










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Biotecnología
Print version ISSN 0123-3475
Rev. colomb. biotecnol vol.18 no.1 Bogotá Jan./June 2016
https://doi.org/10.15446/rev.colomb.biote.v18n1.50274
DOI: http://dx.doi.org/10.15446/rev.colomb.biote.v18n1.50274
ARTÍCULO DE INVESTIGACIÓN
Comparación del potencial oxidativo de Acidithiobacillus ferrooxidans, en un proceso de biodesulfurización de carbón
Comparison of the oxidative potential of Acidithiobacillus ferrooxidans, in biodesulphurization process of coal
María Prada Fonseca*, Gerardo Caicedo Pineda**, Marco Márquez Godoy***
* MSc. Ingeniería de Materiales y Procesos, Universidad Nacional de Colombia -Sede Medellín, Colombia. Calle 51 # 78 A - 73, mcprada@unal.edu.co
** PhD. Ingeniería ciencia y tecnología de los materiales, Universidad Nacional de Colombia - Sede Medellín, Colombia. Calle 10 A # 43D -96, gacaiced@unal.edu.co
*** PhD. Mineralogía, Universidad Nacional de Colombia - Sede Medellín, Colombia. Carrera 80 # 65 -223, mmarquez@unal.edu.co
Recibido: septiembre 16 de 2015 Aprobado: abril 26 de 2016
Resumen
En este estudio se comparó la actividad oxidativa de dos cepas de Acidithiobacillus ferrooxidans en un proceso de desulfurización, empleando un carbón sub-bituminoso alto en azufre (2.30% de azufre total, con 1.06% pirítico, 1.10% orgánico y 0.14% de sulfatos), proveniente de la mina "La Guacamaya" ubicada en Puerto Libertador -Córdoba, Colombia. Se realizaron ensayos a nivel de erlenmeyer, la concentración de hierro total en la solución fue de 200 mg/L y 1200 mg/L respectivamente, empleando sulfato ferroso. El proceso fue monitoreado mediante mediciones periódicas de los principales factores físico-químicos implicados (pH, potencial de óxido-reducción (Eh), hierro en solución y concentración celular). Según los resultados obtenidos, la mayor eficiencia del proceso se logró, al trabajar con microorganismos compatibles con Acidithiobacillus ferrooxidans y una concentración de sulfato ferroso inicial de 1200 mg/L, el cual obtuvo la mayor tasa de oxidación de pirita (Py oxidada) (68%), así como, mejores condiciones en el medio lixiviante (pH: 1,47; Eh: 625 mV; 6.3×108 células/mL) en comparación con el cultivo axénico a las mismas condiciones (Py oxidada: 52%, pH: 1,63, Eh: 580 mV, 5.1×108 células/mL), después de 12 días de experimentación.
Palabras clave: bacteria, pirita, biolixiviación, sulfato ferroso.
Abstract
This study aimed comparing the oxidative activity of two strains of Acidithiobacillus ferrooxidans in a desulphurization process, using a sub-bituminous coal with a high sulfur content (2.30% total sulfur: 1.06% as pyritic, 1.10% as organic and 0.14% from sulfates) from "La Guacamaya" mine, located in Puerto Libertador - Cordoba, Colombia. Several assays were performed in Erlenmeyer flasks, the total iron concentration used in solution were 200 mg/L and 1200 mg/L respectively, using ferrous sulfate. The process was monitored by periodically measuring the main physicochemical factors involved (pH, Eh, cell population and iron in solution). According to the results obtained, the highest efficiency of the process was achieved by working with microorganisms compatible with Acidithiobacillus ferrooxidans and initial concentration of 1200 mg/l of ferrous sulfate, which had higher pyrite oxidation rates (Py oxidized) up to 68% and the best experimental conditions in the leaching medium (pH: 1,47; Eh: 625 mV; 6.3×108 cells/mL), in comparison with the axenic culture on the same conditions (Py oxidized: 52%; pH: 1,63; Eh: 580 mV; 5.1×108cells/mL), after 12 days of experimentation.
Key words: bacteria, pyrite, bioleaching, ferrous sulfate.
Introducción
A nivel mundial, el carbón colombiano es reconocido por tener bajo contenido de cenizas, azufre, ser alto en volátiles y tener un alto valor calorífico (UPME, 2007). No obstante, en algunas regiones la cantidad de algunos componentes presentes en este combustible, tales como el azufre (orgánico e inorgánico) le restan valor y obligan a la aplicación de tratamientos para su utilización (León, 2006).
El dióxido de azufre (SO2) es uno de los gases más tóxicos que contaminan el ambiente, formado principalmente como un subproducto de la combustión del azufre (Chou, 2012). La necesidad de obtener carbones con bajo contenido de este elemento a partir de carbones de baja calidad, ha dado lugar a una gran cantidad de métodos de extracción de azufre basados en técnicas físicas y químicas, pero su utilización requiere altos capitales de inversión, genera residuos y efluentes contaminantes (Prayuenyong, 2002). Por esta razón, los métodos biológicos han despertado gran interés, debido no solo a que presenta altas eficiencias para extraer pirita del carbón, sino porque requiere bajos capitales de inversión y sus subproductos no deterioran el ambiente (Caicedo, 2008).
Los procesos de biolixiviación se basan fundamentalmente en el uso de microorganismos ácidofilos, capaces de prosperar y oxidar sulfuros metálicos insolubles tales como la pirita, en un medio acuoso, con el fin de lixiviar el azufre en forma de sulfatos (Aller et al., 2001; Cara et al., 2005; Harneit et al., 2006). Estudios previos sobre el comportamiento bacteriano en presencia de azufre inorgánico han sido en su mayoría llevados a cabo empleando Acidithiobacillus ferrooxidans (Johnson & Hallberg, 2003). Los resultados de estos estudios son variables debido a las condiciones del medio lixiviante. Por ejemplo, (Caicedo & Marquéz, 2011) obtuvieron 50% de oxidación de azufre piritico, empleando concentraciones de hierro iniciales menores a 1200 mg/L y un cultivo celular mixto (Acidithiobacillus ferrooxidans y Acidithiobacillus thiooxidans). Por otra parte, (Cardona, 2008) alcanzó una oxidación de pirita del 85% empleando microorganismos compatibles con Acidithiobacillus ferrooxidans y 95% con un cultivo mixto (Acidithiobacillus ferrooxidans y Acidithiobacillus thiooxidans), variando el tamaño de partícula del carbón y usando una concentración de hierro ferroso de 1000 mg/L. Finalmente, (Peláez et al., 2013) alcanzaron un 74% de oxidación de pirita empleando una relación inicial de iones Fe3+/Fe2+ y una concentración de hierro total de 1200 mg/L, en un proceso de biodesulfurización de carbón empleando un cultivo axénico de Acidithiobacillus ferrooxidans.
Si bien una serie de bacterias han sido descritas como eficientes en los procesos de biooxidación de pirita del carbón, es poco lo que se sabe sobre su comportamiento en ambientes complejos, en particular, la influencia de la actividad oxidativa de las bacterias en presencia de diferentes concentraciones de hierro ferroso total, como única fuente soluble de energía disponible. El propósito de esta investigación es comparar la actividad oxidativa de un cultivo compatible con Acidithiobacillus ferrooxidans vs. una cepa axénica de Acidithiobacillus ferrooxidans, a nivel de laboratorio, al variar la concentración inicial de hierro ferroso en el sistema, y determinar si los iones Fe2+ adicionados aumentan la eficiencia del proceso de biodesulfurización de un carbón proveniente del municipio de Puerto Libertador - Córdoba, Colombia, es decir, si se incrementa la remoción de azufre inorgánico presente en el carbón.
Materiales y métodos
Tipo de muestra y preparación del carbón
La muestra de carbón fue tomada de la mina "La Guacamaya" (Puerto Libertador, Córdoba-Colombia). A esta se le realizaron cuarteos sucesivos y se le redujo el tamaño de grano -60# Tyler (0.246 mm = 246 µm), empleando un molino de bolas. Con el fin de evaluar las propiedades de la muestra tomada, se realizaron análisis de las formas de azufre presentes en el carbón, bajo la norma ASTM D 2492-02, siguiendo los pasos descritos para la medición de azufre total ASTM 4239-14. Así como análisis próximos a la muestra según las normas ASTM D 3173-11 (Humedad residual), ASTM D 3174 (cenizas), ISO 562 (materia volátil), ASTM D 3172-13 (carbono fijo) y ASTM D 5865 (Capacidad calorífica bruta).
Microorganismos y medios
Del cepario del laboratorio de biominería y biohidrometalurgía de la Universidad Nacional de Colombia, Sede Medellín, se tomó un aislado compatible con Acidithiobacillus ferrooxidans proveniente del depósito de carbón de la mina "La Angostura" (Caicedo, 2008; Cardona, 2008), para corroborar la compatibilidad de la cepa escogida, se realizó una identificación bioquímica, descrita en el manual de Bergey Staley et al.(1994); y un cepa axénica de Acidithiobacillus ferrooxidans (American Type Culture Collection 23270), conservados a 4 °C ± 1 °C, las cuales tuvieron una adaptación previa al proceso de biodesulfurización de carbón, bajo un protocolo preestablecido (Caicedo, 2008).
Biodepiritización del carbón
La preparación de los ensayos se hizo en erlenmeyers de 500 mL, a un volumen de 200 mL, empleando el medio de cultivo T&K modificado (0.5 g/L de (NH4)3SO4, 0.5 g/L de KH2PO4, 0.5 g/L de MgSO4 y FeSO4·7H2O al 33.33 % p/v), 10 % p/v de carbón y 10% v/v de inoculo (107 - 108 células/mL). El pH del medio se ajustó en 1,5 con ácido sulfúrico (HSSO4) 1N. Las variables evaluadas fueron: (i) concentración inicial de hierro en solución: 200 mg/L (C1) y 1200 mg/L (C2), mediante la adición de sulfato ferroso; (ii) cultivo bacteriano: microorganismos compatibles con Acidithiobacillus ferrooxidans (N) y la cepa ATCC 23270 (P). Los ensayos resultantes fueron: NC1, PC1, NC2, PC2. Cada ensayo se preparó por duplicado más un control negativo (abiótico) con medio nutritivo en condiciones estériles. Los experimentos fueron incubados a 30 °C ± 1 °C, con agitación continua a 180 rpm ± 2 rpm durante 12 días, tiempo en el cual los microorganismos alcanzaron la fase estacionaria.
Procedimientos analíticos
El monitoreo de los experimentos se realizó a partir de mediciones de Eh, empleando un equipo SCHOTT Handylab con un electrodo ORP Schott PT6880 de Ag/AgCl, y pH, utilizando equipo HACH HQ40d, con un electrodo intelliCALTM pHC301. El recuento celular se realizó en una cámara de Neubaüer. Adicionalmente, las suspensiones fueron analizadas en un espectrofotómetro marca GENESYS™ 10 UV de barrido, por el método colorimétrico o-fenantrolina (ASTM E394 - 09).
Resultados
Análisis químicos y mineralógicos de la muestra
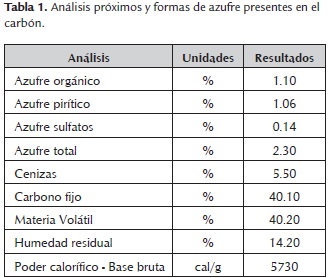
La tabla 1 muestra los análisis próximos y de las formas de azufre presentes en el carbón. Este presentó un contenido de azufre total de 2.30%, del cual 1.06% corresponde a azufre pirítico. Mediante la aplicación de la norma ASTM D388 -12, para la clasificación de carbones por rango, se determinó que el carbón empleado es de tipo sub - bituminoso A.
Biodepiritización del carbón
Mediciones de pH y potencial redox (Eh)
El comportamiento del pH vs. Tiempo se muestra en la figura 1. En general, los primeros dos días del proceso se evidenció un aumento en los valores de pH, los bioensayos PC1 y PC2 alcanzaron mayores valores de pH (2,10 y 2,20 respectivamente), frente a los experimentos NC1: 1,90 y NC2: 2,05. Vale la pena mencionar que al adicionar una mayor concentración de sulfato ferroso inicial (1200 mg/L), se observaron mayores valores de pH, los primeros días. Entre el día dos y el día cuatro, la curva presenta una caída en los valores de todos los ensayos, especialmente en aquellos en los cuales se empleó una mayor concentración de sulfato ferroso inicial (1200 mg/L), este comportamiento se evidenció tanto para el cultivo axénico (PC2: 1,80), como para el aislado bacteriano compatible con Acidithiobacillus ferrooxidans (NC2: 1,76), tendencia que se mantuvo hasta finalizar el proceso. Vale la pena mencionar, que los ensayos en los cuales se utilizó el aislado bacteriano compatible con Acidithiobacillus ferrooxidans, presentaron los menores valores de pH en las dos concentraciones de sulfato ferroso estudiadas (NC1: 1,52 y NC2: 1,47), después de 12 días de experimentación.
Por otra parte, los controles presentaron un aumento en los valores de pH los primeros dos días del proceso, sin embargo después del día dos, el pH se mantuvo sin cambios, finalizando el proceso con un pH cercano al inicial.
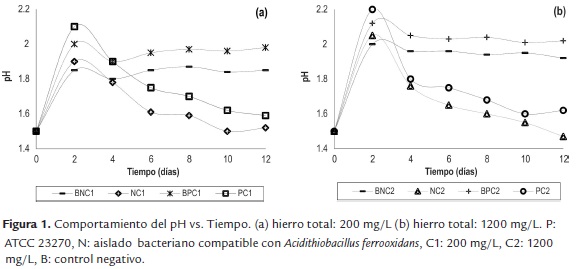
La figura 2 muestra el comportamiento del Eh vs. Tiempo. A nivel general, todos los bioensayos presentaron un continuo aumento en los valores del Eh a lo largo del proceso, alcanzando valores por encima de 560 mV. Cabe mencionar que los ensayos con mayor concentración de hierro total (1200mg/L) empezaron el proceso con menores valores de Eh (NC2: 350 mV PC2: 300 mV) en comparación con los ensayos NC1 y PC1 (390 mV y 400 mV respectivamente). Adicionalmente, se observó una pequeña caída en la velocidad de incremento del Eh (día 0 - 2) en los experimentos a los cuales se adicionó 1200 mg/L, siendo que esta tendencia se invirtió después del día cuatro, a partir del cual, estos ensayos mostraron altos valores de Eh. Vale la pena resaltar que tanto NC1 como NC2, alcanzaron los mayores valores de Eh (610 mV y 625 mV, respectivamente) en comparación con los ensayos PC1 (560 mV) y PC2 (580 mV) en los cuales se empleó la cepa ATCC 23270. Por otra parte, los ensayos abióticos se mantuvieron con una tendencia constante, alrededor de 400 mV en comparación con los ensayos bióticos durante el proceso.
Crecimiento bacteriano
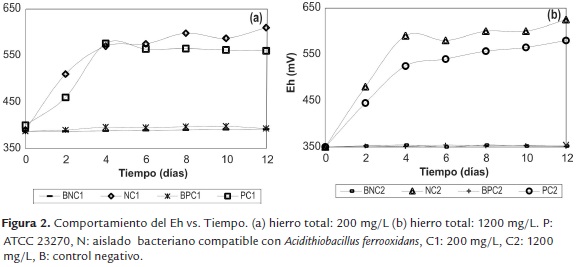
La figura 3 muestra el logaritmo de la concentración celular vs. tiempo. En general, se observó un comportamiento similar, caracterizado por una fase lag los primeros dos días del proceso, seguido por un crecimiento exponencial o fase logarítmica, entre el día dos y el día cuatro, alcanzando posteriormente la fase estacionaria. En este sentido, los experimentos en los cuales se empleó una mayor concentración de sulfato ferroso inicial (NC2 y PC2) presentaron menor velocidad de crecimiento celular durante los primeros dos días del proceso, en relación con sus contrapartes de menor concentración (NC1 y PC1), lo cual es coherente con el comportamiento disímil del pH y Eh en el mismo periodo, siendo que después de esto, el ensayo NC2 alcanzó mayores tasas de crecimiento celular (6.3×108 células/mL) en comparación con el cultivo axénico a las mismas condiciones (5.1×108 células/mL).
Hierro removido y pirita oxidada
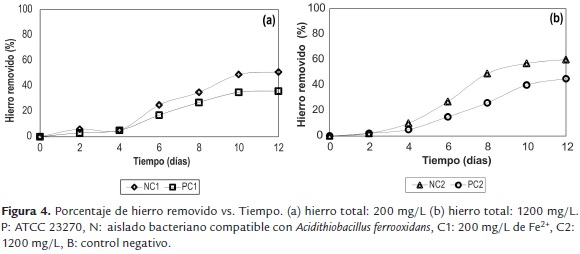
La figura 4 muestra el cambio en la concentración de hierro en solución en el tiempo, para los tratamientos estudiados. Todos los ensayos presentaron una solubilización significativa de hierro a partir del cuarto día. Después de doce días de experimentación, los ensayos que utilizaron cepas nativas, NC1 (51.34%) y NC2 (60.45%), presentaron mayores porcentajes de lixiviación, en comparación con sus análogos en los cuales se empleó el cultivo axénico PC1 y PC2 (36.51% y 45.27% respectivamente).
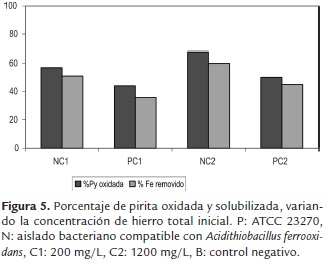
La figura 5 muestra el histograma de pirita oxidada al finalizar el proceso. Se puede observar que los experimentos en los cuales se empleó el aislado bacteriano compatible con Acidithiobacillus ferrooxidans y una concentración de hierro inicial de 1200 mg/L, presentaron un mayor potencial oxidativo y obtuvieron los mejores resultados (NC2: 68%); en comparación con los ensayos en los cuales se utilizó el cultivo axénico ATCC 23270 (PC2: 52%) a las mismas condiciones. De manera análoga, los experimentos con bacterias compatibles con Acidithiobacillus ferrooxidans y menores concentraciones de hierro ferroso inicial (200 mg/L) presentaron mayores porcentajes de pirita oxidada (NC1: 57%), mientras su contraparte PC1 solo alcanzó un 44% de oxidación del sulfuro metálico.
Discusión
Sulfato ferroso y la actividad oxidativa de Acidithiobacillus ferrooxidans
Según el proceso de biolixiviación de pirita en los cuales se emplea Acidithiobacillus ferrooxidans propuesto por algunos autores (Daoud & Karamanev, 2006; Kargi, 1986; Sand et al., 2001); el ácido (H+) es considerado un factor necesario para la (re)generación del ion férrico (Fe3+), responsable de la disolución de pirita. De esta manera, el mayor incremento en los valores de pH (figura 1) observado en los ensayos con mayores concentraciones de hierro (NC2 y PC2) pudo ser debido a un mayor consumo de iones H+ (2H+ por cada Fe2+) (Blázquez et al., 1993). Sin embargo, los ensayos abióticos también evidenciaron un aumento en los valores de pH durante este periodo, indicando que los agentes alcalinizantes presentes en la matriz del carbón también podrían influir sobre este efecto, lo cual coincide con lo reportado en investigaciones previas realizadas sobre carbones de la zona (Caicedo & Marquéz, 2011; Peláez et al., 2013).
La posterior disminución de los valores de pH que se observó a partir del día dos, es una evidencia de producción de iones H+, producto tanto de la reacción de hidrólisis de los iones férricos disueltos, como de la biooxidación de la pirita (Daoud & Karamanev, 2006). Así pues, podría sugerirse que la acidez del medio lixiviante, favorece la solubilidad y disminuye la precipitación de los iones Fe3+, aumentando las tasas de oxidación de pirita y la eficiencia de proceso, debido probablemente a que si el agente químico oxidante se mantiene disponible en la solución por más tiempo, podría darse un mayor ataque oxidativo, permitiendo un incremento en las tasas de extracción del sulfuro metálico.
Vale la pena mencionar, que los microorganismos nativos evidenciaron una mayor disminución en el pH en comparación con el cultivo axénico, independientemente de la concentración de hierro ferroso adicionada a la solución. Esto se debió probablemente a que el aislado nativo logró una mejor adaptación previo al proceso de biodesulfurización, obteniendo una mayor población celular y por ende una mayor actividad oxidativa que le permite generar mayores cantidades de ácido para compensar el consumo de iones H+ por parte de las bacterias durante el proceso. Esta ventaja podría estar relacionada con una capacidad inherente de los microorganismos asociada con al medio natural (drenaje ácido de una mina de carbón) del cual fueron aisladas.
Por otra parte, los resultados de potencial de óxido-reducción obtenidos por los experimentos bióticos realizados (figura 2), mostraron valores alrededor de los 600 mV, similar a otros procesos de biodesulfurización en suspensión (Caicedo & Marquéz, 2011). Podría decirse que los altos valores de Eh fueron debidos al incremento de la concentración inicial de sulfato ferroso, la cual contribuyó con el hecho de que los microorganismos incrementaran su metabolismo, aumentando la relación Fe3+/Fe2+ en el medio lixiviante y generando un ambiente propicio para que se lleve a cabo la oxidación de la pirita (Meruane, 2002).
Adicionalmente, los mayores valores de Eh alcanzados por los microorganismos compatibles con Acidithiobacillus ferrooxidans (NC1 y NC2) indicaron que estos tuvieron una mayor actividad oxidativa en comparación con el cultivo axénico. Esto se debe probablemente a que mantuvieron una constante transferencia de electrones en el sistema, fundamentalmente en lo que se refiere a la regeneración de los iones férricos (Menadier, 2009). Una hipótesis que surge a partir de estos resultados es que el cultivo compatible con Acidithiobacillus ferrooxidans puede contener además de esta bacteria otro tipo de microorganismo causante de este efecto (Boon, 2001).
Vale la pena mencionar que los valores de Eh iniciales están relacionados estrechamente con la concentración inicial de hierro total adicionada al sistema. De esta manera, los ensayos a los cuales se empleó una mayor concentración de hierro (NC2 y PC2) mostraron valores de Eh iniciales bajos en comparación con su contraparte de menor concentración, esto es debido posiblemente a que al iniciar el proceso, la mayor parte del hierro en solución está presente como Fe2+ lo cual generó una disminución en la relación Fe3+/Fe2+ del sistema. En este sentido, al comparar los resultados obtenidos de pH y Eh durante el periodo inicial (día cero a dos), es posible notar que el pH de los ensayos a los cuales se utilizó una mayor concentración de hierro inicial es mayor por lo que se podría inferir que los microorganismos se encuentran activos o preparando la maquinaria metabólica disponible para llevar a cabo el proceso, esta afirmación toma sentido si se tiene en cuenta que los experimentos en los cuales se empleó una menor concentración de hierro inicial mostraron un comportamiento semejante al obtenido en la última etapa de adaptación (datos no mostrados).
Respecto al crecimiento celular, la fase lag evidenciada los primeros días del proceso (figura 3), podría estar relacionada con un atrapamiento de las bacterias en el mineral o con la adición de los iones Fe2+ al sistema. El rápido incremento que exhibió la concentración celular en días posteriores, sugiere que las bacterias prosperaron fácilmente en las condiciones impuestas e indica una respuesta favorable al proceso.
Cabe resaltar, que los resultados obtenidos no indicaron la concentración real de células, debido a que se cuantificó solamente la población de células planctónicas y no se tuvo en cuenta la influencia de las bacterias sésiles. Sin embargo, estudios previos indican que al adicionar iones Fe2+ al medio se disminuye considerablemente la hidrofobicidad, relacionada con la adhesión de las células (mecanismo indirecto de contacto) a los sustratos insolubles tales como pirita (Deveci, 2004).
Las tasas de crecimiento celular alcanzadas por los bioensayos podrían estar asociadas con la disponibilidad de los iones Fe2+ en la solución, de esta manera, al existir una mayor cantidad de sustrato soluble, las bacterias obtienen la energía necesaria para aumentar la población celular. Este comportamiento es coherente con el comportamiento de los demás parámetros evaluados (pH, Eh). Adicionalmente, se observó que adicionar concentraciones iniciales de hierro ferroso propicia la división celular y evita la limitación del crecimiento por falta de sustrato (Bosecker, 1997; Kargi, 1986).
Vale la pena mencionar, que los ensayos NC1 y NC2 lograron las más altas poblaciones celulares, en comparación con sus análogos realizados con la cepa ATCC 23270. Este comportamiento puede estar relacionado con el hecho de que el aislado bacteriano compatible con Acidithiobacillus ferrooxidans se adaptó y próspero fácilmente en las condiciones impuestas, y/o estar asociado con la coexistencia de otro tipo de microorganismos de morfología similar, las cuales se suman al conteo total de células.
El comportamiento estacionario que se evidencia a partir del día cuatro pudo ser consecuencia del agotamiento del sustrato soluble adicionado al sistema, así pues, la pirita es el único sustrato disponible, pero obtener la energía a partir de este es un proceso de cinética lenta, que frena el aumento exponencial del crecimiento celular (Bozdemir et al., 1996). Vale la pena resaltar que a pesar de que la población celular se encontró en fase estacionaria, la transformación de los iones ferrosos en el sistema se mantiene en todos los bioensayos de acuerdo con las curvas de Eh, pH y las medidas de hierro en solución.
En relación con el porcentaje de hierro lixiviado y pirita oxidada (figuras 4 y 5), los ensayos en donde se utilizaron microorganismos compatibles con Acidithiobacillus ferrooxidans presentaron una solubilización significativa de hierro. De esta manera es posible inferir que el proceso de desulfurización depende principalmente de la actividad oxidativa de las bacterias empleadas, esta afirmación adquiere validez si se tiene en cuenta que los experimentos realizados a las mismas condiciones con la cepa axénica (ATCC 23270) mostraron una menor eficiencia. Este hecho podría ser explicado probablemente porque las células compatibles con Acidithiobacillus ferrooxidans transformaron rápidamente los iones ferrosos y mantuvieron las condiciones favorables durante el proceso (bajo valores de pH, altos valores de Eh), lo cual pudo ser la causa del aumento en las tasas de remoción de pirita e impidió la precipitación de los productos de la oxidación respectivamente.
Como ya se ha mencionado anteriormente, los microorganismos compatibles con Acidithiobacillus ferrooxidans mostraron una mayor eficiencia del proceso de biodesulfurización. Una primera hipótesis que surge a partir de estos resultados es que esta ventaja está asociada posiblemente con una capacidad intrínseca -probablemente funcional- referida a un metabolismo hierro oxidante más eficiente, sugiriendo una mayor expresión de proteínas y genes (ej. Iro y rusA), que jugaron un papel central en la cadena de trasferencia electrónica de Fe2+ a O2. El mecanismo biológico que subyace a esta diferencia no es objetivo de este estudio, pero probablemente este asociado a un polimorfismo genotípico de estas células, dependiente del ambiente de evolución que les confiere una ventaja sobre el cultivo axénico (Bird, 2011). Adicionalmente, una segunda hipótesis indica que este comportamiento (mayor actividad oxidativa) podría estar relacionado con un trabajo sinérgico asociado a otro tipo de bacterias quimiolitotróficas presentes en el cultivo o a bacterias mutantes de la misma especie, las cuales contribuyen no solo con la disolución del sulfuro metálico sino que permiten una mayor disminución de los valores del pH y un incremento significativo de los valores de potencial redox (Kargi, 1986). Esta hipótesis resulta probable ya que se debe tener en cuenta que aunque se llevaron a cabo pruebas bioquímicas para corroborar la compatibilidad de la cepa escogida (Acidithiobacillus ferrooxidans), cualquier otra colonia de bacterias de metabolismo similar (oxidantes de hierro y azufre) podría estar presente. Finalmente, los resultados obtenidos en este estudio sugieren que el mecanismo indirecto de no contacto es el más probable para la disolución del sulfuro de hierro, debido posiblemente a que la biolixiviación de metales sulfurados insolubles en ácido esta exclusivamente basado en el ataque oxidativo de los iones Fe3+ (Sand et al., 2001).
Conclusiones
Los ensayos en los cuales se empleó microorganismos compatibles con Acidithiobacillus ferrooxidans presentaron mayor eficiencia en comparación con los experimentos en los cuales se utilizó la cepa axénica (ATCC 23270), ya que obtuvieron el mayor porcentaje de biolixiviación de hierro en solución en las dos concentraciones de hierro inicial evaluadas (200 mg/L y 1200mg/L), alcanzando 68% y 57% de oxidación de pirita respectivamente, después de 12 días de proceso. Esto indica que la adición de los iones ferrosos al medio aumenta la actividad oxidativa de las bacterias y mejora significativamente la oxidación de la pirita. Esto constituye la base principal de este estudio y se considera una buena alternativa a la hora de optimizar el proceso biológico de extracción de azufre inorgánico del carbón.
Agradecimientos
A la empresa cementera ARGOS S.A, a COLCIENCIAS y al grupo de mineralogía aplicada y bioprocesos (GMAB) de la Universidad Nacional de Colombia - sede Medellín, donde se llevó a cabo la investigación.
Referencias bibliográficas
Aller, A., Martinez, O., de Linaje, J. A., Méndez, R., & Morán, A. (2001). Biodesulphurization of coal by microorganisms isolated from the coal itself. Fuel Processing Technology, 69(1), 45-57. [ Links ]
Bird, L. J., Bonnefoy, V., & Newman, D. K. (2011). Bioenergetic challenges of microbial iron metabolisms. Trends in Microbiology, 19(7), 330-340. [ Links ]
Boon, M. (2001). The mechanism of "direct" and "indirect" bacterial oxidation of sulphide minerals. Hydrometallurgy, 62(1), 67-70. [ Links ]
Bosecker, K. (1997). Bioleaching: Metal solubilization by microorganisms. FEMS Microbiology Reviews, 20(3-4), 591-604. [ Links ]
Caicedo, G. (2008). Biodesulfurización de carbón de la zona Río Guachinté - Río Aznazú Valle Del Cauca y Cauca en biorreactores de columna, utilizando microorganismos nativos (tesis de maestría). Universidad Nacional de Colombia, Medellín. [ Links ]
Caicedo, G., & Marquéz, M. (2011). Influencia de la concentración de hierro y pH iniciales en un proceso de biodesulfurización de carbón - ensayos a nivel de laboratorio. Revista Colombiana de Biotecnología, 13(2), 199- 209. [ Links ]
Cara, J., Carballo, M., Morán, A., Bonilla, D., Escolano, O., & García Frutos, F. (2005). Biodesulphurisation of high sulphur coal by heap leaching. Fuel, 84, 1905-1910. [ Links ]
Cardona, C. (2008). Mineralogía del proceso de biodesulfurización de carbones provenientes de la zona del río Guachinté - río Aznazú (Valle del cauca y cauca), (tesis de maestría). Universidad Nacional de Colombia, Medellín. Colombia. [ Links ]
Chou, C. (2012). Sulfur in coals: A review of geochemistry and origins. International Journal of Coal Geology, 100, 1-13. [ Links ]
Daoud, J., & Karamanev, D. (2006). Formation of jarosite during Fe2+ oxidation by Acidithiobacillus ferrooxidans. Minerals Engineering, 19, 960-967. [ Links ]
Harneit, K., Göksel, A., Kock, D., Klock, J. H., Gehrke, T., & Sand, W. (2006). Adhesion to metal sulfide surfaces by cells of Acidithiobacillus ferrooxidans, Acidithiobacillus thiooxidans and Leptospirillum ferrooxidans. Hydrometallurgy, 83(1-4), 245-254. [ Links ]
He, Z., Yin, Z., Wang, X., Zhong, H., & Sun, W. (2012). Microbial community changes during the process of pyrite bioleaching. Hydrometallurgy, 125-126(3), 81-89. [ Links ]
Johnson, D., & Hallberg, K. (2003). The microbiology of acidic mine waters. Research in Microbiology, 154(7), 466-473. [ Links ]
Juszczak, A., Domka, F., Kozowski, M., & Wachowska, H. (1995). Microbial desulfurization of coal with Thiobacillus ferrooxidans bacteria. Fuel, 74(5), 725-728. [ Links ]
León, E. (2006). La importancia del carbón mineral en el desarrollo. Revista Del Instituto de Investigaciones FIGMMG, 9(18), 91-97. [ Links ]
May, N., Ralph, D. E., & Hansford, G. S. (1997). Dynamic redox potential measurement for determining the ferric leach kinetics of pyrite. Minerals Engineering, 10(11), 1279-1290. [ Links ]
Menadier, A. (2009). Biolixiviación de piritas por Acidithiobacillus ferrooxidans y cepas nativas (tesis pregrado).Universidad de Chile, Santiago de Chile. Chile. [ Links ]
Meruane, G. (2002). Oxidación bacteriana de sulfato ferroso con Acidithiobacillus ferrooxidans (tesis doctoral). Universidad de Chile, Santiago de chile. Chile. [ Links ]
Peláez, H., Prada, M., Caicedo, G., Moreno, C., & Marquéz, M. (2013). Influencia de la relación inicial de Fe2+/Fe3+, en un proceso de biodesulfurización de carbones en suspensión. Revista Internacional de Contaminación Ambiental, 29(2), 211-217. [ Links ]
Prayuenyong, P. (2002). Coal biodesulfurization processes. Songklanakarin Journal of Science and Technology, 24, 493-507. [ Links ]
Rawlings, D. (2001). Molecular genetics of Thiobacillus ferrooxidans and other mesophilic, acidophilic, chemolithotrophic, iron- or sulfur-oxidizing bacteria. Hydrometallurgy, 59(2-3), 187-201. [ Links ]
Sand, W., Gehrke, T., Jozsa, P., & Schippers, A. (2001). (Bio)chemistry of bacterial leaching - direct vs. indirect bioleaching. Hydrometallurgy, 59(2-3), 159-175. [ Links ]
Zhang, L., Qiu, G., Hu, Y., Sun, X., Li, J., & Hu, G. (2008). Bioleaching of pyrite by A. ferrooxidans and L. ferriphilum. Transactions of Nonferrous Metals Society of China, 18(6), 1415-1420. [ Links ]

