerance in plants. Journal of Experimental Botany, 51 (342), 89-97. [ Links ]
Dunnette, D. A. (1979). A geographically variable water quality index used in Oregon. Journal of the Water Pollution Control Federation, 51 (1), 53-61. [ Links ]
Ellis, M. H., Dennis, E. S. & Peacock, W. J. (1999). Arabidopsis roots and shoots have different mechanisms for hypoxic stress tolerance. Plant Physiology, 119, 57-64. [ Links ]
Escobar, J. S. (2013). Características morfológicas y anatómicas de maracuyá (Passiflora edulis) sometida a anegamiento. Universidad del Valle, Cali, Colombia. [ Links ]
Fukao, T., Kennedy, R. A., Yamasue, Y. & Rumpho, M. E. (2003). Genetic and biochemical analysis of anaerobically induced en zymes during seed germination of Echinochloa crus-galli varieties tolerant and intolerant of anoxia. Journal of Experimental Botany, 54 (386), 1421-1429. [ Links ]
Fukao, T., Xu, K., Ronald P. C. & Bailey-Serres, J. (2006). A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. The Plant Cell, 18, 2021-2034. [ Links ]
Gaut, B. S. & Clegg, M. T. (1991). Molecular evolution of alcohol dehydrogenase 1 in members of the grass family. Proc. Natl. Acad. Sci. USA, 88, 2060-2064. [ Links ]
Hossain, A. & Nasir, S. (2011). Mechanisms of waterlogging tolerance in wheat: Morphological and metabolic adaptations under hypoxia or anoxia. Australian Journal of Crop Science, 5(9), 1094-1101. [ Links ]
Huang, Y., Picha, D. H. & Kilili, A. W. (2002). Atmospheric oxygen level influences alcohol dehydrogenase and pyruvate decar-boxylase activities in sweetpotato roots. J. Plant Physiol ., 159, 129-136. [ Links ]
Johnson, J. R., Cobb, C. & Drew, M. C. (1994). Hypoxic induction of anoxia tolerance in roots of Adh1 Null Zea mays. Plant Physiol ., 105, 61-67. [ Links ]
Joshi R. & Kumar P. (2012). Lysigenous aerenchyma formation involves non-apoptotic programmed cell death in rice (Oryza sativa L.) roots. Physiol Mol Biol Plants, 18(1), 1-9. [ Links ]
Kimmerer, T. W. (1987). Alcohol dehydrogenase and pyruvate de-carboxylase activity in leaves and roots of eastern cottonwood (Populus deltoids Bartr.) and soybean (Glycine max L.). Plant Physiol ., 84, 1210-1213. [ Links ]
Kulkarni, S. S. & Chavan, P. D. (2013). Study of some aspects of anaerobic metabolism in roots of finger miller and rice plants subjected to waterlogging stress. International Journal of Botany, 2013, 9 (2),1-6. [ Links ]
Kumutha, D., Sairam, R.K., Ezhilmathi, K., Chinnusamy, V. & Meena, R.C. (2008). Effect of waterlogging on carbohydrate metabolism in pigeon pea (Cajanus cajan L.): Upregulation of sucrose synthase and alcohol dehydrogenase. Plant Science, 175, 706-716. [ Links ]
Medina, C., Sanches, M. C., Tucci, M. L., Sousa, A. F., Cuzzuol, G. R. & Joly, C. A. (2009). Erythrina speciosa (Leguminosae-Papilionoideae) under soil water saturation: morphophysiological and growth responses. Annals of Botany, 104, 671-680. [ Links ]
Menezes, M. A., Donizeti, J. & Mota, L. E. (1995). Anaerobic metabolism of Euterpe oleracea. II plant tolerance mechanism to anoxia. R. Bras. Fisiol. Veg ., 7(1), 47-51. [ Links ]
Nishiuchi, S., Yamauchi, T., Takahashi, H., Kotula, L. & Nakazono, M. (2012). Mechanisms for coping with submergence and waterlogging in rice. Rice, 5(2), 1-14. [ Links ]
Oliveira, H. C., Freschi, L. & Sodek, L. (2013). Nitrogen metabolism and translocation in soybean plants subjected to root oxygen deficiency. Plant Physiology and Biochemistry, 66, 141-149. [ Links ]
Oluoha, U. (1995). Purification and kinetic properties of alcohol dehydrogenase from yellow yam tubers (Dioscorea cayenensis). Plant Science, 107, 1-7. [ Links ]
Potosí, C. T., Espinosa, F. & Guevara, P. C. (2008). Comportamiento agronómico de maracuyá Amarillo Passiflora edulis var. flavicarpa en condiciones ambientales de sabana inundable, municipio de Arauca. Universidad Nacional de Colombia, Sede Orinoquia, Arauca, Colombia. [ Links ]
Roberts, J. K. M., Callis, J., Jardetzky, O., Walbot, V. & Freeling, M. (1984). Cytoplasmic acidosis as a determinant of flooding intolerance in plants. Proc. Natl. Acad. Sci. USA, 81 , 6029-6033. [ Links ]
Roorkiwal, M. & Chand, P. (2012). Sequence similarity based identification of abiotic stress responsive genes in chickpea. Bioinformation, 8 (2), 092-097. [ Links ]
Sachs, M. M., Freeling, M. & Okimoto, R. (1980). The anaerobic proteins of maize. Cell, 20, 761-767. [ Links ]
Sairam, R. K., Kumutha, D., Ezhilmathi, K., Deshmukh, P. S. & Srivastava, G. C. (2008). Physiology and biochemistry of waterlogging tolerance in plants. Biologia Plantarum, 52(3), 401-412. [ Links ]
Sierra, J. A. (2008, Marzo). Embalse de La Salvajina. Inundaciones otra vez. Procaña. Recuperado de http://issuu.com/procana.org/docs/4_ded7b0d6224fff [ Links ]
Voesenek, L. A. C. J., Colmer, T. D., Pierik, R., Millenaar, F. F. & Peeters, A. J. M. (2006). How plants cope with complete submergence. New Phytologist, 170, 213-226. [ Links ]
Wignarajah, K., Greenway, H. & John, C. D. (1976). Effect of waterlogging on growth and activity of alcohol dehydrogenase in barley and rice. New Phytol ., 77, 585-592. [ Links ]
Recibido: 18 de Enero de 2015; Aprobado: 28 de Septiembre de 2015
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
