











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
La habichuela es la legumbre inmadura del frijol común Phaseolus vulgaris L y una de las hortalizas más importante en la dieta humana (Reis Filgueira, 2013). Se consume de manera integral (legumbre con semillas vestigiales a mediano desarrollo) y es fuente importante de vitaminas, minerales y otros compuestos vegetales secundarios; en Colombia, el uso de la habichuela va en ascenso, con demanda en los mercados locales y en la gastronomía vernácula colombiana (Barraza-Alvarez, 2021). A nivel mundial, se producen, aproximadamente, 27 millones de toneladas de habichuelas (FAO, 2021).
Por otro lado, Estados Unidos tiene el 71 % de la producción mundial de habichuelas y el consumo per cápita al año es de 3 kg (USDA, 2019).
En el 2019, la producción de habichuela en Colombia fue de 45.806 ton y los departamentos de Cundinamarca, Valle del Cauca, Santander, Huila y Tolima son los principales productores (MADR, 2021); sin embargo, la producción por hectárea de este cultivo ha venido decreciendo por diversas causas, entre las que se pueden enumerar: el uso de un único cultivar, el Blue Lake y la escasa rotación de cultivos o problemas fitosanitarios del cultivo. La baja productividad hace que su producción no sea rentable para el horticultor. En este sentido, la obtención de cultivares con mayor producción constituye una prioridad para los programas de mejoramiento genético en esta especie; sin embargo, el desarrollo de un programa de mejoramiento necesita información sobre la naturaleza de las acciones de los genes, que controlan los caracteres relacionados con la producción de habichuela.
Los cruzamientos dialélicos son los sistemas de apareamiento más utilizados, que permiten identificar progenitores e híbridos prometedores, además permiten obtener información sobre el control genético de los caracteres de interés, lo que hace más eficientes los procesos de mejoramiento (Cruz et al. 2004). A partir del análisis dialélico, se recopila información sobre los efectos, debido a la habilidad combinatoria general (HCG) y la habilidad combinatoria específica (HCE) de los progenitores en los cruzamientos (Yanchuk, 1996; Glover et al. 2005). La estimación de los efectos de la HCG y la HCE permiten estimar la acción de genes aditivos y no aditivos (Griffing, 1956). El efecto de la HCG ayuda a identificar progenitores superiores, mientras que el efecto de la HCE estima el rendimiento de los híbridos (Cruz et al. 2004).
El objetivo de la presente investigación fue conocer la acción génica que controla la producción de habichuela y los caracteres relacionados a su calidad.
MATERIALES Y MÉTODOS
Material genético. Material genético de la primera etapa: se formó la población dialélica. Se emplearon seis genotipos, dos cultivares comerciales: (1) G17723 (Blue Lake de la compañía Ferry-Morse Estados Unidos) y (6) G51158 (UNAPAL-Milenio de la Universidad Nacional de Colombia Sede Palmira) y cuatro líneas: (2) G23956, (3) G20400, (4) G18212 y (5) G16806, de la colección mundial de fríjol de la Unidad de Recursos Genéticos del Centro Internacional de Agricultura Tropical (URG-CIAT), previamente seleccionadas por el Programa de Hortalizas de la Universidad Nacional de Colombia- Sede Palmira. Material genético de la segunda etapa: se evaluaron los 21 genotipos que conformaron el dialelo. Seis progenitores: (1) G17723; (2) G23956; (3) G20400; (4) G18212; (5) G16806 y (6) G51158 y 15 híbridos directos sin recíprocos: 1x2, 1x3, 1x4, 1x5, 1x6, 2x3, 2x4, 2x5, 2x6, 3x4, 3x5, 3x6, 4x5, 4x6 y 5x6.
Localización. El trabajo se realizó en dos etapas. La primera etapa consistió en la obtención de los híbridos F1, la cual, se llevó a cabo durante el segundo semestre del 2018, en la granja Mario González Aranda (MGA), de la Universidad Nacional de Colombia -sede Palmira, departamento de Valle del Cauca, ubicada a 3°30'26,8” N 76°18'47,6” O, a una altura de 1.000 m s.n.m., con temperatura media anual de 24 °C, precipitación anual de 1.020 mm y humedad relativa del 72 %. La segunda etapa estuvo relacionada con la evaluación de los progenitores y sus respectivos híbridos F1, que se desarrolló en el segundo semestre del 2019 (Ambiente1) y primer semestre del 2020 (Ambiente 2), en el Centro Experimental de la Universidad Nacional de Colombia -sede Palmira (CEUNP), localizado en la vereda el Carmelo del municipio de Candelaria, Valle del Cauca, a 3°25’34” N 76°25’53” O, a una altura de 1.000 m s.n.m., con temperatura media anual de 26 °C, precipitación anual de 1.028 mm y humedad relativa del 75 % (Ortiz Grisales et al. 2020).
Variables de respuesta evaluadas. Número de vainas por planta (NVP): al momento de la cosecha, manualmente, se determinaron las vainas que tenían, por lo menos, una semilla viable y se realizó el conteo de estas en cada planta muestreada. Producción de vainas por planta (PPP): se tomó la totalidad de las vainas por planta y se pesaron en una balanza digital; los resultados se expresaron en kilogramos. Peso promedio de la vaina (PPV): cociente entre producción de vainas por planta sobre el número de vainas por planta y se expresó en gramos. Longitud de vaina (LDV): se tomaron diez vainas por unidad experimental, se midió en centímetros, desde su inserción en el pedicelo hasta el extremo libre del ápice.
Diseño experimental. Se utilizó el diseño experimental de bloques completos al azar (Baena, 2012) con cuatro repeticiones. La unidad experimental consistió en ocho plantas, en surcos de 8 m de largo, con una distancia entre surco de 1,50 m y 1,0 m entre plantas, donde se evaluaron las cuatro plantas centrales.
Análisis estadístico-genético. La comparación de medias se realizó con la prueba de diferencia mínima significativa (DMS), con un nivel de significancia de p ≤0,05. Se realizaron los análisis de varianza por ambiente para la obtención de los cuadrados medios residuales, a objeto de realizar el análisis de homogeneidad, verificada tal condición, se realizó el análisis de varianza combinado para cada carácter, empleando el software Genes (Cruz, 2020). El análisis de la habilidad combinatoria se realizó según el Método 2 modelo 1, de Griffing (1956), teniendo en cuenta las combinaciones p(p+1)/2, correspondientes a los progenitores y sus cruzamientos directos (Ortiz Grisales et al. 2020). Para las pruebas de significancia y ajuste de medias, se consideraron los genotipos y los ambientes con efectos fijos en el análisis de varianza combinado. El análisis genético se llevó a cabo con la ayuda del software Genes (Cruz, 2020).
RESULTADOS Y DISCUSIÓN
Análisis de los valores promedios. En la tabla 1, se presenta el comportamiento promedio de los seis progenitores y quince híbridos F1. En el carácter número de vainas por plantas (NVP), se presentaron variaciones en el material genético evaluado. Los valores promedios de los progenitores oscilaron entre 80,66 y 130,81 vainas por planta. Los progenitores 5 (G16806) y 6 (G51158) presentaron los mayores promedios. La mayoría de los cruzamientos superaron el promedio de los mejores progenitores, destacándose los híbridos (1×2) y (4×5), por la mayor cantidad de vainas por planta, lo que indicó la presencia de efectos heteróticos en los cruzamientos evaluados, en concordancia con lo expuesto por Whankate et al. (2021), quienes obtuvieron en habichuela promedios, entre las 59,40 y 162,40 vainas.
Tabla1. Comparación de medias para número de vainas por planta (NVP), peso promedio de vainas por planta (PPV), longitud de la vaina (LDV) y producción por planta (PPP), en progenitores y cruzamientos de habichuela (Phaseolus vulgaris L), evaluados en dos ambientes. Centro Experimental de la Universidad Nacional de Colombia -sede Palmira (CEUNP) 2019 - 2020.
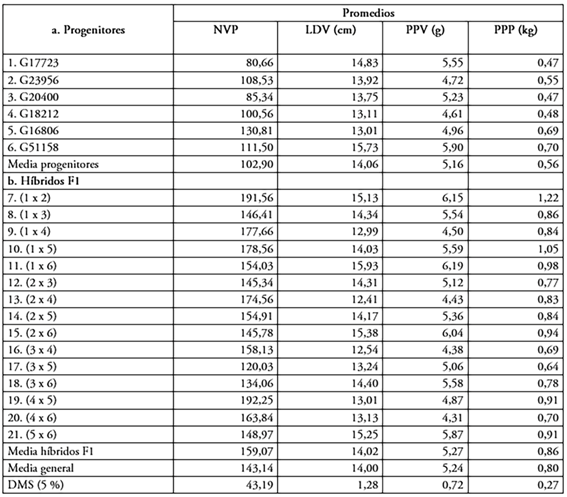
Los progenitores 5 (G16806) y 6 (G51158) presentaron la mayor PPP (Tabla 1). La mayoría de los híbridos se destacaron por su alto promedio en producción por planta en comparación con los mejores progenitores, indicando la presencia de heterosis. Lo anterior, indicó que los progenitores que estuvieron involucrados en la formación de estos híbridos podrían ser empleados en futuros programas de mejoramiento, para el carácter considerado.
Para el carácter peso promedio de vainas por planta (PPV), los progenitores con altos promedios fueron 1(G17723) y 6(G51158). Los cruzamientos que superaron en promedio al mejor progenitor fueron 1×2, 1×6 y 2×6, corroborando la existencia de heterosis en los genotipos evaluados, resultados que difieren en magnitud, de acuerdo con los genotipos empleados por Alves Francelino et al. (2011), quienes obtuvieron valores de 2,27 a 3,81 gramos por vaina y a los de Cordeiro de Araújo et al. (2012), quienes reportaron pesos promedio de 2,51 gramos por vaina. El cultivar 1×2 fue el que obtuvo un mayor peso promedio de vaina por planta, debido a que es un genotipo que se caracteriza por ser altamente productivo y presentar una mayor longitud. Estos resultados brindan una ventaja al agricultor, en donde el incremento en peso del fruto de habichuela cosechado aporta una mayor rentabilidad al cultivo, lo que incrementa los ingresos obtenidos.
Para el carácter longitud de la vaina (LDV), los promedios de mayor valor se encontraron en los progenitores 1 (G17723) y 6 (G51158). Ningún híbrido superó al mejor progenitor, aunque los híbridos 1×2, 1×6, 2×6 y 5×6, con promedios entre 15,93 y 15,13 cm, presentaron diferencias significativas, lo que indica que, probablemente, el progenitor 6 (G51158) está siendo heterótico, con tres progenitores diferentes. Alves Francelino et al. (2011) reportaron valores de 9,31 a 18,79 cm por vaina; Sánchez del Castillo et al. (2017), al evaluar genotipos de crecimiento indeterminado encontraron valores entre 9,3 a 14,5 cm de longitud, en las vainas de habichuela; Vidyacar et al. (2017) obtuvieron valores entre 8,0 a 14,8 cm por vaina; Gomes et al. (2019) encontraron valores de 10,60 a 19,57 cm y Pereira Da Silva et al. (2020) reportaron valores entre los 9,47 a 11,38 cm por vaina. La longitud de las vainas es un carácter importante en la aceptación comercial de los cultivares de habichuela arbustiva y se considera que los valores deben ser superiores a 13 cm (Pandey et al. 2011), por lo que, con base en los resultados obtenidos, es posible seleccionar cultivares que cuenten con este atributo.
Análisis de Varianza Combinado. Los cuadrados medios del ANDEVA combinado presentaron diferencias altamente significativas (P<0,01), para Ambientes (A), en todas las variables evaluadas (Tabla 2). Los genotipos (G) mostraron diferencias altamente significativas (P <0,01), para todos los caracteres, lo que sugiere la presencia de una amplia variabilidad genética, entre los genotipos estudiados. Al descomponer la fuente de variación genotipos (G), se encontró que tanto la HCG como la HCE, presentaron diferencias significativas (P<0,01), para NVP, LDV, PPV y PPP (Tabla 2). Estas diferencias indicaron que en la expresion de dichos caracteres intervinieron de manera significativa, tanto la HCG (acción génica aditiva) como la HCE (acción génica no aditiva), lo que genera la posibilidad de obtener nuevas líneas o híbridos (Da Silva et al. 2004). Resultados similares fueron reportados en habichuela, por Arunga et al. (2010), para las características número de vainas por planta, peso promedio de vainas y longitud de las vainas y Cunha Dias et al. (2016), para número de vainas por planta y longitud de las vainas en Caupi, indicando la posibilidad de ganancias con la selección.
Tabla 2 Suma de cuadrados expresado en porcentaje de la suma de cuadrado total, significancia de los cuadrados medios del análisis de varianza combinado y componentes de varianza de la habilidad combinatoria general (HCG) y la habilidad combinatoria específica (HCE).
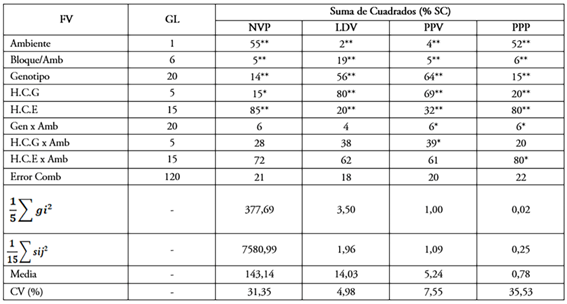
*,** significativo para el test F (P < 0,05) y (P < 0,01), respectivamente; NVP= número de vainas por planta; LDV= longitud de la vaina; PPV= peso promedio de vainas por planta y PPP= producción por planta
La interacción G x A fue significativa (P <0,05), para PPV y PPP, lo que explica el comportamiento diferencial de los genotipos al someterlos a ambientes. Algunos estudios, como los de Getachew et al. (2015), Mounir et al. (2015), Fayaz Ahmad et al. (2016) y Taha (2019), encontraron interacciones significativas entre los genotipos de Phaseolus vulgaris, en uno o más caracteres de interés. La interacción HCG x A fue significativa (P<0,05), solo para el carácter PPV, indicando que los efectos de HCG cambian según el ambiente, lo que sugiere la necesidad de seleccionar progenitores, con efectos de HCG significativos, a través de los ambientes. En cuanto a la interacción HCE x A fue significativa (P<0,05), solo para el carácter PPP, lo que indica que las combinaciones híbridas no mantienen sus efectos de HCE, a través de los ambientes.
En la participación de las fuentes de variación en la suma de cuadrados, para el carácter NVP (Tabla 2), se observó que la variación fenotípica fue explicada por el ambiente (A), con 55 %, seguido por el genotipo (G), con 14 % y en menor proporción la interacción G x A, con 6 %; para el carácter PPP, la variación fenotípica fue explicada por el ambiente (A), con 52 %, seguido por el genotipo (G), con 15 % y, en menor proporción, la interacción G x A, con 6 %. La fuente de variación ambiente (A) muestra que gran parte de la variación fenotípica fue debido a las variaciones ambientales que se presentaron. La mayor magnitud de la suma de cuadrados de los genotipos (G), en comparación con la interacción G x A, muestra la respuesta promedio diferente de los genotipos, por la divergencia genética y como su comportamiento se vio afectado por la oferta ambiental. Para los caracteres LDV y PPV, la mayor contribución a la suma de cuadrados fue dada por el genotipo (G), con 56 % y 64, respectivamente, seguido de la interacción GxA, con 4 y 6 %, respectivamente y, en menor proporción, el ambiente (A), con 2 y 4 %, respectivamente; estos resultados, se deben a la divergencia genética presente en los genotipos empleados, además de la oferta ambiental que favorece la expresión. La manifestación de estas diferencias supone la presencia de variabilidad genética entre los genotipos y se espera que sea de fundamental importancia para trabajos de esta naturaleza, como enfatizan Krause et al. (2012), Da Rocha et al. (2014), Machado (2014) y Pereira Leite et al. (2019).
La suma de cuadrados de los efectos genéticos de HCG fueron de mayor expresión en LDV (80 %) y PPV (69 %) y los efectos de HCE, lo fueron en NVP (85 %) y PPP (80 %). Estos resultados concuerdan con los reportados por Iqbal et al. (2011), Rodrigues et al. (2018) y Owusu et al. (2020), quienes encontraron que la acción genética no aditiva constituye el componente genético más importante del carácter número de vainas por planta (NVP), en Caupí y habichuela, pero difieren de los reportados por Arunga et al. (2010), Da Rocha et al. (2014) y Cunha Dias et al. (2016), quienes afirman que en el NVP los efectos de HCG son el componente más importante en Caupí y Phaseolus.
En el análisis de los componentes de varianza (Tabla 2), se confirmó que en el control genético de los caracteres NVP y PPP hubo predominio de los efectos no aditivos (
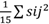 =7580,99 y 0,25, respectivamente), sobre los aditivos (
=7580,99 y 0,25, respectivamente), sobre los aditivos (
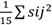 =377,69 y 0,02, respectivamente), por lo que se recomienda mejorar estos caracteres por hibridación. Por el contrario, para el caracter LDV, el componente de varianza, debido a la HCG
=377,69 y 0,02, respectivamente), por lo que se recomienda mejorar estos caracteres por hibridación. Por el contrario, para el caracter LDV, el componente de varianza, debido a la HCG
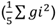 , contribuyó más a la variación genética, con un valor de 3,50, con relación al componente de varianza, debido a la HCE
, contribuyó más a la variación genética, con un valor de 3,50, con relación al componente de varianza, debido a la HCE
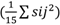 , con 1,96; por lo tanto, se recomienda mejorar este carácter por selección, para así seguir acumulando genes deseables. Para PPV, los componentes de varianza, debidos a la HCG
, con 1,96; por lo tanto, se recomienda mejorar este carácter por selección, para así seguir acumulando genes deseables. Para PPV, los componentes de varianza, debidos a la HCG
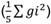 , y HCE
, y HCE
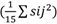 fueron igualmente importantes, ya que los componentes de varianza son prácticamente equivalentes con la ocurrencia de valores 1,00 y 1,09, respectivamente. Resultados similares obtuvieron Das et al. (2014) y Nayak et al. (2018), para este carácter; esto muestra que los progenitores se pueden utilizar en programas de mejoramiento, ya sea dentro de poblaciones, como entre poblaciones.
fueron igualmente importantes, ya que los componentes de varianza son prácticamente equivalentes con la ocurrencia de valores 1,00 y 1,09, respectivamente. Resultados similares obtuvieron Das et al. (2014) y Nayak et al. (2018), para este carácter; esto muestra que los progenitores se pueden utilizar en programas de mejoramiento, ya sea dentro de poblaciones, como entre poblaciones.
Efectos de la habilidad combinatoria general (ĝi). Las estimaciones de los efectos de habilidad combinatoria general se presentan en la tabla 3. Estos efectos determinan el desempeño promedio de cada progenitor en las combinaciones híbridas en las que participa y es una estimación de su valor genético (Vallejo & Estrada, 2013). Así, los progenitores con estimaciones de ĝi altas y positivas son los que más contribuyen al aumento de la expresión de la característica heredada genéticamente, mientras que aquellos con valores ĝi altos y negativos, contribuyen a la reducción de su manifestación. El conocimiento de la habilidad combinatoria, junto con el desempeño per se, es de gran utilidad para la mejora de cualquier carácter (Priya et al. 2018). Por lo tanto, los progenitores con los valores más altos de ĝi y buen desempeño per se, se podrían incluir en programas de mejoramiento genético, para cumplir con los objetivos de selección de nuevos cultivares y así lograr una mejora sustancial en el rendimiento y los componentes del rendimiento. Para el carácter número de vainas por planta (NVP), los progenitores 4 (G18212) y 5 (G16806) presentaron valores positivos y altos. El progenitor 5 (G16806) presentó el mayor valor promedio número de vainas por planta (130,81 vainas por planta), lo que indica que este progenitor es el más prometedor, para mejorar este carácter en habichuela.
Tabla 3 Estimativas de los efectos de habilidad combinatoria general (ĝi), en seis progenitores y efectos de habilidad combinatoria específica (Ŝij), en 15 híbridos de habichuela (Phaseolus vulgaris L). Centro Experimental de la Universidad Nacional de Colombia -sede Palmira (CEUNP). 2019 - 2020.
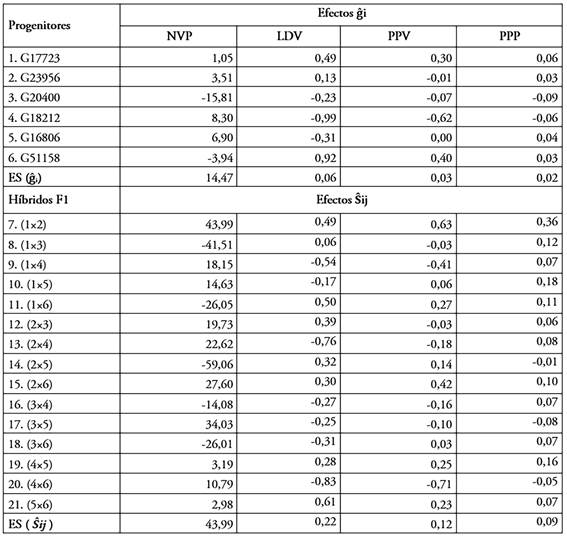
NVP= número de vainas por planta; LDV= longitud de la vaina; PPV= peso promedio de vainas por planta y PPP= producción por planta; ES= Error estándar.
Los progenitores 5 (G16806) y 6 (G51158) mostraron efectos positivos para el carácter producción por planta (PPP) e, igualmente, exhibieron los mayores valores en producción per se. En ese sentido, la contribución de estos progenitores es significativamente superior al resto de progenitores. Dichos progenitores realizan una contribución genética positiva para aumentar el carácter producción por planta. En el caso de la habichuela, que es una planta autógama, esta importancia es aún mayor porque lo único que el mejorador puede aprovechar al final del proceso de mejora es la acción genética aditiva.
Para el carácter peso promedio de la vaina (PPV), los progenitores 1 (G17723) y 6 (G51158), tuvieron estimaciones altas y positivas de ĝi, además de promedios per se altos, lo que indica que estos progenitores podrían contribuir en el incremento del peso promedio de las vainas, ya que estas líneas tienen el potencial para transferir genes favorables a sus descendientes.
Para la longitud de la vaina (LDV), los progenitores 1 (G17723) y 6 (G51158) presentaron las estimaciones más altas y positivas de ĝi y un mayor valor promedio per se, aunque el progenitor 6 (G51158), se destacó por su alto promedio. Estas altas y positivas estimaciones de ĝi y promedio per se muestran una alta varianza aditiva, lo que convierte a estos progenitores en fuertes candidatos para producir buenas progenies. Estos resultados son similares a los encontrados por Gomes et al. (2019), quienes afirmaron que los efectos ĝi positivos son indicadores útiles para el carácter LDV y, posteriormente, sugirieron que estos podrían ser explotados para la obtención de líneas superiores de habichuela.
De forma general, se pudo observar que para los caracteres PPV y LDV, los progenitores 1 (G17723) y 6 (G51158), manifestaron efectos positivos y altos de HCG (acción de genes aditivos), mientras para el carácter NVP, fueron los progenitores 4 (G18212) y 5 (G16806) y para el carácter PPP, lo fueron los progenitores 5 (G16806) y 6 (G51158), por tanto, se pueden utilizar como progenitores en un programa de hibridación, para obtener recombinantes superiores en la progenie.
Efectos de la habilidad combinatoria específica (Ŝij). En la tabla 3, se muestran las estimaciones de los efectos de HCE (Ŝij). Los efectos de Ŝij permiten identificar progenitores con buena complementariedad, teniendo en cuenta la frecuencia de genes con efectos no aditivos (Benin et al. 2009). Cruz et al. (2012) afirman que los híbridos más prometedores serán aquellos que muestren efectos elevados de HCE y que provienen del cruzamiento de progenitores divergentes, donde al menos uno de ellos tiene una HCG alta. De este modo, los híbridos 1×2 y 3×5 tuvieron los mayores efectos positivos de Ŝij y alto rendimiento per se, para el carácter número de vainas por planta (NVP), lo que confirma aún más la superioridad de la acción génica no aditiva en la herencia de este carácter. Los altos efectos de HCE, derivados de progenitores con HCG bajo × bajo y bajo × alto, pueden ser debidos a la complementación de loci, con valores altos y bajos de reducida combinación (Raut et al. 2017). De manera similar, la participación de dos combinadores pobres y un combinador pobre por uno alto también produjo cruzamientos potenciales, como se informó por Ayo-Vaughan et al. (2013) y Owusu et al. (2018), en Caupí, para el carácter número de vainas por planta.
Para el carácter produccion por planta (PPP), las estimaciones positivas de Ŝij y rendimientos per se más altos pertenecen a los híbridos 1×2; 1×5; 1×6; 2×6; 4×5 y 5×6. Estos mismos híbridos mostraron, más allá de un valor de Ŝij alto, al menos, un progenitor con un valor de HCG alto, lo cual, es deseable. Estos resultados sugieren el importante papel de acción génica no aditiva en la herencia del carácter producción por planta. La HCE es principalmente una función de las varianzas de dominancia, que ayuda a identificar las combinaciones híbridas superiores para la explotación comercial de la heterosis. Resultados similares fueron reportados por Owusu et al. (2018), Nayak et al. (2018) y Priya et al. (2018).
Los resultados de la HCE, para el peso promedio de las vainas (PPV), mostraron que los híbridos 1×2, 1×6 y 2×6, registraron los valores más altos de Ŝij y rendimiento per se, en el que, al menos, uno o ambos progenitores tuvieron un valor positivo y alto de HCG, lo que indica que estas combinaciones híbridas tienen un alto potencial para avanzar a la homocigosidad, con una posterior selección de líneas fijas para este carácter. Estos resultados coinciden con los presentados por Rodrigues et al. (2016) y Nayak et al. (2018), quienes mostraron altos rendimientos en el carácter peso promedio de la vaina, a partir de progenitores con altas y bajas estimaciones de HCG y la posterior obtención de combinaciones híbridas sobresalientes.
En cuanto al carácter de longitud de vaina (LDV), los híbridos 1×2, 1×6 y 5×6 presentaron las estimaciones más altas de Ŝij y rendimientos per se. En estos cruzamientos intervienen, tanto líneas con alta HCG como con valores bajos y negativos. En este sentido, los híbridos de mayor longitud fueron el resultado de cruzar dos líneas de altos efectos positivos de HCG (progenitores 1 y 6) y, al menos, una línea de alta HCG; combinaciones híbridas similares fueron obtenidas por Rodrigues et al. (2016), Owusu et al. (2018) y Priya et al. (2018). Por lo tanto, se establece que las combinaciones híbridas con valores altos de HCE se pueden considerar en un programa de mejoramiento genético por hibridación.
El análisis de la HCE mostró que el híbrido 1×2 fue el que presentó los valores más altos y positivos para todas las variables estudiadas. La combinación híbrida entre los progenitores 1 (G17723) y 2 (G23956) puede ser explotada en el futuro en un programa de mejoramiento, debido a su potencial para generar líneas superiores para producción y sus componentes.

