











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Muchos suelos presentan problemas de disponibilidad de fósforo, especialmente los Andisoles derivados de cenizas volcánicas (Sánchez & Rubiano, 2015). Antes de ser absorbido por las raíces, una cantidad del elemento aplicado se transforma rápidamente en formas menos disponibles, es así que para satisfacer los requerimientos de las plantas para su normal desarrollo y crecimiento, se acude a la fertilización química (Khan et al., 2010). La constante aplicación de fósforo puede alteras algunas propiedades químicas y biológicas del suelo, causando reducción en el rendimiento de los cultivos (Khan et al., 2009), lo anterior genera altos costos (Kaur & Reddy, 2014), obligando a reemplazar el uso de fertilizantes químicos por biofertilizantes (Malusá, 2012).
La comprensión del papel que juegan los microorganismos en el ciclo del fósforo según Mahdi et al. (2011), permite plantear como alternativa viable la introducción como biofertilizantes de microorganismos solubilizadores de P (MSF); estos se concentran en alta proporción en la rizósfera de las plantas, y son metabólicamente más activos que los que provienen de otras fuentes (Arun, 2007). En los últimos años el conocimiento sobre los MSF se incrementó significativamente (Sharma et al. 2013), entre estos se encuentran las bacterias solubilizadoras de fosfato (BSF) capaces de solubilizar varios fosfatos inorgánicos a través de diferentes mecanismos como la producción de ácidos orgánicos (Mishra et al., 2013).
De diferentes suelos se han aislado BSF pertenecientes a los géneros Pseudomonas, Bacillus, Rhizobium, Agrobacterium, Burkholderia, Achromobacter, Acinetobacter, Azospirillum, Azotobacter, Microccocus, Aerobacter, Flavobacterium, Mesorhizobium, Enterobacter y Erwinia (Zhao et al., 2014). Por lo anterior, la presente investigación tuvo como objetivo aislar e identificar rizobacterias con potencial solubilizador de P, en un suelo Andisol localizado en Cajibío (Cauca, Colombia) bajo diferentes sistemas de manejo, posteriormente evaluar la eficiencia solubilizadora de estos microorganismos cuando crecen sobre fosfatos de calcio, aluminio y hierro, utilizando condiciones in vitro.
Materiales y métodos
Aislamiento y evaluación in vitro de la eficiencia solubilizadora de fosfatos. A partir de un Typic Melanudand del municipio de Cajibío, Departamento del Cauca, Colombia, se tomaron muestras de suelo rizosférico en tres agroecosistemas: relicto de bosque secundario (RBS), café (Coffea arabica variedad Caturra bajo sombra de guamo, Inga spp. (CCS) y café (Coffea arabica variedad Caturra) a libre exposición solar o sin sombra (CSS). Una parte de estas muestras fueron analizadas en el Centro Internacional de Agricultura Tropical, CIAT, Palmira, Colombia, otra se trasladó al laboratorio de Microbiología de la Universidad Nacional de Colombia (sede Palmira), donde se procedió al aislamiento de bacterias solubilizadoras de fosfatos (BSF). Para ello, se prepararon diluciones seriales hasta 10-6 con agua destilada estéril. Por cada dilución se sembraron tres repeticiones en agar Pikovskaya - PVK - con una dosis menor de P (Cisneros y Sánchez de P., 2015). Las siembras se incubaron a 27 oC por una semana, luego se seleccionaron aquellos cultivos que presentaron halo claro alrededor de la colonia, el cual resulto de la solubilización de fosfato tricálcico.
Después de obtener los aislados bacterianos, estos se purificaron en el mismo medio por resiembras. Luego, cada aislamiento se sembró en PVK, utilizando por separado tres distintas fuentes de fósforo: fosfatos tricálcico (Ca-P), de aluminio (Al-P) y de hierro (Fe-P). En cada una de las placas de Petri con las diferentes fuentes de P se midió el diámetro de la colonia y del halo que formaron por espacio de 7 días. La evaluación de la Eficiencia Solubilizadora de Fosfato (ESF), en medio sólido, se realizó de acuerdo con la expresión:
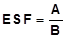
Dónde: A = Diámetro (mm) del halo de solubilización (diámetro colonia + halozona) y B = Diámetro (mm) de crecimiento de la colonia (Cisneros & Sánchez de P., 2015).
Luego se seleccionaron cinco aislados de BSF con mayor ESF promedio en las tres fuentes de P a evaluar en PVK líquido. Para ello, los cultivos bacterianos se incubaron a 28 oC durante siete días, en un agitador orbital a 150 rpm. Al cabo de ese tiempo de incubación, se midió el pH por Potenciometría y el P soluble empleando el Método de Molibdovanadato (Cisneros & Sánchez de P., 2015). Después, se seleccionaron los dos aislamientos bacterianos con mayor capacidad solubilizadora de P promedio en los medios probados. Estos aislados se identificaron por técnicas de biología molecular, se evaluaron para producción de ácidos orgánicos y posteriormente su efecto en plántulas de café (Cisneros et al., 2017).
Identificación de las BSF. A los dos aislamientos bacterianos seleccionados en medio líquido, se les extrajo el ADN utilizando el kit comercial: QIAamp DNA Mini Kit De Qiagen(. La identificación de los aislamientos bacterianos se llevó a cabo mediante el gen que codifica la región 16S rRNA, se amplificó a partir de las BSF seleccionadas, por la reacción en cadena de la polimerasa (PCR) utilizando los cebadores fD1 (5'-AGAGTTTGATCCTGGCTCAG-3') y rD1 (5'-AAGGAGGTGATCCAGCC-3') (Pérez et al., 2007; Matías et al., 2009). Se empleó unas reacciones de PCR en un volumen total de 50 µl que contenía 0.1 mM desoxinucleótidos (DNTPs), 0.2 mM de cada cebador, 2.5 mM de MgCl2, 20 mM de Tris HCl, 50 mM KCl, 1U de Taq polimerasa (Invitrogen) y 50 ng de ADN. Se completó con agua libre de RNasas. La amplificación se realizó en un termociclador PT100 (MJ-Research, Watertown, MA). Luego de la amplificación, se tomaron 5 µl de los productos amplificados, los cuales se visualizaron en gel de poliacrilamida al 7% (37:1 acrilamida-bisacrilamida) corridos a 160 voltios por una hora y teñidos en bromuro de etidio (Sambrook et al., 1989).
El tamaño aproximado de las bandas del producto PCR se comparó con un marcador de peso molecular GeneRulerTM de 100bp Thermo Scientific®. Los amplificados observados en los pesos registrados se enviaron a purificar y a secuenciar a Macrogen inc. Korea. Las secuencias sentido y antisentido ("forward" y "reverse") fueron editadas utilizando el software de bioinformática BioEdit v7.2.5 (Hall, 1999), una vez limpias las secuencias, se realizó el alineamiento con la opción online de BLAST (bl2seq) y se construyó la secuencia consenso, finalmente se comprobó su identidad en la base de datos de secuencias de DNA (http://www.ncbi.nlm.nih.gov/blast/Blast.cgi), utilizando el algoritmo BLAST.
Producción de ácidos orgánicos como posible mecanismo microbiano de solubilización de fosfatos. Pasados 7 días de incubación en medio líquido, se tomó una alícuota de 20 µl de los filtrados de las dos bacterias más eficientes en solubilización de fosfatos e identificadas molecularmente, según metodología descrita arriba (medio líquido), con el fin de evaluar la producción de los siguientes ácidos orgánicos (acético, cítrico, glucónico, málico y láctico) por Cromatografía Líquida de Alta Eficiencia (HPLC) con una columna de exclusión por tamaño de ion. Para la detección de cada ácido orgánico se utilizó un equipo marca Waters serie con software Breeze, usando las recomendaciones de Mardad et al., 2013. Dichas condiciones fueron: Columna Aminex HPX-87H (300 mm x 7.8 mm), temperatura de 35oC, fase móvil H2SO4 0.0025 M, flujo de 0.5 ml/min, detector: ultravioleta (UV Waters 2487), longitud de onda: 210 nm. La identificación de los ácidos orgánicos se llevó a cabo por comparación de los tiempos de retención de sus estándares patrón con los de los picos del cromatograma de cada muestra obtenida (Mardad et al., 2013).
Análisis estadístico. Para evaluar la eficiencia solubilizadora de las BSF en medios sólido y líquido, por cada aislado bacteriano, se utilizó un diseño completamente al azar, con tres fuentes de fósforo (Ca-P, Al-P y Fe-P) y tres repeticiones. Las diferencias en las actividades de los microorganismos se analizaron a través de una estadística descriptiva, de un análisis de varianza, de prueba de promedios (Tukey) y correlaciones de Pearson. En todos los casos con un nivel de significancia del 95%. Todos los análisis se realizaron utilizando el software SAS versión 9.3 (2010).
Resultados y discusión
Los análisis químicos del suelo son consistentes con los descritas para Andisoles, ya que presentan algunas de sus características como pH ácido (<5.5); alto contenido de materia orgánica (>10%), alta capacidad de intercambio de cationes (CIC) y muy baja disponibilidad de P (Cisneros y Sánchez de P., 2015).
Aislamientos de bacterias solubilizadoras de fosfatos (BSF)
Los resultados indican que la población bacteriana con capacidad solubilizadora de fosfatos está presente en los tres agroecosistemas, de hecho se lograron aislar 26 bacterias con capacidad solubilizadora en fosfato tricálcico.
Los aislados bacterianos que formaron halo, correspondieron al 63.6% de los obtenidos en RBS, 60.0% de los procedentes de CCS y 52.6% de CSS. Lo anterior muestra que aunque un número mayor de poblaciones bacterianas se adquirieron de los agroecosistemas CSS y CCS, las BSF tuvieron mayor frecuencia relativa en RBS, cuyos contenidos de fósforo (P-Bray II) son los más bajos, lo cual sugiere la escasa disponibilidad del fósforo como una causa de adaptación de estos microorganismos al suelo (Cisneros R. et al., 2014). También estos resultados reflejan la baja intervención antrópica en RBS comparada con CSS y CCS, es decir, son consecuencia de prácticas agrícolas basadas en productos de síntesis química industrial aplicados a los dos últimos agroecosistemas.
Evaluación in vitro de la eficiencia solubilizadora de las BSF en medio sólido
El análisis de varianza muestra diferencias significativas en la actividad solubilizadora de las bacterias; la prueba de Tukey revela que el mayor valor de Eficiencia Solubilizadora de Fosfato (ESF) se presenta en el medio Ca-P, cuyo valor difiere significativamente de lo observado en Al-P y, en Fe-P cuyos valores fueron menores. Lo anterior es registrado por investigadores como Acevedo et al. (2014), y Asuming & Aferi (2014), quienes reportaron que algunos microorganismos presentan alta capacidad de solubilización de Ca-P comparado con otras fuentes como Al-P y Fe-P. Dicha solubilización es explicada a través de los valores de Kps (constante del producto de solubilidad o constante de equilibrio de iones en solución) para los diferentes fosfatos (Khan et al., 2007).
Con lo anterior, se procedió a seleccionar los cinco aislamientos bacterianos más eficientes en las tres fuentes de P, los cuales fueron identificados como UNB1, UNB2, UNB3, UNB4 y UNB5. Posteriormente, estos fueron evaluados en medio de cultivo líquido.
Estimación de P soluble por parte de los microorganismos seleccionados
Se presentan diferencias altamente significativas en la actividad solubilizadora de estas bacterias de acuerdo con la fuente de fosfato empleada. Para todos los microorganismos, los mayores valores de solubilización de P insoluble se obtuvieron en Ca-P, seguido de Al-P y de Fe-P (tabla 1).
Tabla 1 Concentración de P soluble (en %) en los tres medios de cultivo inoculados con los aislamientos bacterianos. Agrupamiento según la prueba de Tukey. Nota: Dentro de la misma columna, promedios con la misma letra no difieren estadísticamente.
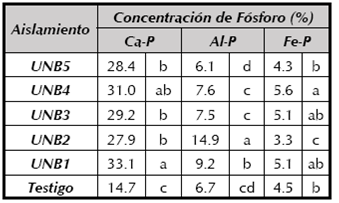
De forma similar varios autores han reportado esta secuencia de solubilización (Acevedo et al., 2014; Asuming & Aferi, 2014). En Ca-P, los aislamientos UNB1 y UNB4 presentan mayor porcentaje de P soluble, sin embargo, éstos no muestran diferencias significativas entre sí, evidenciándose un efecto contrario con el testigo (tabla 1). Con respecto a Al-P, las bacterias UNB2 y UNB1, obtienen el porcentaje de P soluble más alto, los cuales son estadísticamente diferentes, y el aislado UNB5 el menor valor, superado en términos estadísticos por el testigo. Finalmente en Fe-P, no existen diferencias estadísticas entre los porcentajes de P soluble obtenidos de los aislados UNB4, UNB3 y UNB1, presentando los valores más altos, mientras que UNB2 presenta el menor valor, el cual, a su vez, se vio superado por el testigo y el aislamiento UNB5.
La solubilidad de P procedente de cada fosfato representa las concentraciones de sus iones constituyentes que pueden mantenerse en solución bajo condiciones de equilibrio. Cuando la concentración del P soluble es mayor que la soportada por la solubilidad del fosfato, el P soluble se precipita con los cationes Ca2+, Al3+ y Fe3+ y forma los fosfatos mencionados. Cuando el P soluble es menor, los fosfatos se disuelven en la solución. Lo anterior es explicado a través del principio de Le Châtelier que está relacionado con los sistemas en equilibrio.
Con respecto al pH (datos no mostrados), después de 7 días de cultivo, este se reduce entre 1 y 2 unidades aproximadamente en relación con el pH que presenta el testigo (aproximadamente neutro). La reducción es en general más marcada cuando se utiliza Al-P y Fe-P, siendo esta última fuente de P la que muestra mayor acidificación con respecto al testigo. Los valores de pH bajo obtenidos (alta acidez) están ligados a la liberación de iones H+ que balancean la toma de cationes amonio, NH4 + procedente del medio de cultivo y a la producción de ácidos orgánicos de bajo peso molecular por las bacterias solubilizadoras de fosfato, BSF, cuyas propiedades quelantes ayudan a la formación de complejos insolubles con iones metálicos como Ca2+, Al3+ y Fe3+, con la consecuente liberación del P en forma de fosfato (Beltrán, 2014).
Finalmente, se encontró relación inversa entre pH y P soluble en los medios Ca-P y Fe-P, con coeficientes de correlación de Pearson de -0.895 y -0.893 respectivamente, resultados que coinciden con registros de Prada (2013).
Identificación molecular de las bacterias seleccionadas
Con los resultados obtenidos de P soluble en las tres fuentes de fosfatos, se seleccionaron los dos aislamientos bacterianos en promedio con mayor capacidad solubilizadora de fosfatos (tabla 1), los cuales fueron identificados como UNB1 y UNB2.
La amplificación por PCR de la región 16S del ADNr de las bacterias, generó un amplicón aproximadamente de 1500 pb con los cebadores fD1 y rD1, además de un peso molecular igual al obtenido por Patiño & Sánchez de P. (2012).
Análisis de secuencias en el GenBank
Las secuencias obtenidas fueron comparadas con la base de datos del GenBank mediante el programa BLAST, revelando que el organismo UNB1 está relacionado con el género Kocuria sp., identidad de 99% (www.ncbi.nlm.nih.gov/genbank). Las bacterias del género Kocuria (familia Micrococcacea, orden Actinomycetales, clase Actinobacteria) son cocos grampositivos (Dastager et al., 2014). Han sido aisladas de variedad de fuentes naturales incluyendo el suelo, la rizósfera, ambientes acuáticos (dulces y marinos), entre otros (Chandna et al., 2013).
Estas bacterias en forma general, han sido consideradas como organismos promotores de crecimiento vegetal por su capacidad de producir sustancias benéficas que también ayudan a controlar la población de hongos fitopatógenos (Goswami et al., 2014); también cumplen un papel importante en la descomposición de materia orgánica, pudiendo ser empleadas como agentes de carga en la producción de compost de alta calidad (Chandna et al., 2013), además presentan un amplio uso en la bioremediación de suelos (Dastager et al., 2014).
Prada (2013) señaló que de 57 aislados de actinobacterias sólo diez mostraron mediante evaluaciones cualitativas y cuantitativas tener una actividad solubilizadora de fósforo significativamente alta y la presencia dominante de los ácidos oxálico, cítrico y glucónico. De igual manera, Goswami et al. (2014), encontraron que Kocuria turfanensis es un eficiente microorganismo solubilizador de fosfato, productor de sideróforos y amoniaco. Por otra parte, el análisis de las secuencias del aislamiento UNB2 mostró un nivel de identidad 98% con Bacillus subtilis (www.ncbi.nlm.nih.gov/genbank).
Las bacterias grampositivas, aerobias facultativas, que pertenecen al género Bacillus (familia Bacilliaceae), se encuentran distribuidas en diferentes tipos de ambientes como el suelo, plantas y ecosistemas de aguas dulce y marina (Badía et al., 2011). Son microorganismos capaces de resistir condiciones físicas extremas por formar endosporas lo cual influye en su amplia distribución y adaptabilidad a diferentes ecosistemas (Kumar et al., 2012; Chen et al., 2015). Entre las especies de Bacillus se registran B. subtilis, entre otras (Kumar et al., 2012). Este género ha demostrado capacidad para incrementar el crecimiento de las plantas (Angulo et al., 2014). Varias investigaciones han demostrado la capacidad del género Bacillus como solubilizador de fosfato (Maheswar & Sathiyavani, 2012; Tejera et al., 2013). A su vez, tiene capacidad para fijar nitrógeno (Beneduzi et al., 2008) y producir reguladores del crecimiento vegetal como auxinas, citoquininas y/o giberelinas (Mutua et al., 2011).
Estimación de la producción de ácidos orgánicos
Los aislamientos acidificaron el medio de cultivo, posiblemente producto de la biosíntesis y liberación de una amplia variedad de ácidos orgánicos, considerado uno de los mecanismos más importantes utilizado por los microorganismos para solubilizar fuentes de fosfatos. Las dos bacterias aisladas e identificadas (Kocuria sp, y Bacillus subtilis) produjeron diferentes ácidos orgánicos en las tres fuentes de fosfato, excepto ácido oxálico (utilizado como uno de los estándares que confirmaron la presencia de los diferentes ácidos orgánicos), en menos de 20 minutos (figura 1): 1. ácido oxálico: 8.1 minutos, 2. ácido cítrico: 10.1 minutos, 3. ácido glucónico: 10.7 minutos, 4. ácido D-málico: 12.2 minutos, 5. ácido D-láctico: 13.9 minutos, 6. ácido L-láctico: 15.3 minutos, 7. ácido L-málico: 17.8 minutos y 8. ácido acético: 19.1 minutos. Sus concentraciones variaron con la fuente de P a la que se expuso el microorganismo (tabla 2). Estos resultados concuerdan con los obtenidos por varios autores que también encontraron una concentración y perfil de ácidos diferentes según la fuente de P usada (Vyas & Gulati, 2009; Prada, 2013).
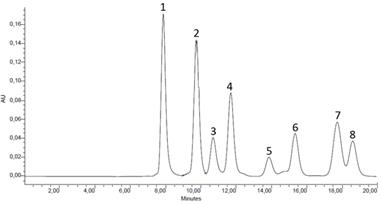
Figura 1 Cromatograma de HPLC de los estándares de ácidos orgánicos, separados en menos de 20 minutos. Nota: longitud de onda utilizada 210 nm (Mardad et al., 2013)
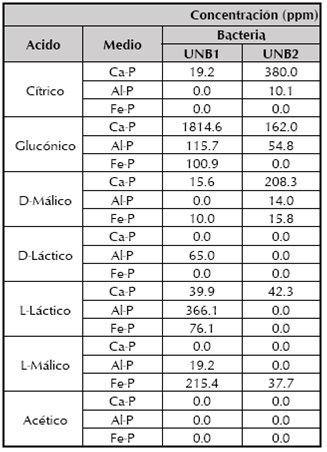
Tabla 2 Concentración de ácidos orgánicos producidos por las bacterias en las tres fuentes de P empleadas.
Los diferentes ácidos orgánicos presentes en una de las dos muestras de cultivos bacterianos evaluados, se ilustran en los cromatogramas de la figura 2: UNB1, Kocuria sp (a, b, c). En los cromatogramas se observan compuestos desconocidos (*), adicionalmente se visualizan picos con intensidades diferentes para cada fuente de fosfato, por lo cual se puede deducir que las concentraciones de ácidos orgánicos producidas por las bacterias también varían con respecto a la fuente de P empleada. Cromatogramas similares se observan en UNB2 (Bacillus subtilis).
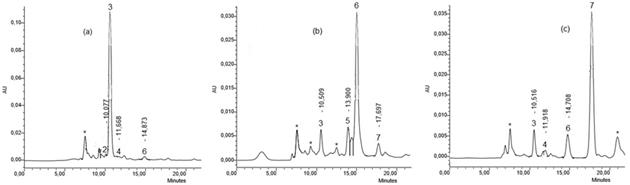
Figura 2 Cromatogramas de HPLC de los ácidos orgánicos producidos por UNB1 (a) con Ca3(PO4)2, (b) con AlPO4 y (c) con FePO4 como fuente de fósforo
De acuerdo con la tabla 2, los ácidos orgánicos producidos en mayor proporción en las tres fuentes de P, por las bacterias, en su orden son: ácidos glucónico > L-láctico > cítrico > L-málico.
En general, hay mayor diversidad de ácidos orgánicos liberados en Ca-P que en Al-P y en Fe-P. Este hecho puede estar relacionado con la facilidad de liberar mayor cantidad de ortofosfato a partir de Ca-P que de Al-P y de Fe-P; adicionalmente se destaca que Kocuria sp con los ácidos glucónico y L-láctico, fue el microorganismo que formó ácidos orgánicos en las tres fuentes de P evaluadas (tabla 2). Con respecto a la concentración de cada ácido orgánico, en Ca-P y en Fe-P, Kocuria sp fue el microorganismo que más altas concentraciones generó (ácido glucónico: 1814.6 ppm y ácido L-málico: 215.4 ppm). Existen pocos reportes de la actividad solubilizadora de P y producción de ácidos orgánicos de Kocuria sp, sin embargo Goswami et al. (2014), encontraron que Kocuria turfanensis es un eficiente microorganismo solubilizador de fosfato, productor de sideróforos y amoniaco. En cuanto a Bacillus subtilis, Sharma et al. (2013), y Behera et al. (2014), mencionan la producción de ácidos cítrico, láctico, málico, glucónico y oxálico, responsables de la solubilización de fosfatos y que explican la reducción de pH en los medios de cultivo.
Los ácidos orgánicos producidos por las bacterias pueden disolver los fosfatos empleados como resultado de intercambio aniónico o quelatación con los iones Ca+2, Al+3 o Fe+3 asociados con los fosfatos insolubles Ca-P, Al-P y Fe-P (Sharma et al., 2013), se cree que más que la concentración o cantidad de ácidos orgánicos, es su estructura la que puede determinar la solubilización de fosfatos (Vyas & Gulati, 2009), en ese sentido, los ácidos orgánicos producidos por los microorganismos se desprotonan para quelatar los cationes metálicos Ca+2, Al+3 y Fe+3 involucrados en los fosfatos empleados.
Por otra parte, los coeficientes de correlación de Pearson entre la producción de ácidos orgánicos y el fósforo soluble en los tres fosfatos no arrojaron valores significativos (p> 0.05), por lo anterior se cree que la obtención de ácidos orgánicos no parece ser el único mecanismo de solubilización en este trabajo, ya que en algunos casos pueden intervenir otros factores encargados de reducir el pH de los medios de cultivo, por ejemplo, la fuente de N empleada. Se han registrado altas tasas de solubilización en presencia de sales de amonio como sulfato de amonio (empleado en este trabajo). Lo anterior se atribuye a la liberación de H+ que compensan la toma del ion amonio, bajando el pH (Beltrán, 2014). Adicionalmente, Bolívar et al. (2016), mencionan que en Burkholderia tropica, existe un mecanismo adicional, ya que esta especie solubiliza fosfato incluso en un medio fuertemente amortiguado.
Las bacterias pueden tener capacidad de hacer uso de diferentes mecanismos para solubilizar el P, la estimulación puede depender de diversos factores, entre ellos, la procedencia de los aislamientos, las poblaciones alcanzadas, entre otros. Paralelamente, el mecanismo de solubilización de P, puede estar influenciado por el género, la especie o medio de cultivo en el cual se realice la evaluación (Panhwar et al., 2012). En los medios de cultivo, las fuentes de C, de N y las sales (MgSO4, KCl, NaCl), influyen en gran medida en los potenciales de solubilidad de fósforo de los microorganismos. Sin embargo, el uso de una fuente de N (sulfato de amonio) en la composición estimula de manera diferente el metabolismo del C e induce la vía metabólica, lo que puede generar la producción de compuestos complejos con capacidad quelante pero con diferente estructura u origen (Prada, 2013; Sharma et al., 2013).
Conclusiones
En tres agroecosistemas (café sin sombra, café con sombra y relicto de bosque secundario) de un Typic Melanudand de Cajibío (Cauca), se identificaron bacterias con capacidad solubilizadora de fosfatos de Ca, Al y Fe, encontrándose la máxima actividad solubilizadora en Kocuria sp. y Bacillus subtilis.
En las tres fuentes de P (fosfatos de Ca, Al y Fe), se estimaron los siguientes ácidos orgánicos: cítrico, glucónico, D- y L-málico, D- y L-láctico y acético, por parte de las dos bacterias con mayor capacidad solubilizadora de P, se observó que existe mayor diversidad de ácidos orgánicos en la fuente Ca-P y que sus concentraciones variaron con respecto a la fuente de P para cada microorganismo.

