











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
El zapallo Cucurbita moschata Duchesne es una hortaliza popular para consumo en fresco en Colombia y las Américas (Ortiz et al. 2013) y está siendo adoptada en Europa de manera creciente (Andrejiová et al. 2018). Según el anuario estadístico de la Faostat para el 2017, se produjo un total de 27 millones de toneladas de cucúrbitas, incluyendo, zapallos, calabazas y calabacines, donde el 60,5% de la producción mundial se registró en Asia, con una participación del 43%, proveniente de China y del 18%, de India. Europa registró una producción total de 4 millones de toneladas: España, Francia y Alemania representaron los países con el 80% de la producción total en el continente. Por su parte, en América, se produjeron 3,5 millones de toneladas, que participaron con el 12,8% de la producción mundial; los países productores de zapallos y de calabacines, fueron: Argentina, México y Perú (FAO, 2017). El zapallo, por su valor, merece ser investigado a nivel básico, en especial, para identificar efectos de heterosis en el peso del fruto (Restrepo et al. 2018) o avanzar hacia la identificación de padres sobresalientes para producción en campo y calidad de fruto, mediante pruebas de progenie, como habilidad combinatoria por cruzamientos dialélicos.
El concepto de habilidad combinatoria fue propuesto por Sprague & Tatum (1942), como “la productividad de los cruzamientos entre una línea pura con el mayor número de líneas puras posibles”. Según Allard (1978), significó una nueva manera de hacer pruebas de progenie y, en su forma general, ha sido considerado como un método ideal para evaluar el comportamiento de líneas o variedades, cuando se usan como progenitores en combinaciones híbridas, de manera que, las líneas o variedades difieren marcadamente en su habilidad para cruzarse, en especial, porque las combinaciones híbridas son un reflejo directo de la habilidad para combinarse, por parte de los progenitores y de la expresión de la acción génica de tipo aditiva (Falconer & Mackay, 1996).
En síntesis, habilidad combinatoria general, designa el comportamiento promedio de una línea en sus combinaciones híbridas y, habilidad combinatoria específica, designa aquellos casos en que, ciertas combinaciones, tienen un comportamiento mejor o peor del que se puede esperar, sobre la base del comportamiento promedio de las líneas consideradas. La habilidad combinatoria es una herramienta importante en la indagación y propuesta de metas del mejoramiento vegetal (Ceballos, 1998; Griffing, 1956; Lobo & Marín, 1973; Vallejo & Estrada, 2013).
En los últimos años, se han realizado numerosos estudios en variables agronómicas y agroindustriales (Rodríguez et al. 2018; Restrepo et al. 2018; Valdés et al. 2017; Ortiz et al. 2015) y sobre habilidad combinatoria en especies del género Cucurbita. Restrepo et al. (2018) determinaron que en la expresión del carácter peso promedio del fruto y producción de frutos por C. moschata, la habilidad combinatoria general (HCG), en las generaciones de endogamia S0, S1 y S2, fueron altamente significativas. En el mismo sentido, en pepino Cucumis sativus, el color del fruto fue controlado por genes del tipo aditivo con evidente capacidad de habilidad combinatoria general y la forma del fruto, por genes de tipo aditivo y no aditivo (Olfati et al. 2013). Por su parte, Davoodi et al. (2016) consideraron que en los cruzamientos entre C. moschata y C. pepo, la mayoría de las características son reflejo de expresiones de habilidad combinatoria específica o de tipo heterótico.
El objetivo de esta investigación fue analizar el efecto de la endocría en la habilidad combinatoria de caracteres asociados con rendimiento y calidad del fruto en zapallo, C. moschata Duchesne.
MATERIALES Y MÉTODOS
Localización. El trabajo de campo, se realizó en dos localidades: Centro Experimental de la Universidad Nacional de Colombia, sede Palmira -CEUNP-, ubicado en el municipio de Candelaria, departamento del Valle del Cauca (Colombia), a 03° 25' 3 latitud Norte y 76° 25' 47.8” longitud Oeste, 973m s.n.m., con 26°C de temperatura promedia anual, 1.100mm de precipitación anual y 76% de humedad relativa y en las instalaciones del Servicio Nacional de Aprendizaje -SENA-, ubicado en el municipio de Buga, departamento del Valle del Cauca (Colombia), con coordenadas: 3° 53’ 3.27” latitud Norte y 76° 18’ 56.03 de longitud Oeste, a 969m s.n.m., 23°C de temperatura promedia anual, 980mm de precipitación anual y 75% de humedad relativa (Ortiz et al. 2009). La valoración macromolecular de los frutos, se realizó en los laboratorios de Nutrición Animal, Química y Semillas de la de la Universidad Nacional de Colombia, sede Palmira.
Material genético. A partir de la colección de zapallo del Programa de Hortalizas de la Universidad Nacional de Colombia, sede de Palmira, se seleccionaron 10 introducciones de zapallo, con alto contenido de materia seca en el fruto (MSF) y grado de endogamia cero (So). Se efectuó autopolinización en cada una de las 10 introducciones, para generar semilla S1. Posteriormente, se autofecundaron las flores S1, para generar semillas S2. Se seleccionaron seis líneas S2 y sus correspondientes antecesores, S1 y S2, para realizar tres sistemas de cruzamientos dialélicos: S0xS0, S1xS1 y S2xS2, siguiendo el método 2 y modelo I (6 progenitores, 15 híbridos F1, sin incluir los recíprocos), propuesto por Griffing (1956). Los híbridos F1, provenientes de los tres sistemas de cruzamiento dialélicos, se sembraron y se evaluaron en los campos experimentales de Candelaria y de Buga. Los 6 progenitores fueron: introducciones 2 y 6, procedentes de Pradera-Valle del Cauca; introducciones 28 y 34, procedentes del Patía-Cauca y las introducciones 79 y 80, de Santa Marta (Magdalena) (Ortiz et al. 2009).
Caracteres evaluados. Se evaluaron las siguientes variables: Número de frutos por planta (NFP); peso promedio del fruto en kg (PPF); peso total por planta en kg (PTP); materia seca del fruto en % (MSF); color de pulpa, empleando un abanico Roche; carotenos totales µg.g-1 (CT), mediante la técnica de extracción por éter de petróleo-acetona-lavado con agua-decantado y espectrofotometría 450nm (Rodríguez-Amaya & Kimura, 2004); almidón total en la pulpa mg.g-1 (AL), mediante gelatinización y dispersión del almidón en KOH 4M, seguida por una hidrólisis mediada por una amiloglucosidasa termo resistente y determinación específica de la glucosa, por un reactivo glucosa oxidasa y espectrofotometría a 500nm (Cumarasamy et al. 2002; Lasek et al. 2012) y proteína cruda en la pulpa del fruto (%) (PC), estimada como N total, por el método de Kjeldahl, por la constante 6,25 (Nielsen, 2017).
Diseño experimental. En cada localidad, se utilizó el diseño parcelas, divididas en bloques completos al azar, con 4 repeticiones, para generación de endocría, donde la parcela principal correspondió a generaciones y la sub parcela a los padres y los híbridos F1, resultantes de cada cruzamiento dialélico: S0xS0, S1xS1 y S2xS2. Cada genotipo dispuso de cinco plantas por repetición, para disponer de tres plantas finales, como unidad experimental.
Análisis genético. El análisis genético - estadístico, se realizó mediante la metodología propuesta por Hallauer & Miranda (1988) (Tabla 1), con el cual, se puede identificar si existen diferencias significativas entre padres, entre cruzas y entre padres y cruzas, de modo que, en segunda instancia, emplea un análisis de varianza combinado (Tabla 2), que combina las ventajas de la metodología de Gardner & Eberhart (1966) y Gardner (1967), con la de Griffing (1956).
Tabla 1 Esquema del análisis funcional de la varianza para los resultados obtenidos por localidad.
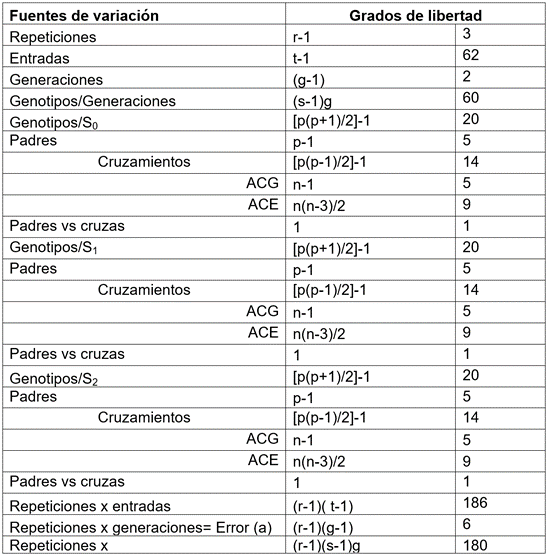
Adaptado de Hallauer & Miranda (1988).
Tabla 2 Cuadrados medios del análisis de varianza combinado para los caracteres asociados a producción por planta y calidad del fruto de zapallo.
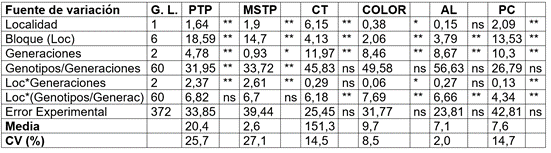
Donde: Número de frutos por planta (NFP); peso promedio del fruto en kg (PPF); peso total por planta en kg (PTP); materia seca del fruto en % (MSF); color de pulpa (Color); carotenos totales µg.g-1 (CT); almidón total en la pulpa mg.g-1 (AL) y proteína cruda en la pulpa del fruto (%) (PC). * Probabilidad del error con 5%; **; Probabilidad del error con 1%; ns= sin diferencias significativas.
Para el procesamiento de la información, se utilizó SAS-9.1 (versión Windows) SAS Institute Inc (2012) y GENES (versión Windows 2004 y 2019) (Cruz, 2019).
RESULTADOS Y DISCUSIÓN
Efecto de la endocría. En la tabla 2, se relacionan los cuadrados medios del análisis de varianza combinado, para los caracteres asociados a producción por planta y calidad del fruto. Se presentaron diferencias significativas y altamente significativas entre las tres generaciones de endocría, para todos los caracteres evaluados (P<0,05 y P<0,01). La interacción genotipo dentro generación de endocría fue altamente significativa, solamente para los caracteres producción y materia seca total por planta. La interacción generaciones de endocría por localidad fue significativa para la mayoría de los caracteres, con excepción de carotenos totales y contenido de almidón.
Lo anterior indicó, que la totalidad de los caracteres evaluados tienen un comportamiento diferencial dependiendo del nivel o grado de endocría, que puede ser aumentando, disminuyendo o permaneciendo constante su expresión fenotípica. Igualmente, las diferentes introducciones y sus cruzamientos de Cucurbita, utilizadas en el trabajo, respondieron de manera diferente a los niveles de endocría y a la localidad donde fueron evaluadas. A consideraciones similares llegó Restrepo et al. (2018), en si valoración del efecto de endocría en producción de zapallo.
El comportamiento de los genotipos, provenientes de los diferentes sistemas de cruzamientos dialelicos, depende del origen de las introducciones de Cucurbita utilizadas, del grado de endocría y de la localidad o ambiente donde se evalúen dichos genotipos y ello supone una herramienta para identificar genotipos específicos para localidades específicas, con alto grado de adaptación a un ambiente determinado (Restrepo et al. 2018). En ese mismo sentido, Vallejo et al. (2010) y Baena et al. (2010) mencionan que C. moschata responde de manera positiva a los ambientes favorables de siembra y que la respuesta al proceso de endocría, probablemente, esté influida por las condiciones de clima y de suelo de la localidad de siembra de las poblaciones básicas (S0) y sus respectivas líneas endogámicas (S1 y S2). Todo ello, sin menoscabo de depresión por endocría y, en algunos casos, “si la endogamia es una alternativa para cambiar la frecuencia genotípica de una población y, por lo tanto, la expresión del carácter en estudio (Falconer & Mackay, 1996), el efecto positivo de la endocría podría atribuirse a la alta dominancia (Ceballos, 1998) e interacciones de dominancia por dominancia, con probable presencia de dominancia remanente, a expensas de la aditividad, a medida que se acentúa la endocría, generando, de ese modo, en generaciones endogámicas tempranas, el fenómeno de epistasis sinérgica (Crow & Kimura, 1970), incrementando las variables de producción” (Ortiz et al. 2009)
Las variables producción total por planta y calidad del fruto expresaron diferencias altamente significativas (P<0,01), dependiendo del nivel de endocría (generaciones) (Tabla 2). La interacción genotipo dentro generación de endocría para los caracteres PTP y MSF expresaron diferencias altamente significativas (P<0,01), mientras que para las variables de calidad, no presentaron diferencias (Tabla 2).
Para los caracteres peso total por planta PTP y materia seca total por planta MSTP, el cuadrado medio del error asociada con la fuente de variación generaciones de endocría, fue considerablemente más alta en Buga que en Candelaria. En contraste, la magnitud del error fue 4,5 veces mayor en Candelaria que en Buga. Lo anterior, explica que pequeñas diferencias entre los promedios de generaciones S0, S1 y S2 sean declarados como no significativos en la primera localidad (Candelaria) y como significativos en la segunda (Buga), lo cual, demuestra, por un lado, que la producción total por planta varía entre generaciones de endocría y del ambiente donde se realice la evaluación (SC entre generaciones en Buga > SC entre generaciones Candelaria) y por otro, la influencia del microambiente (SC repetición x generación) y del macroambiente (SC genotipo x localidad) de siembra sobre el comportamiento de los genotipos evaluados.
Espitia et al. (2006) no encontraron diferencias significativas en producción por planta de zapallo al evaluar variedades (S0) y líneas S1 en diferentes ambientes, contrario a los resultados obtenidos por Gwanama et al. (1998) y Mohanty & Prusti (2002), quienes detectaron como significativa la interacción genotipo x ambiente (P<0,05), tal como ocurrió en el presente trabajo. Las diferencias entre los resultados de uno u otro estudio son explicables por el tipo de material genético utilizado (introducciones S0, líneas S1 y S2, híbridos F1) y por los ambientes (localidades) utilizados (El-Adl et al. 2014).
Análisis de habilidad combinatoria. De acuerdo con los resultados que se presentan en las tablas 3 y 4, se deduce que la contribución (%) de los cuadrados medios de la habilidad combinatoria a la SC de la fuente de variación híbridos, depende del ambiente de evaluación y del nivel de endogamia de los genotipos. La habilidad combinatoria permite evaluar el comportamiento de líneas o variedades, cuando se usan como progenitores en combinaciones híbridas (Hallauer & Miranda, 1988). Dicho de otra forma, la habilidad combinatoria sirve para seleccionar buenos progenitores para los diferentes programas de mejoramiento, en especial, porque es un método efectivo en la identificación de progenitores sobresalientes. La habilidad combinatoria general se debe a la presencia de efectos genéticos asociados con la varianza aditiva en la población base, de la cual, son tomados los progenitores (Falconer & Mackay, 1996).
Tabla 3 Contribución (%) de los cuadrados medios de la habilidad combinatorias general (HCG) y específica (HCE), en los híbridos provenientes de tres cruzamientos dialélicos, con diferentes niveles de endocría, evaluados en dos localidades.
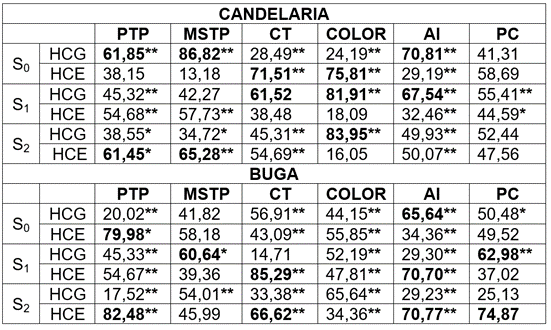
Donde: Número de frutos por planta (NFP); peso promedio del fruto en kg (PPF); peso total por planta en kg (PTP); materia seca del fruto en % (MSF); color de pulpa (Color); carotenos totales µg.g-1 (CT); almidón total en la pulpa mg.g-1 (AL) y proteína cruda en la pulpa del fruto (%) (PC). * Probabilidad del error con 5%; **; Probabilidad del error con 1%; ns= sin diferencias significativas.
Tabla 4 Contribución de los efectos de HCG y HCE en la manifestación de los caracteres evaluados, en dos localidades.
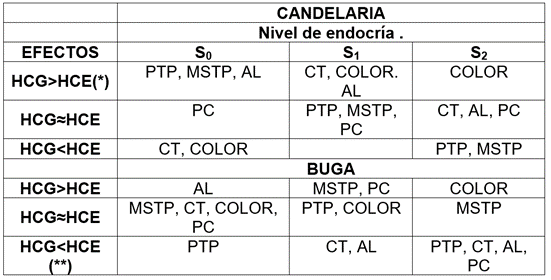
Donde: (*) = HCG> 60%; (**) = HCE> 60%. Número de frutos por planta (NFP); peso promedio del fruto en kg (PPF); peso total por planta en kg (PTP); materia seca del fruto en % (MSF); color de pulpa (Color); carotenos totales µg.g-1 (CT); almidón total en la pulpa mg.g-1 (AL) y proteína cruda en la pulpa del fruto (%) (PC). * Probabilidad del error con 5%; **; Probabilidad del error con 1%; ns= sin diferencias significativas.
En Candelaria, los híbridos entre progenitores S0 mostraron predominio de los efectos de HCG en PTP, MSTP y AL, en tanto que, para CT y COLOR, los efectos de HCE fueron más acentuados. Para PC, los dos efectos son relativamente similares en su magnitud. En Buga, con excepción del AL (HCG>HCE) y la PTP (HCG <HCE), los demás caracteres muestran una contribución relativamente similar entre los efectos de HCG y HCE.
Para los híbridos formados entre líneas S1, la HCG es superior en las variables de calidad del fruto (CT, COLOR, AL) en Candelaria, mientras que en Buga, este modo de acción génica influyó de manera similar, tanto en producción como en calidad (MSTP y PC). Los efectos de HCG y HCE presentaron contribuciones relativamente similares para las variables PTP, MSTP y PC, en Candelaria y PTP y COLOR, en Buga (Tabla 3).
Para los híbridos formados entre líneas S2 es evidente que son más importantes los efectos de HCG en COLOR para las dos localidades, en contraste con el predominio de los efectos de HCE para PTP y MSTP (Tabla 3). Los demás caracteres o presentan efectos en magnitud relativamente similar o con predominio de la HCE, dependiendo de la localidad.
Del análisis anterior, se deduce que, la HCG y HCE para los caracteres de calidad del fruto y de producción presentan un patrón de comportamiento variable y no existe, por tanto, un predomino de un modo de acción génica específica, que permita definir una única estrategia de mejoramiento para los caracteres considerados. Además, debido a la evidente interacción generación de endocría x localidad, se debe realizar selección de progenitores en las diferentes localidades consideradas. A consideraciones similares llegan Elhameid et al. (2017), trabajando con líneas endocriadas de C. pepo.
Los progenitores con efectos de Habilidad Combinatoria General (gi), significativamente mayores que cero, en una determinada generación de endocría, no preservan esta cualidad en la siguiente generación (Tabla 4) y ello supone, entonces, identificar buenos progenitores en cada generación de endocría.
En la localidad de Candelaria, por ejemplo, la población 80-S0 que posee excelente capacidad combinatoria para producción por planta y calidad de fruto (excepto CT y PC), se podría recombinar con los progenitores 6-S0 o 28-S0, para mejorar el contenido de carotenos, manteniendo o incrementando los indicadores de producción. Otra alternativa sería recombinar líneas de la población 34-S1 con la población 80-S0, con el consecuente aumento en Color, CT y MSTP.
Debido a la interacción generación de endocría por localidad es necesario seleccionar progenitores en cada localidad que presente alta HCG y altos promedios para los caracteres evaluados. En Candelaria, probablemente, el uso de líneas endogámicas S1 y S2 no sea recomendable, pero en Buga, dichas líneas serían los mejores progenitores, porque poseen los mayores efectos de HCG, para rendimiento y calidad.
Existen varios progenitores, en generaciones S0, S1 o S2, que presentan efectos de HCG, significativamente mayores que cero, que pueden ser utilizadas en progenitores de selección para incrementar los diferentes caracteres asociados con producción y calidad de frutos.
Análisis de Habilidad Combinatoria Específica y heterosis. La identificación de combinaciones híbridas con efecto de habilidad combinatoria específica (HCE), significativamente mayor que cero, para caracteres de rendimiento y calidad de fruto, abre la posibilidad de explotar el fenómeno de la heterosis, en la producción de alimentos. Los resultados que se presentan en la tabla 5 resumen información sobre el comportamiento en términos de efecto de HCE, significativamente mayor que cero; heterosis relativa media (HRM) y heterobeltiosis (HB) para las combinaciones híbridas de más alto desempeño en cada generación y localidad.
Tabla 5 Progenitores que presentaron efectos de Habilidad Combinatoria General HCG (gi) significativamente mayores que cero, en dos localidades.
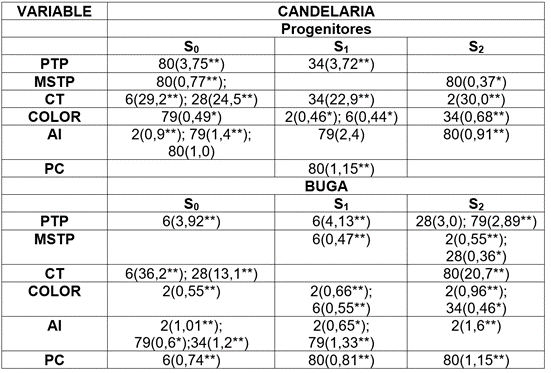
Donde: Número de frutos por planta (NFP); peso promedio del fruto en kg (PPF); peso total por planta en kg (PTP); materia seca del fruto en % (MSF); color de pulpa (Color); carotenos totales µg.g-1 (CT); almidón total en la pulpa mg.g-1 (AL) y proteína cruda en la pulpa del fruto (%) (PC). * Probabilidad del error con 5%; **; Probabilidad del error con 1%; ns= sin diferencias significativas. El valor numérico antes del paréntesis corresponde al padre, que expresa habilidad combunatoria general y el valor entre paréntesis al renqueo, que arroja el análisis de habilidad combinatoria derivado de Hallauer & Miranda, que no tiene magnitud.
Se presentó respuesta diferencial en el comportamiento de los híbridos provenientes de las introducciones a libre polinización, líneas endocriadas S1 y S2. Igualmente, los híbridos presentaron comportamiento diferencial dependiendo de las localidades en donde se evaluaron. Los híbridos provenientes de generaciones S0, se expresan de manera diferente en las localidades. Ello, supone un efecto ambiental marcado sobre los híbridos provenientes de la generación S0 y que, por lo tanto, se debería identificar un cruzamiento particular para cada localidad, según sea el interés del mejorador. En la generación S1, por su parte, sobresalieron para PTP los cruzamientos 6x34 y 80x34, por expresar HCE altamente significativa con HRM y HB (Tabla 6), sobresalientes para las dos localidades simultáneamente. Asimismo, para AL en S1, solamente el cruzamiento 2x79 expresó complementariedad genética para HCG, altamente significativa con HRM y HB, relevantes en las dos localidades simultáneamente.
Tabla 6 Efectos de Habilidad Combinatoria Específica (Sij), significativamente mayores que cero, heterosis relativa media (HRM) y heterobeltiosis (HB), para los híbridos de mejor desempeño.
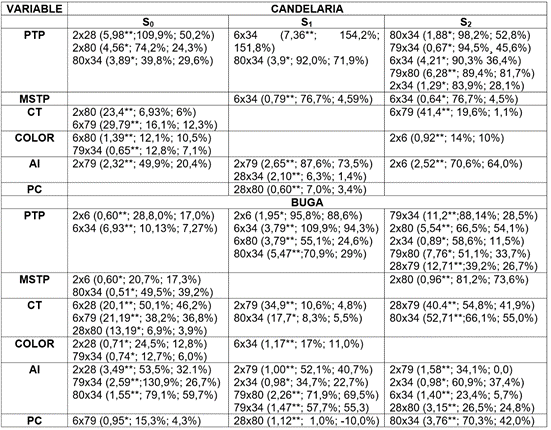
Donde: Número de frutos por planta (NFP); peso promedio del fruto en kg (PPF); peso total por planta en kg (PTP); materia seca del fruto en % (MSF); color de pulpa (Color); carotenos totales µg.g-1 (CT); almidón total en la pulpa mg.g-1 (AL) y proteína cruda en la pulpa del fruto (%) (PC). * Probabilidad del error con 5%; **; Probabilidad del error con 1%; ns= sin diferencias significativas. El valor numérico antes del paréntesis corresponde a los padres que concurre en el cruzamiento y el valor entre paréntesis al renqueo, que arroja el análisis de habilidad combinatoria específica, derivado de Hallauer & Miranda, que no tiene magnitud y corresponden a: Habilidad Combinatoria Específica (Sij) significativamente mayores que cero, heterosis relativa media (HRM) y heterobeltiosis (HB), para los híbridos de mejor desempeño (%) respectivamente, dentro del paréntesis.
En la generación S2, probablemente, se haya presentado mejor complementariedad genética para la expresión de HCE, altamente significativa con HRM y HB relevantes para PTP, posiblemente, porque la endocría acumuló genes favorables. Ello, se confirma con la respuesta positiva y simultánea en las dos localidades de los cruzamientos 2x34, 79x34 y 79x80 para PTP. De todo esto, se desprende que si la decisión del programa de mejoramiento genético es producir híbridos, la estrategia sería concentrar genes favorables para rendimiento y calidad, a través de la formación de híbridos dobles o triples; por ejemplo, para condiciones de Candelaria, se sugiere la formación de los siguientes híbridos dobles o triples: (2x80-S0 x 79x34-S0), (2x28-S0 x 79x34-S0), (28x80-S0) x 2-S0, (79x34-S0) x 80-S0, (80x34-S0) x 79-S0 y para las condiciones de Buga, se sugiere la formación de los siguientes híbridos dobles o triples: (6x34S1 x 79x80S1); (2x6S1 x 79x34S1); (79x80S1) x 34S1; (6x34S1) x 80S1. Para la generación S2, dado que presentó híbridos con alta HCE, HRM y HB, en las dos localidades, entonces, la estrategia, probablemente, sería concentrar genes para rendimiento y calidad del fruto, mediante el siguiente híbrido doble: (79x34S2 x 2x6S2).
De todo lo anterior y a modo de corolarium, se deben seleccionar los mejores híbridos para cada localidad, debido a la interacción significativa generación de endocría por localidad; además, los caracteres relacionados con la calidad del fruto fueron más sensibles (negativamente) al efecto de endocría que los caracteres relacionados con el rendimiento.
La habilidad combinatoria general de un progenitor (g i ) y la habilidad combinatoria específica (S ij ) de un cruzamiento, en caracteres asociados con calidad del fruto y rendimiento, presentaron comportamientos diferenciales, dependiendo de la generación de endocría y localidad de evaluación, donde los progenitores de mejor desempeño para los caracteres relacionados con producción, fueron: 80S0; y 34S1, en Candelaria y 6S0; 6S1; 28S2; 79S2, en Buga y para caracteres de calidad de fruto (en especial, almidón), fueron: 2S0; 79S0; 79S1; 80S2, en Candelaria y 2S0; 79S0; 34S0; 2S1; 79S1 y 2S2, en Buga.
Aunque en Candelaria no fue posible identificar un híbrido con efecto de HCE alto para todos los caracteres asociados con rendimiento y calidad, se destaca algunas combinaciones, como 2x28-S0, 2x80S0, 28x80-S0 y 80x34-S0 y para las condiciones de Buga, no se identificaron híbridos derivados de S0, S1 y S2, que tuviesen un buen desempeño para rendimiento y calidad. Las combinaciones 2x6-S1, 80x34-S1, 2x80-S2 y 79x34-S2 sobresalieron únicamente por la HCE para rendimiento.
Parece que el uso de líneas endogámicas S2 es la estrategia más adecuada para producción de híbridos, debido a la expresión de HCE altamente significativa con HRM y HB, relevantes para PTP. Ello, se confirma con la respuesta positiva y simultánea en las dos localidades de los cruzamientos 2x34, 79x34 y 79x80, para PTP.

