











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
INTRODUCCIÓN
El cacao (Theobroma cacao L.) es un cultivo tropical originario de la cuenca Amazónica, abunda en los bosques húmedos de Colombia (González-Orozco et al. 2020), donde predominan suelos ácidos (Quesada et al. 2009). Allí crece el cacao, aunque estas condiciones de acidez (pH<5,5) disminuyen la disponibilidad y la absorción de nutrientes (Rosas-Patiño et al. 2017) y limita la concentración en los tejidos (Quinteiro et al. 2013), afectando el desarrollo de las plantas y, por consiguiente, su rendimiento.
El encalado aumenta el pH y la fertilidad en los suelos, mejora la relación suelo-planta e incrementa la absorción y concentración de nutrientes en la planta (Schlindwein & Gianello, 2004); esta concentración de nutrientes en un órgano especifico depende de la interacción entre ambiente, genotipo y manejo del cultivo (Correndo & García, 2012).
Los análisis de tejidos permiten diagnosticar el contenido y la cantidad de nutrientes que extrae el cultivo (Marrocos et al. 2020a) y su distribución en hoja, cáscara y grano, información de gran relevancia para mitigar el impacto de la excesiva fertilización y hacer un eficiente manejo nutricional.
Existen algunas investigaciones acerca de la concentración de nutrientes en cacao, entre ellos, a nivel internacional, Abreu (1996), Malavolta et al. (1997) y Sodré (2002), en Brasil; Aikpokpodion (2010), en África y, más recientemente, Furcal-Beriguete (2017), en Costa Rica; asimismo, a nivel nacional, Puentes-Páramo et al. (2016a; b), sin embargo, son escasos los reportes relacionados con el efecto de los cambios del pH del suelo sobre la extracción de nutrientes en clones de cacao en la Amazonia colombiana.
El objetivo de este trabajo fue evaluar el efecto del pH en un suelo ácido (Typic Udorthents), sobre la concentración de nutrientes en hoja, cáscara y grano, para cuatro clones de cacao, dos autocompatibles (ICS-1, CCN-51) y dos autoincompatibles (ICS-39, TSH-565).
MATERIALES Y MÉTODOS
La investigación, se desarrolló en un Typic Udorthents, localizado en el paisaje de lomerío del municipio de Puerto Rico, Departamento de Caquetá, Colombia. La zona, se encuentra en condiciones de Bosque Húmedo Tropical (Ruíz et al. 2007), a 1°52´12” Latitud Norte y 75°12´24” Longitud Oeste, a 250m s.n.m. La región presenta valores medios anuales de precipitación de 2.960mm, temperatura de 25,3°C y humedad relativa de 84% (IDEAM, 2017).
Se utilizaron cuatro clones de cacao de uso comercial, dos autocompatibles (ICS-1, CCN-51) y dos autoincompatibles (ICS-39, TSH-565), establecidos durante siete años, en el Jardín Clonal de la Asociación Departamental de Productores de Cacao y Especies Maderables del Caquetá (ACAMAFRUT).
Se estableció un diseño factorial con cuatro clones (factor A), cuatro tratamientos definidos por el encalado y la fertilización del suelo (factor B) y cuatro repeticiones. Los tratamientos fueron (Tabla 1): T0: testigo o suelo con fertilidad natural; T1: disponibilidad de nutrientes después de 90 días de aplicar 7t ha-1 de cal dolomita, para llevar el suelo a pH≥5,5 (Rosas-Patiño et al. 2017); T2: aplicación de N-P-K en T0, hasta alcanzar los requerimientos del cultivo (Puentes-Páramo et al. 2014) y T3: aplicación de N-P-K en T1, hasta alcanzar los requerimientos del cultivo propuestos por los precitados autores.
Tabla 1 Valor de pH y concentración de nutrientes por tratamiento.
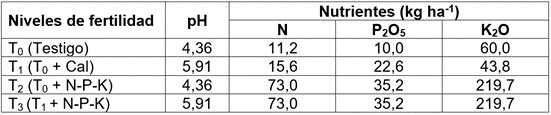
N: nitrógeno; P2O5: pentóxido de fósforo; K2O: óxido de potasio.
Para garantizar el efecto de los tratamientos en las unidades experimentales, se omitió la fertilización durante un año y se eliminaron los frutos para estandarizar la producción. Posteriormente, la fertilización para cada tratamiento, se fraccionó en dos momentos; uno, al inicio de la época lluviosa, en marzo-abril y, otro, en agosto. La unidad experimental estuvo conformada por cuatro árboles sembrados a 3mx3m, para una densidad poblacional de 1.111 plantas por hectárea.
Los muestreos foliares y de fruto, se realizaron para el año cacaotero 2016-2017. En cada tratamiento y por cada clon, se colectó, de la parte media de cada árbol en plena floración, la cuarta hoja (en total, 25 hojas). Además, se colectaron siete mazorcas maduras y sanas, para determinar la concentración y extracción de nutrientes, por 1.000kg de grano seco. Para estimar la extracción, se tuvo en cuenta el tratamiento con mayor rendimiento obtenido en cada clon, de acuerdo con Rosas-Patiño et al. (2019). Las muestras de hoja, cáscara y grano, se secaron en estufa, a 103°C, durante 24 horas; se molieron y se enviaron al laboratorio de suelos de Agrosavia (Tibaitatá, Colombia) y se determinó el contenido de nitrógeno (N), por EPA 351,3; de fósforo (P), por espectrofotometría; de potasio (K), calcio (Ca) y magnesio (Mg), por espectrofotometría de emisión de plasma inductivamente acoplado; de azufre (S), por turbidimetría; el hierro (Fe), cobre (Cu), manganeso (Mn) y zinc (Zn), se determinaron por absorción atómica, mientras que el boro (B), por ignición.
Los datos fueros sometidos a un Test de normalidad, mediante la prueba Shapiro-Wilks; las variables con comportamiento normal, se evaluaron mediante análisis de varianza, para determinar el efecto de los tratamientos y las interacciones; el ANAVA, para los datos con comportamiento no paramétrico, fue realizada mediante la prueba Kruskal Wallis. Se realizaron pruebas de comparación de medias (Tukey) y correlaciones de Pearson entre variables, con p≤0,05, utilizando el programa InfoStat-Profesional, versión 2020p (Di Rienzo et al. 2020).
RESULTADOS Y DISCUSIÓN
El análisis de varianza (Tabla 2) evidenció efectos altamente significativos en la interacción clon*tratamiento en todos los tejidos, para K y micronutrientes, con excepción de Cu, que no presentó diferencias en la concentración foliar. N presentó efecto (p<0,01), en cáscara y hoja (p<0,05) y Ca, en hoja (p<0,01).
Tabla 2 ANAVA de la concentración de nutrientes en hoja (H), cáscara (C) y grano de cacao (G), por efecto del clon, tratamiento e interacción.
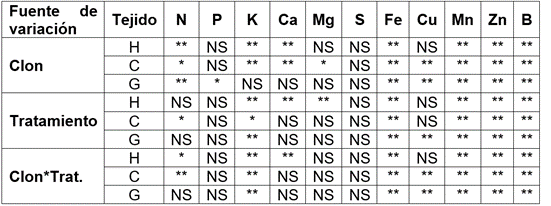
**diferencias altamente significativas (p<0,01); *diferencias significativas (p<0,05); NS diferencias no significativas (p>0,05).
El Mg presentó diferencias (p<0,01) solo en el factor tratamiento. Por su parte, el factor clon evidenció diferencias altamente significativas para N, en grano y Ca, en cáscara; del mismo modo, los clones presentaron efectos significativos en P almacenado en grano y Mg, en cáscara. Las concentraciones de P, en hoja y cáscara; Ca y Mg, en grano; Cu, en hoja y S, en todos los tejidos, no se vieron afectadas por ningún factor. Los resultados sugieren un efecto de los cambios de pH y la disponibilidad de nutrientes en el suelo en la absorción de nutrientes en clones de cacao, como lo reportan Baligar & Fageria (2005), van Vliet et al. (2015) y Rosas-Patiño et al. (2019).
Concentración de macronutrientes en hoja, cáscara y grano de cacao. En general, se observó mayor concentración N, P y Mg, en grano; K, en cáscara; Ca y S, en hoja. Se evidenció, además, el efecto del clon y las condiciones del suelo en la extracción de nutrientes.
Con relación a la acumulación de nutrientes en el tejido foliar, las mayores cantidades de N se concentraron en grano, probablemente, debido a la necesidad de los genotipos, para favorecer la producción de la semilla (Nehe et al. 2020), seguido de hoja y cáscara, en menor proporción (Figura 1). Este comportamiento es normal en cacao, cuando las tasas de asimilación de N son más altas en hojas, se ve afectada la producción de los granos (Marrocos et al. 2020b).
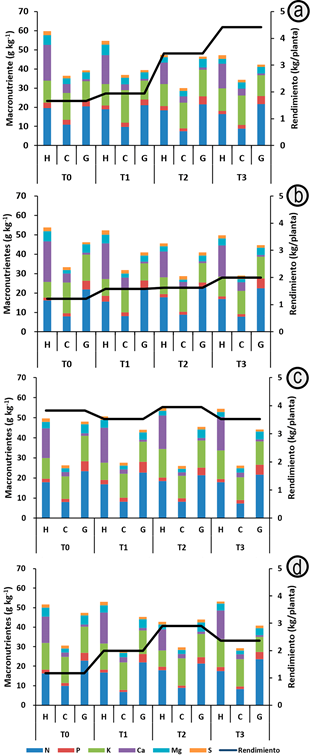
Figura 1 Concentración de macronutrientes en hoja (H), cáscara (C), grano (G) y rendimiento, para cuatro clones de cacao. a) ICS-1; b) ICS-39; c) CCN-51; d) TSH-565.
La mayor concentración foliar de N se presentó en T0 (19,57g kg-1), para ICS-1 (Figura 1A) y en T2, para los demás clones, superando los niveles reportados por van Vliet et al. (2015); probablemente, las mayores concentraciones N foliar, en los tratamientos en condiciones de acidez, se debe a las altas concentraciones de Mn, que limitan la redistribución de N a los demás órganos de la planta (Lavres et al. 2010).
Respecto al P en hoja, el clon ICS-39 (Figura 1B) presentó significativamente el mayor valor en T1 (2,97g kg-1), en donde mejoró la disponibilidad de este elemento en la solución del suelo, producto del encalado, como lo sugieren Anikwe et al. (2016); las mayores concentraciones de K en hoja, se observaron en el clon CCN-51, para T2 (14,27g kg-1) y en T3, para el resto de clones. El Ca registró el mayor valor en el tejido foliar del clon ICS-39, en T0 (20,93g kg-1), dado que, en suelos ácidos, se incrementa la absorción de Ca2+, proporcionalmente a las concentraciones de H+ y Al3+, como mecanismo de tolerancia al estrés abiótico (Bojórquez et al. 2014). La mayor extracción de Mg, la presentó ICS-1 (5,57g kg-1), en T1, valor considerado normal, dada la mayor disponibilidad y la absorción de este nutriente, en suelos encalados, como lo sugieren Ramírez et al. (2005); la mayor extracción de S (2,23g kg-1), la alcanzó ICS-39, en T1. Estos datos son superiores a los reportados por Puentes-Páramo et al. (2016b), quienes evaluaron las extracciones de clones de cacao en zonas de menor temperatura y precipitación media anual y mayor altura sobre el nivel del mar (1.129m más alta); probablemente, los contrastes en las condiciones agroclimáticas de los sitios de estudio resultaron un factor importante en estas diferencias.
En este sentido y tal como se observa en la figura 1, el orden descendente en la concentración de nutrientes foliares para cada clon, en el tratamiento de mayor rendimiento, fue: CCN-51 (T1) Ca>N>K>Mg>P>S, ICS-1 (T2) y TSH-565 (T2) N>K>Ca>Mg>P>S e ICS-39 (T3) N>Ca>K>Mg>S>P. En estos tratamientos, las concentraciones de Mg, P y S conservaron el mismo orden en todos los clones, salvo ICS-39, donde la concentración de S fue mayor a P, en contraste con N, K y Ca. Asimismo, se evidenció el efecto de condiciones edáficas sobre el genotipo, especialmente, para CCN-51 (Figura 1C), dado que este clon concentró mayor cantidad de Ca en lugar de N, lo contrario (N>Ca) sugieren Puentes-Páramo et al. (2014). Marschner (2012) afirma que Calcio es un importante mensajero para la respuesta fisiológica de la planta para adaptarse a condiciones ambientales adversas (como la acidez de los suelos), por tanto, es probable que la habilidad de CCN-51 para extraer Ca, corresponda a un mecanismo de respuesta y adaptación del clon a los ambientes tropicales.
La mayor concentración de N para cáscara se presentó en T0 (10,93g kg-1), en el clon ICS-1 y en T2, para los demás clones, excepto, TSH-565 (Figura 1D); este mismo clon presentó la menor concentración en T0 (9,8g kg-1). Respecto a P, el clon ICS-1 presentó el mayor valor en T0 (2,53g kg-1), el resto de clones en T1, exceptuando el clon TSH-565, que extrajo la menor cantidad (1,43g kg-1) en T0. La mayor concentración de K en cáscara (17,13g kg-1), se encontró en ICS-1, para T1 y la menor (11,87g kg-1), el clon CCN-51, en este mismo tratamiento.
Con relación al Ca (Figura 1), el mayor valor (4,67g kg-1), se registró para ICS-1, en T0 y menor para CCN-51, en T3 (2,33g kg-1); la mayor y menor absorción de Mg, se obtuvo en el T1, para el clon ICS-1 (3,40g kg-1) e ICS-39 (2g kg-1), respectivamente y para el S, el T1 del clon ICS-39, reportó el mayor valor (1,80g kg-1). Estas concentraciones de N, P, K, Ca y Mg fueron mayores a las refereridas por Furcal-Beriguete (2017). La secuencia de concentración de nutrientes en cáscara en el tratamiento de mayor rendimiento fue (Figura 1): CCN-51 K>N>Ca>P>Mg>S, ICS-1 K>N>Ca>Mg>P>S, TSH-565 K>N>Ca>Mg>P>S e ICS-39 K>N>Ca>Mg>P>S; las concentraciones de macronutrientes conservaron el mismo orden, excepto Mg y P, mientras la mayor concentración en cáscara corresponde a K, como lo reportaron Puentes-Páramo et al. (2014).
La mayor acumulación de N en grano (23,57g kg-1) la alcanzó el clon TSH-565 (Figura 1D), en T3, consecuente con mayor disponibilidad de nutrientes en este tratamiento, debido al encalado y la fertilización; CCN-51 logró almacenar la mayor cantidad de P (5,47g kg-1), en T1 mientras que las concentraciones de K más altas (13,9g kg-1), las alcanzó ICS-1, en T3. Con relación a Ca, se registró la mayor concentración (1,33g kg-1) en el clon TSH-565, en T1, mientras que la mayor y menor concentración de Mg (3,40g kg-1 y 3,07g kg-1 en los clones ICS-1 e ICS-39, respectivamente), se alcanzaron luego del encalado (T1); el S, se almacenó en mayor cantidad (1,80g kg-1) en el clon ICS-39, en condiciones de T1 (Figura 1).
Las concentraciones de N, P, K, Ca y Mg fueron mayores comparadas con Furcal-Beriguete (2017). Los clones ICS-1 y TSH-565 (Figura 1A y D) mostraron el siguiente orden en la concentración de N>K>Mg>P>Ca>S y los clones ICS-39 y CCN-51, presentaron N>K>P>Mg>Ca>S (Figura 1B y C), encontrando la concentración de N y S con mayor y menor valor, respectivamente, generalizada para los cuatro clones; además, coincidió con la menor concentración de S, en hoja y en cáscara.
En síntesis, la concentración de N y P fue mayor en grano>hoja>cáscara, dado que fósforo se acumula en semillas, como lo propone Marschner (2012); K presentó la mayor concentración en cáscara, seguido hoja y por último grano; Ca fue mayor en hoja>cáscara>grano; Mg en hoja>grano>cáscara y S hoja>cáscara>grano. En este orden de ideas, las hojas presentaron mayor concentración de Ca, Mg y S, comparadas con la cáscara y el grano; a su vez, la cáscara concentró mayor K y el grano mayor N y P. El clon CCN-51 evidenció mejor comportamiento agronómico puesto que, con valores similares en la concentración de macronutrientes en los tres tejidos, presentó el mayor rendimiento (Figura 1C); así también lo sugieren Cuenca-Cuenca et al. (2019).
Concentración de micronutrientes en hoja, cáscara y grano de cacao. La concentración de micronutrientes (Fe, Mn, Zn y B) en los tres tejidos (Figura 2), siguió, de manera general, el siguiente orden: hoja>cáscara>grano, correspondiente con el rango de 66-73%, 15-19% y 11-15%, respectivamente, salvo Cu, que se concentró, principalmente, en grano, dado que este elemento es esencial para la producción del fruto y la semilla (Marschner, 2012); en este sentido, la mayor concentración de Fe foliar (192,25mg kg-1), se presentó en TSH-565 (Figura 2D), en T2 ,tratamiento con alta disponibilidad de cationes ácidos en el suelo, al igual que en el T0; Cu, en CCN-51 (7,81mg kg-1), en T3, Mn, en T0 (1253,45mg kg-1), para el clon ICS-39, concentración considerada tóxica para cacao (Baligar et al. 2001).
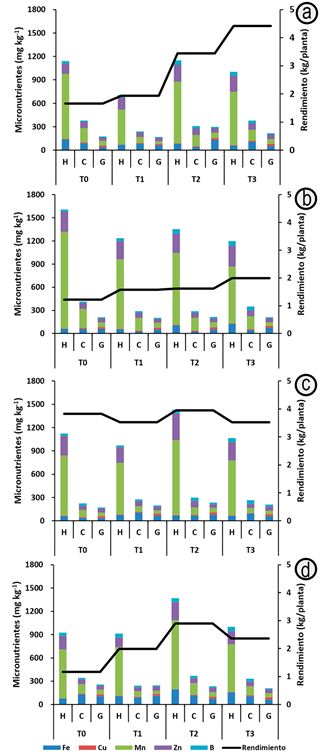
Figura 2 Concentración de micronutrientes en hoja (H), cáscara (C), grano (G) y rendimiento, para cuatro clones de cacao: ICS-1 (A); ICS-39 (B); CCN-51 (C); TSH-565 (D).
En el clon ICS-1, para T1 (Figura 2A), se presentó el menor valor de Mn (443,59mg kg-1). Es probable que el incremento de pH, Ca2+ y Mg en el suelo disminuyeran las concentraciones de Mn en los tejidos de cacao (Anda et al. 2013); no obstante, este valor continúa siendo tóxico (Hänsch & Mendel, 2009). Las cantidades de Zn registradas en CCN-51 (Figura 2C), para T2 (341,18mg kg-1), son consideradas fitotóxicas, dado que excede el límite de 244mg kg-1, sugerido por De Oliveira et al. (2015), al igual que B, en T3 (60,26mg kg-1), para el clon ICS-39 (Figura 2B); comparadas con la referencia de Puentes-Páramo et al. (2016b), con relación a otros micronutrientes, las cantidades extraídas se consideran normales para Fe, bajas para Cu y altas para Mn, Zn y B.
La concentración de micronutrientes, en el tratamiento de mayor rendimiento, fue (Figura 2): CCN-51(T1) Fe>Mn>Zn>B>Cu, ICS-1 (T2) Mn>Zn>Fe>B>Cu, TSH-565 (T2) Mn>Fe>Zn>B>Cu e ICS-39 (T3) Mn>Zn>B>Fe>Cu, coincidiendo en presentar la mayor concentración de Fe o Mn, debido a la mayor disponibilidad y absorción de estos nutrientes, favorecida por la acidez del suelo, como lo sugieren Ramírez et al. (2005). Las menores concentraciones fueron para B y Cu, resultados similares a los obtenidos por Puentes-Páramo et al. (2016a).
En cáscara, la mayor concentración de Fe (129,19,25mg kg-1), se presentó para el clon TSH-565, en T0; Cu, en CCN-51, en T1 (13,65mg kg-1); Mn, en T0 (250,13mg kg-1), para el clon ICS-39; Zn, en CCN-51, para T2 (82,23mg kg-1) y B, en T3 (49,21mg kg-1), clon CCN-51. En orden descendente, la concentración de micronutrientes en el tratamiento con mayor rendimiento fue (Figura 2): CCN-51: Fe>Mn>Zn>B>Cu, ICS-1: Mn>Zn>Fe>B>Cu, TSH-565: Fe>Mn>Zn>Cu>B e ICS-39: Mn>Zn>B>Fe>Cu, coincidiendo en presentar la mayor concentración de un catión ácido, sea Fe o Mn, favorecida por la acidez natural del suelo, como lo sugieren Rosas-Patiño et al. (2017); las menores concentraciones fueron para B y Cu, resultados similares obtuvo Furcal-Beriguete (2017).
En grano, la mayor concentración de Fe (131,42mg kg-1), se presentó para ICS-1, en T2; Cu, en TSH-565, en T3 (29,48mg kg-1); Mn, en T2 (68,56mg kg-1), en ICS-1; Zn, en TSH-565, para T2 (55,70mg kg-1) y B, en T3 (25,37mg kg-1), clon CCN-51. La concentración de micronutrientes en el tratamiento que presentó el mayor rendimiento fue (Figura 2): clon CCN-51 (T1) Fe>Zn>Mn>Cu>B, ICS-1 (T2) Fe>Mn>Zn>B>Cu, TSH-565 (T2) Fe>Mn>Zn>Cu>B e ICS-39 (T3) Fe>Zn>Mn>Cu>B, coincidiendo en presentar la mayor concentración de Fe, favorecida por la acidez del suelo, como lo sugieren Ramírez et al. (2005). Del mismo modo, se presentaron las menores concentraciones de Cu y B, para todos los clones en estos tratamientos, dado que a pH superiores a 5,4, se afecta la disponibilidad y absorción (Ambrosini et al. 2015).
La concentración de Fe, Mn, Zn y B en orden descendente para los tres tejidos fue hoja>cáscara>grano, mientras Cu, se almacenó en grano>cáscara>hoja. Lewis et al. (2021) reportaron una correlación positiva entre la disponibilidad de Fe, Mn y Zn en el suelo y las cantidades acumuladas en hojas de cacao. En este orden de ideas, las hojas presentaron mayor concentración de micronutrientes y salvo de Cu; en contraste, el grano concentró mayor Cu y la cáscara presentó la menor concentración de Fe. Además, fue evidente la eficiencia en el uso de nutrientes del clon CCN-51 (Figura 2C), dado que, con menor concentración de micronutrientes en los tejidos, presentó el mayor rendimiento, datos que concuerdan con lo reportado por Cuenca-Cuenca et al. (2019).
Correlación entre la concentración de nutrientes y pH del suelo. Las correlaciones entre la concentración de nutrientes en hoja, cáscara y grano de cacao, por efecto del pH (Tabla 3), evidenciaron diferencias entre clones y puede estar relacionado con la diferentes vías de distribución de nutrientes entre hoja-cáscara-grano de cacao (Quintino et al. 2020); los clones presentan correlaciones positivas para P, K, Mg, S, Cu y negativas para Mn en hoja; para cáscara coincidieron en correlaciones positivas para P, Ca, Mg, S, B y negativas para Mn y Zn y el grano presentó correlaciones positivas para N, P, Ca, Mg, S, Cu y negativas para K y Mn. De acuerdo con lo anterior, los tres tejidos concentran mayor P, Mg, S, cuando se eleva el pH del suelo y cuando es ácido, concentran mayor cantidad de Mn.
Tabla 3 Correlaciones de Pearson entre la concentración nutrimental en hoja, cáscara, grano seco, para cuatro clones de cacao.
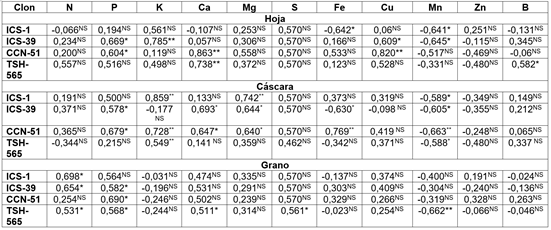
** Correlación altamente significativa al nivel 0,01, * correlación significativa al nivel 0,05, NS no significativa.
Extracción de nutrientes por 1000kg de grano seco de cacao. Los clones ICS-1 y TSH-565 coincidieron en las cantidades de macronutrientes extraídos, salvo en los contenidos de S, al igual que se observan las mismas preferencias por Mn, Zn y B, pero difieren en la asimilación de Fe y Cu. Por su parte, CCN-51 e ICS-39 extraen similares cantidades de nutrientes, con excepción de K, dado que CCN-51 es el clon que menos cantidades requiere de este elemento (Tabla 4). Los registros de extracción de N, P y K son menores a los reportadas por Quintino et al. (2020), particularmente, las cantidades de K extraídas por CCN-51. Lo anterior puede corresponder a la habilidad del clon para adaptarse a condiciones de extrema acidez y menor fertilidad natural, sin presentar estrés abiótico (Marschner, 2012).
Tabla 4 Extracción de nutrientes en kg por 1.000kg de grano seco, para cuatro clones de cacao.
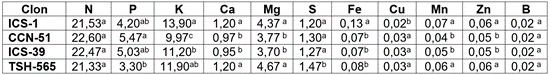
Valores por columna con letras iguales no difieren en forma significativa (P <0,05), según prueba de Tukey.
Las extracciones de micronutrientes (Tabla 4) difieren a las reportadas por Furcal-Beriguete (2017), aunque en otros genotipos, lo cual, evidencia la selectividad en la extracción de micronutrientes entre clones.
Se observó el siguiente orden de extracción de nutrientes en grano (Tabla 4): ICS-1: N>K>Mg>P>Ca=S>Fe>Mn>Zn>Cu=B; CCN-51 e ICS-39: N>K>P>Mg>S>Ca>Fe>Zn>Mn>Cu>B; TSH-565: N>K>Mg>P>S>Ca>Fe>Mn=Zn>Cu>B, siendo estos valores y preferencias diferentes y evidenciando el efecto genotipo, como lo sugieren Marrocos et al. (2020b). La secuencia de extracción de los clones, en general, para grano fue: N>K>P>Mg>S>Ca>Fe>Mn-Zn>Cu>B, ligeramente diferente con los reportes de Furcal-Beriguete (2017).
Los resultados obtenidos permiten concluir que el cambio de pH edáfico y la fertilización afecta la concentración de nutrientes de manera diferencial entre los clones evaluados, a pesar de presentar comportamiento similar en la absorción de nutrientes; CCN-51 alcanza mayor rendimiento de cosecha, incluso, en suelos con pH extremadamente ácidos. A pesar de las diferencias, los clones presentan las mayores concentraciones de N, P, Mg y Cu asociadas a grano, Ca, Fe, Mn, Zn y B asociadas a hoja y K, preferiblemente, en cáscara; asimismo, concentran mayor P, Mg, S, cuando pH≥5,5 y mayor Mn, cuando el suelo se encuentra en condiciones de acidez.

