











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Oxalis tuberosa Molina, conocida como oca en Perú, Ecuador y Bolivia, cuiba o quiba en Venezuela, ibia en Colombia, truffette acide en Francia o papa extranjera en México es después de la papa, el tubérculo más conocido en la región Andina, en donde sus tubérculos amiláceos constituyen un componente básico para la alimentación. Hoy en día es cultivada en países como México y Nueva Zelanda en donde son apreciadas por presentar colores y sabores muy particulares. Es una planta dicotiledónea, herbácea, anual, de sabor dulce y consistencia harinosa; cultivada en altitudes comprendidas entre los 2800 y 4000 msnm, toleran climas fríos, fotoperiodo óptimo es de 9 horas y prosperan en suelos con pH de 5.3 a 7.8, presenta resistencia a plagas y otros problemas fitosanitarios, por lo tanto tiene una mayor adaptación a ambientes en donde otros cultivos no pueden sobrevivir (Rosero, 2010; Clavijo y Pérez, 2014).
Las ibias presentan alta variación en sus niveles nutritivos dependiendo de la variedad, con un contenido de proteína de aproximadamente el 9%, altos porcentajes de almidón (13.2%), minerales y ácidos orgánicos los cuales facilitan su uso en procesos industriales tales como la panificación, deshidratación y extracción de alcohol mediante fermentación (Moscoe et al., 2016). El tubérculo presenta altas concentraciones de ácido oxálico y de una proteína de almacenamiento llamada ocatina, las cuales juegan un papel importante en la resistencia a los patógenos (Bradbury & Emshwiller, 2011). A pesar de su potencial económico promisorio, debido a sus propiedades agroindustriales y medicinales y gran adaptabilidad a condiciones ambientales difíciles, es una especie marginal por factores tales como las dificultades en el mercadeo del producto, bajo prestigio social (constituyen un alimento básico para las comunidades pobres de los Andes), laboriosos procesos de cocción y bajo retorno económico (Clavijo et al., 2011, Malice et al., 2010; Jiménez et al., 2015).
Históricamente los países en donde se han realizado más estudios sobre colecta, conservación y utilización de estos recursos genéticos son Bolivia, Perú y Ecuador, por el hecho de que estos países presentan la mayor diversidad de estas especies (Rosero, 2010, Clavijo et al., 2015). En Colombia estos cultivos se encuentran distribuidos en los pisos térmicos fríos de los departamentos de Nariño, Cauca, Cundinamarca y Boyacá, en donde hacen parte del patrimonio cultural heredado por los Paeces, Muiscas y Laches, entre otros. En la provincia de Sugamuxi, departamento de Boyacá se conservan aún los cubios o nabos, rubas o chuguas e ibias como parte fundamental de su alimentación; sin embargo, se encuentran las mayores producciones en los municipios de Sogamoso, Aquitania y Gámeza (Clavijo et al., 2011; Aguirre et al., 2012).
Las ibias han sido propagadas exclusivamente a través de tubérculos, dando origen a un gran número de variedades clonales que pueden ser distinguidas especialmente por el color, textura y la forma del tubérculo (Bradbury & Emswiller, 2011). Por lo tanto, el uso de tan pocos criterios de selección podría explicar la confusión entre plantas que comparten fenotipos similares pero diferentes genotipos y lo tanto, la existencia de la diversidad genética intravarietal ya observada por Pissard et al. (2007). Oxalis tuberosa, podría considerarse una especie modelo para los estudios de diversidad genética en plantas marginales y propagadas vegetativamente, como se les conoce a los tubérculos andinos. Esta diversidad está dada por el sistema de reproducción, la prevalencia de variedades tradicionales, la falta de mejoramiento, la introducción inconsciente de materiales producto de la reproducción sexual y su adaptación a condiciones extremas, entre otros (Clavijo et al., 2015; Moscoe et al., 2016).
La diversidad genética de Oxalis tuberosa no ha sido estudiada suficientemente en Colombia y no existen bancos de germoplasma que conserven una muestra representativa de su variabilidad (Rosero, 2010). Estudios de caracterización morfológica en diferentes tubérculos andinos muestran que los descriptores cualitativos que permiten una mejor diferenciación de los materiales son el color de los tallos aéreos, pigmentación de las axilas, color del follaje, color de la flor, color del pedúnculo y pedicelo, color predominante en la superficie de los tubérculos, color de la pulpa, color y forma de los tubérculos, lo cual ha permitido la selección y la identificación de las variedades por parte de los agricultores, quienes usan como criterio único de selección características morfológicas asociadas principalmente al tubérculo (Cadima 2006; Malice et al., 2007; Iriondo et al., 2008; Rosero, 2010).
Las ibias son un cultivo que se propaga vegetativamente y que por procesos antrópicos y ambientales se han generado variedades clonales, sin embargo, se hace necesario una caracterización más objetiva del germoplasma de este recurso fitogenético, que nos permita entender mejor la evolución de los cultivos clonales dentro de los sistemas agrícolas tradicionales considerando el papel cultural, económico y ecológico de ambiente, el impacto del intercambio de material de siembra y la influencia de la reproducción sexual en la generación de su diversidad genética (Malice et al., 2010). Hoy en día, existen herramientas de la biología molecular, que nos permiten identificar esa variación a nivel del ADN, como son los marcadores moleculares los cuales no están sujetos a la variación ambiental y nos permiten una estimación más objetiva de la diversidad genética presente en el germoplasma (Kuchma et al., 2011). Los estudios de caracterización molecular que usan marcadores dominantes y codominantes en O. tuberosa muestran que hay una alta diversidad genética y que no hay relación entre los morfotipos y los genotipos, que algunos agrupamientos pueden estar determinados por la mayor o menor cantidad de ácido oxálico presente en los materiales evaluados y que también éstos se corresponden con los niveles de ploidía (Pissard et al., 2006, 2007a, b, 2008; Emshwiller et al., 2009; Malice et al., 2007, 2009, 2010; Morillo et al., 2016).
Los marcadores moleculares conocidos como los RAMs o ISSR (Intesimple Sequence Repeat) son útiles para medir la diversidad genética en plantas y animales, permiten identificar diferencias entre familias, entre especies y al interior de las especies muestran la base misma de la variación de los individuos, permiten analizar la información que se expresa como la que no lo hace (Muñoz et al., 2008). Esta metodología es factible para pequeños laboratorios, no requiere el conocimiento previo de la secuencia y se ha utilizado para la determinación de la diversidad genética en plantas cultivadas como en tubérculos andinos (Mora et al., 2013; Morillo et al., 2015; Morillo et al., 2016). La caracterización molecular usando diferentes tipos de marcadores ha permitido sentar las bases para la investigación y conservación de materiales propagados vegetativamente, el análisis comparativo del cariotipo y la identificación de metabolitos de interés, entre otros (Bonna et al., 2017; Moscoe et al., 2016, Zhang et al., 2017).
Considerando que los tubérculos andinos son cultivos relegados pero que tienen una gran importancia social a nivel de las poblaciones rurales andinas y que hay diversidad en estos materiales, la cual está amenazada por la pérdida de la diversidad genética, el presente trabajo de investigación tuvo como objetivo principal la caracterización morfológica y molecular de materiales de ibias en el departamento de Boyacá, como una primera aproximación hacia el mejoramiento genético de esta especie en esta región.
MATERIALES Y MÉTODOS
Colecta
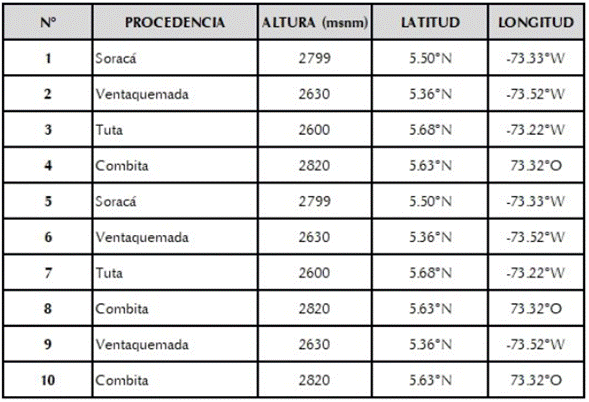
Los materiales utilizados para la caracterización morfológica y molecular fueron colectados en los principales municipios productores del departamento de Boyacá: Soracá, Ventaquemada, Tuta y Combita. Se seleccionaron plantas con buen porte, arquitectura, buen rendimiento, con ausencia de síntomas causados por factores bióticos o abióticos. Los materiales colectados se sembraron en una parcela demostrativa en la finca “Asuazan” ubicada en el municipio de Combita-Boyacá (5° 38´ 30.27” N, 73° 19´34.84” E), la cual presenta una precipitación promedio de 870 mm/año, temperatura promedio anual de 15°C y una humedad relativa entre el 70 y 80%; en donde cada uno de los materiales colectados se sembró en diferentes sitios de la parcela, para lograr así tener cuatro plantas por genotipo para posteriormente hacer las evaluaciones correspondientes (tabla 1) y poder tener así una colección in situ de la variabilidad genética colectada.
Caracterización morfológica
La caracterización morfológica de los materiales colectados se hizo de acuerdo con los descriptores evaluados por Rosero (2010). Para la medición de las variables cualitativas como fue el color, se utilizó un colorímetro digital marca Minolta, donde se determinaron los parámetros del sistema CIELab “L”, “a” y “b”, a cada fruto se le hicieron tres lecturas en el diámetro ecuatorial, donde L indica la luminosidad, tomando valores de 0 que es negro y 100 es blanco; para los valores de “a” donde menor a 0 es verde y mayor a 0 es rojo, para “b” los valores menores de 0 indican azul y mayores de 0 amarillo (tabla 2).
Caracterización molecular
La caracterización molecular se llevó a cabo en los laboratorios de investigación en Biología Molecular de la Universidad Pedagógica y Tecnológica de Colombia, Tunja. La extracción del ADN se hizo utilizando tejido foliar joven siguiendo el protocolo de Dellaporta et al. (1983), y se visualizó en geles de agarosa al 0.8% teñidos con Gel Red en una cámara Maxicell Primo EC-340 Electroforesis Gel System; se cuantificó mediante curva de dilución con el ADN del bacteriófago Lambda (20 ng/µl). El ADN cuantificado se diluyó en agua tipo HPLC a un volumen total de 100 μl a 10 ng/μl y se almacenó a -20 °C. Para la caracterización molecular se utilizaron siete cebadores RAMs sintetizados por Technologies Inc. (tabla 2). Para la reacción de amplificación con RAMs se preparó el cóctel en un tubo estéril de microcentrífuga (1.5 ml) para un volumen final de 25 μl. La mezcla de reacción se preparó con buffer1X, MgCl2 1.5 mM, DNTPs 0.2 mM, Taq Polimerasa 1U, cebador 2 μmM y ADN genómico 10ng.
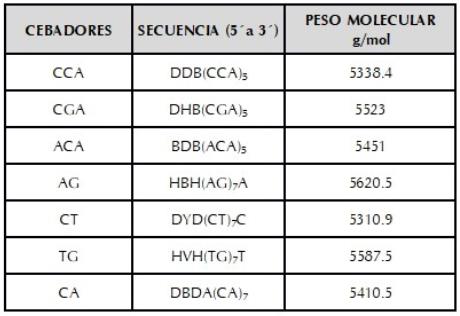
Las siguientes designaciones son usadas para los sitios degenerados: H (A ó T ó C); B (G ó T ó C); V (G ó A ó C) y D (G ó A ó T).
La amplificación se llevó a cabo en un termociclador PTC 100 Programmable Termal Controller (MJ. Research, Inc.). La desnaturalización inicial fue a 95 °C durante 5 minutos; desnaturalización a 95 °C por 30 segundos, hibridación a una temperatura de 50 °C (cebador AG y CA), 55 °C (cebador CCA-TG-CT) y 58 °C (cebador CGA) durante 45 segundos, una extensión de 72 °C por 2 minutos, 37 ciclos desde la desnaturalización a extensión y por último una extensión a 72 °C durante 7 minutos. Los productos de amplificación se separaron por electroforesis en geles de agarosa de alta resolución al 1.5% a 90 voltios durante 3 horas, tenidos con gelred y se visualizó en un transiluminador.
Análisis de la información
Con los datos obtenidos de la caracterización morfológica se realizaron análisis multivariados usando el programa estadístico InfoStat versión 2015; se obtuvo la matriz de correlaciones y se hizo el Análisis de Componentes Principales, para determinar las características más discriminantes dentro del conjunto de variables evaluadas. Con los datos moleculares se generó una matriz binaria de ausencia (cero) y presencia (uno) de alelos, la similitud genética se calculó utilizando el coeficiente de similitud de Nei y Li (1979). El análisis cluster se realizó por el método UPGMA y se generó un dendrograma considerando los datos morfológicos y moleculares, utilizando el paquete estadístico NTSYS (Numerical Taxonomy System for personal Computer, versión 2.02 PC). Para evaluar la variabilidad genética se estimó la heterocigosidad insesgada y el porcentaje de loci polimórficos utilizando el paquete estadístico TFPGA (Tools For Population Genetic Analices, versión 1.3, 1997). Se determinó el f estadístico insesgado con un intervalo de confianza del 95 % en los materiales de ibias colectados.
RESULTADOS Y DISCUSIÓN
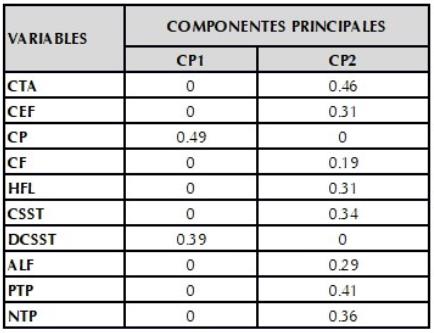
En la colecta realizada en la zona de estudio se encontraron 10 materiales de ibias con características fenotípicas distintas y clasificadas como variedades por parte de los agricultores (tabla 1). En el análisis de componentes principales se encontró que los cuatro primeros componentes explican más del 70% de la variación fenotípica observada en los materiales de ibias evaluados, siendo el Componente principal 1 (CP1) y el dos (CP2) los que tienen una contribución de más del 90% Las variables que aportan de manera significativa en cada uno de estos componentes son para el CP1: Color del peciolo (CP), distribución del color secundario de los tubérculos (DCSST) y CP2: Color de los tallos aéreos (CTA), color del envés de los foliolos (CEF), hábito de floración (HFL), color secundario de la superficie de los tubérculos (CSST), peso de tubérculos por planta (PTP) y número de tubérculos por planta (NTP) (figura 1).
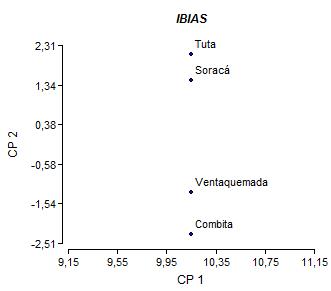
Figura 1 Dispersión de los materiales de ibias evaluados con base en los caracteres para tallos, hojas, flores y tubérculos asociados a CP1 y CP2 por localidad.
En la tabla 4 se muestran la contribución de cada una de las variables a los primeros componentes principales, en el primero se puede observar que las variables distribución del color secundario en la superficie del tubérculo (DCSST, 0.39), color del pecíolo (CP, 0.49) y la forma del tubérculo son las variables que mayor aporte están haciendo a la variabilidad fenotípica observada. En cuanto al componente principal 2, las variables más discriminantes son el color de los tallos aéreos (CTA, 0.46), peso de los tubérculos por planta (PTP, 0.41), número de tubérculos por planta (NTP, 0.36), color secundario de la superficie del tubérculo (CSST, 0.34), color del envés del foliolo (CEF, 0.31) y hábito de floración (HFL, 0.31), características que deben ser tenidas en cuenta en estudios de la identificación de la variabilidad fenotípica en materiales de ibias, especialmente el color característica asociada a la presencia de metabolitos secundarios de interés industrial y/o farmacéutico ( Cruz et al., 2017; Zhang et al., 2017).
Tabla 4 Vectores propios y la contribución a la varianza total explicada por los componentes principales uno y dos.
En Colombia se han venido realizando estudios de caracterización morfológica en papa en donde Moreno y Valbuena (2006) encontraron que cinco caracteres cualitativos a nivel del tubérculo y de la flor discriminaron la colección. Bernal et al. (2007), observaron que las variables que más discriminan la colección Colombiana de papa son el color primario de la piel del tubérculo y su intensidad, color secundario de la flor y su distribución, color secundario de la piel del tubérculo y su distribución, color secundario de la pulpa del tubérculo y su distribución, color primario de la flor y color del pedicelo, características que han mostrado ser discriminantes en diferentes especies tuberosas. Los resultados encontrados en este estudio concuerdan con las observaciones realizadas en otros estudios de caracterización morfológica en materiales de ibias en donde se consideran que las características asociadas al color y forma del tubérculo son determinantes a la hora de evaluar morfológicamente los materiales y es el único criterio de selección usado por los agricultores para la determinación e identificación de variedades (Cadima, 2006; Bernal et al., 2007; Morillo et al., 2016).
Estudios de colecta, caracterización y conservación de la variabilidad de la ibia en agroecosistemas paramunos del departamento de Nariño muestran al igual que en este estudio que las características más discriminantes están relacionadas con el color del tubérculo, que presenta una variación continua desde el blanco hasta el negro, pasando por diferentes tonalidades de amarillo, rosado y rojo. El color de la pulpa parece tener también una pequeña variación de tipo continuo, con rangos que van desde el blanco marfil, amarillo y púrpura-morado Una gran cantidad de formas presenta el anillo vascular del tubérculo pigmentado con la misma coloración de la piel, seguido con la intensidad del color la médula (Rosero, 2010).
Como se había reportado ya en caracterizaciones anteriores las variables cualitativas que están diferenciando fácilmente los materiales de ibias son: color de los tallos aéreos, pigmentación de las axilas, color del follaje, color de la flor, color del pedúnculo y pedicelo, color predominante de la superficie del tubérculo, color secundario de la superficie del tubérculo, color predominante y secundario de la pulpa, distribución del color secundario de la pulpa y forma de los tubérculos (Pissard et al., 2007; Pissard et al., 2008; Rosero, 2010; Malice et al., 2007, 2009, 2010).
El análisis mediante el coeficiente de Nei-Li a un nivel de similaridad de 0.65 permitió diferenciar a los materiales de ibias en dos grandes grupos (figura 1). En el grupo I se encuentran las ibias que fueron colectadas en los municipios de Soracá, Tuta y Ventaquemada y que presentan características similares en cuanto a la floración que va de moderada a abundante, el color de la superficie del tubérculo presenta una distribución alrededor de las yemas y el color predominante de la pulpa es blanco amarillento (Morillo et al., 2016). En el grupo II a una similaridad de 0.75 hay una distribución de materiales de diferente origen geográfico, lo cual refleja el intercambio de material de siembra entre los diferentes municipios productores, lo cual hace que estas ibias no difieran mucho morfológicamente unas de otras, especialmente en características como color y forma del tubérculo, longitud y ancho de la lámina foliar, color de la pulpa sin embargo, los materiales de Combita fueron los que presentaron los menores valores de similitud en comparación con las otras ibias evaluadas, esto puede deberse a que presentan características morfológicas altamente diferenciables como hábito de floración abundante, la distribución del color secundario de la superficie de los tubérculos se ubican directamente en las yemas, el color predominante de la pulpa es blanco amarillento con una longitud y ancho de la lámina foliar de 4.3 cm y 4.8 cm, respectivamente (Morillo et al., 2016).
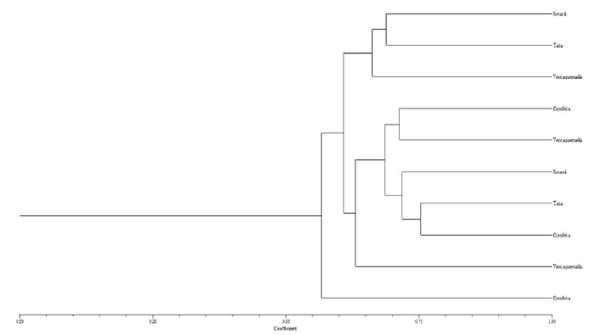
Figura 2 Dendrograma de ibias, basado en el coeficiente de similitud de Nei-Li y calculado con siete marcadores RAMs, con el método de cal con el método de clasificación UPGMA, SAHN y TREE de NTSYs-pc Versión 1.8 [Exeter Software, Y, USA].
En términos generales los agrupamientos de los materiales evaluados con marcadores RAMs y los descriptores usados por Rosero (2010), no se corresponden a los sitios geográficos done fueron colectadas las ibias, sino más bien son las características morfológicas asociadas al color de los tallos, foliolos y tubérculos, hábito de floración, entre otros los que están determinando dichos agrupamientos, características las cuales ya habían sido reportadas en otros estudios de caracterización de la diversidad genética en esta especie como altamente discriminantes (Bernal et al., 2007; Rosero, 2010). Las distancias genéticas son menores a los encontrados en otros estudios de diversidad genética en ibias (Pissard et al., 2006; Malice et al., 2007, 2010), lo cual revela un menor grado de variabilidad en los materiales evaluados.
El análisis de la caracterización molecular usando los marcadores Microsatélites Amplificados al Azar (RAMs) en ibias mostraron que el cebador CGA fue el que mayor aporte hizo a la variabilidad genética observada, Fst de 0.32, lo cual significa que es un marcador discriminante que puede ser tenido en cuenta a la hora de evaluar la diversidad genética en otras especies tuberosas (tabla 5).
Tabla 5 Valores de Heterocigosidad, Fst y Porcentaje de loci polimórficos para los cebadores RAMs evaluados en los diez materiales de ibias.
| Cebador | N° Loci | He Estimada | % Loci Polimórficos (95%) | Fst | SD |
|---|---|---|---|---|---|
| ACA | 23 | 0.39 | 95 | 0.24 | O.07 |
| AG | 19 | 0.42 | 97 | 0.26 | 0.04 |
| CA | 22 | 0.46 | 98 | 0.11 | 0.05 |
| CCA | 33 | 0.32 | 97 | 0.31 | 0.05 |
| CT | 28 | 0.42 | 85 | 0.22 | 0.04 |
| TG | 39 | 0.42 | 98 | 0.16 | 0.04 |
| CGA | 25 | 0.40 | 98 | 0.32 | 0.02 |
| TOTAL | 190 | 0,39 | 96 | 0.23 | 0.02 |
De acuerdo con otros estudios, tanto las repeticiones di y trinucleótidas son más útiles para la amplificación de las bandas polimórficas (Parra et al., 2015). Los cebadores con repeticiones AG y GA son más eficientes ya que producen patrones de bandas más definidos y polimórficos, estos motivos parecen ser más abundantes en el genoma de la ibia (Ruas et al., 2003, Malice et al., 2007).
El valor de heterocigosidad promedio para toda la población fue de 0.39, el cual puede ser bajo si se compara con los valores obtenidos en estudios de diversidad genética en otras especies tuberosas como la papa (Navarro et al., 2010; Onamu et al., 2015); pero más alto que lo encontrado por Pissard et al. (2006), al evaluar 32 accesiones de ibias (O. tuberosa M.) con marcadores ISSR (He= 0.28), y en los estudios realizados por Malice et al. (2010), en el microcentro de Huanuco en Perú (Ht= 0.33). Lo anterior muestra que existe diversidad genética en los materiales de ibias (O. tuberosa M.) colectados en el departamento de Boyacá que debe ser conservada como parte de la seguridad alimentaria. Los resultados anteriores son corroborados con el valor de Fst (0.23) el cual muestra una gran diferenciación genética, lo cual es debido al nivel de estructuración de la población como al proceso de propagación vegetativa que han venido sufriendo estos materiales ciclo tras ciclo. El análisis de varianza molecular (AMOVA) mostró que gran parte de la variación genética observada en los materiales de ibias colectados (89%) se encuentra dentro de cada grupo, lo cual indica niveles de niveles de subdivisión y jerarquización mayores a los evaluados en este estudio.
La existencia de variabilidad genética en cultivos propagados vegetativamente ya había reportado anteriormente y en el caso de la oca se atribuye al sistema de reproducción el cual es clave en la creación y conservación de la diversidad genética, las plantas han desarrollado sistemas de mejoramiento diametralmente opuestos en términos de creación por el contrario, la conservación de genotipos existente, por una parte la reproducción sexual que permite la creación de nuevos genotipos a través de la recombinación y segregación de cromosomas y la propagación clonal que asegura la conservación de los mejores genotipos (Jarvis et al., 2011). La ibia es propagada vegetativamente y tiene un sistema de incompatibilidad heteromórfico lo cual le favorece la reproducción sexual. Por lo tanto, la existencia simultánea de ambos sistemas reproductivos permite tanto la creación de nuevos genotipos como el mantenimiento de los ya existentes a través de la propagación vegetativa. Lo cual juega un papel importante en el entendimiento de la diversidad genética de esta especie, la cual está influenciada por la incorporación de individuos derivados de la reproducción sexual dentro del germoplasma que manejan los agricultores (Bonnave et al., 2014) lo cual también ya sido reportado en papa (Solanum spp.) en Perú y Bolivia (Quiros et al.,1992), yuca (Manihot esculenta Crantz) (Elías et al., 2001), ñame (Dioscorea sp.) en Benin (Scarcelli et al., 2006) y otros cultivos menores (McKey et al., 2010).
Otros factores que están influenciando la diversidad genética de los materiales de ibias es su nivel de ploidia ya que es considerada un octaploide con origen interespecífico (Malice et al., 2007); los arreglos espaciales que utilizan los agricultores en sus fincas, en los cuales existen polivariedades (mezcla de variedades) (Malice et al., 2010); el intercambio de material de siembra así como la alta variación intraespecífica ya identificada anteriormente por estudios moleculares realizados con otros genotipos de Oxalis (Emswiller, 2006). El conocer los procesos que generan variación en especies de reproducción asexual, es una herramienta importante que debe tenerse en cuenta a la hora de plantear estrategias de conservación y mejoramiento genético.
CONCLUSIONES
El análisis de caracterización morfológica y molecular en los materiales de ibias evaluados, permitieron formar dos grupos de acuerdo principalmente a características relacionadas con el color y la forma de las hojas, tallos y tubérculos.
A pesar de que la ibia es un material que se ha venido propagando vegetativamente ciclo tras ciclo, se pudo observar que existe variación a nivel fenotípico y genotípico que deber conservada y usada para la generación de nuevos y mejores materiales que respondan a los intereses de los agricultores, productores y consumidores.


