











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Sequence learning or learning of a stimulus series has been experimentally studied for several decades. Its purpose has been to analyze how humans manage to learn, remember, and anticipate sequences of events (Kausler, 1966; Lindsay, 2019; Nissen & Bullemer, 1987). One of the procedures used to evaluate sequence learning consists in the presentation of a sequence of events at a controlled rate after which the participant must reproduce the sequence in the same order in which it was presented. These types of procedures are recognized as reproduction or immediate serial recall (ISR) which it can occur in different response modalities, either as vocal or written reproduction or, in the case of position sequences, by pointing out the place where events appeared (Botvinick et al., 2009; Logan, 2021).
Since it has been linked to the study of memory, sequence learning has been considered by representational-type mediational models (Clegg et al., 1998; Healey et al., 2019; Majerus & Oberauer, 2020; Hurlstone & Hitch, 2015). For example, Nairne (2015) has argued that sequence learning is a prototypical task for studying short-term memory. In this case, a participant is presented with a list of 3 to 9 items (numbers, letters, syllables, words, or pictures) and, after a very short time, he or she is asked to say the items out loud in the order in which they appeared. Correct recall of the series has been considered as a type of episodic memorization since information must be associated with the moment and relative position in which it occurred.
In general, two hypotheses have been proposed to explain what happens in a sequence learning situation: associative chaining or event-position association. In the case of associative chaining, it is stated that the participant establishes associations among events in such a way that sequence recall implies that an event serves as a signal for a subsequent event (DuBrow & Davachi, 2013; Ebenholtz, 1963; Lewandowsky & Murdock, 1989; Young, 1962; Young et al., 1963). Initially, each element of the sequence was independent of the others and, therefore, when an error occurred, the participant would no longer be able to recall the sequence. Currently, chaining based only on associations among events is little accepted given the experimental evidence of the role played by the relative position of events and other contextual cues (Lindsay, 2019; Lindsay & Logan, 2021; Tamayo Tamayo, 2019).
On the other hand, the positional hypothesis assumes that the association occurs between an event and the position it occupies in the sequence (Brown et al., 2000; Friedman, 1993; Henson, 1998; Hurlstone et al., 2014; Kao et al., 2020). Current evidence indicates that sequence recall is more effective if events are perceived as being "clustered" (Hintzman, 2016; Lindsay, 2019). Therefore, one of the variables that is considered relevant corresponds to the relationships of contiguity. According to Solway et al. (2012), experimental evidence tends to show that temporal contiguity is more recurrent and relevant than spatial contiguity (or positional contiguity, in terms of the authors).
When a sequence is learned, the first events are the ones that are most easily remembered. It has been called the primacy effect (Tan & Ward, 2000). This effect is identified as a substantial decrease of errors in the first events of the sequence and as an increase of errors in subsequent events (Logan, 2021; Solvay et al., 2012). In general, errors occur more frequently in the central elements of the sequence. For this reason, Leite et al. (2018) carried out four experiments to explore the experimental conditions that promote learning of central elements of a sequence. In a first experiment, they presented 36 lists of 9 consonants each. Every three lists, elements of central positions were repeated with the purpose of giving them salience. Participants were required to orally reproduce each list after it was presented. A primacy effect was found, with low performances in memorization of central and final positions. In the second experiment, the consonant that appeared in position 4 was enclosed in a circle with the purpose of marking the beginning of the block of letters that were repeated every three lists. Results showed an increase in performance of memorizing the element in that position, but the signal was not effective to correctly reproduced the subsequent letters. In the third experiment, all the letters in the central block were circled. As a result, better performances were achieved in memorizing intermediate positions. Finally, in experiment 4, consonants in the central positions were circled but, in each list, they used different letters. The facilitating effect of the signal was lost. Research by Leite et al. (2018) showed that the implementation of signaling strategies of central positions of a sequence favored its recall. It is a relevant finding since errors mostly occur in central positions. Another possibility, however, could be to present the intermediate positions in spatial contiguity and see if this causes a decrease in the number of errors.
In order to separate spatio-temporal contiguity from exclusively temporal contiguity, Tamayo Tamayo (2019) analyzed sequence learning of events that varied in their ordinal position of occurrence. Sequences showed nine asterisks that were presented one by one (with an interval of 1.5s between occurrences) in nine spaces on a computer screen. That is, on a computer screen nine empty spaces appeared in a row. An asterisk appeared randomly in one position at a time (instead of appearing in a sequential order in any direction). After the sequence was presented, the participant had to reproduce it as it had occurred. Once the participant learned the sequence of positions, a sequence of colors and/or numbers was superimposed on the previously learned sequence. Findings indicated that learning the sequence of positions markedly facilitated learning the sequences of numbers and colors. This author concluded that the role of the position of events in a sequence is relevant, especially when analyzing sequences whose components allow the establishment of spatio-temporal relationships. When the latter is the case, it is feasible to analyze the role of spatial and/or temporal contiguity in sequence learning.
The importance of contiguity has been evidenced even in non-human species. McClearn and Harlow (1954) showed how stimulus-response contiguity affects discriminative learning in monkeys. Specifically, using tasks to assess sequence learning such as immediate serial recall tasks, Botvinick et al. (2009) identified human-like performance patterns in macaques. Precisely, they observed that recall accuracy decreased with increasing sequence size (primacy effect). Inoue and Matsuzawa (2009) evaluated differences in performance between young and adult chimpanzees on tasks involving sequence following. After mastering sequences of up to nine numbers, in experiments 3 and 4, the authors evaluated whether the adjacency or non-adjacency of numbers on the screen affected subjects' performance. Specifically, in experiment 4, they used a masking task in which, after the subject pressed the first numeral in the sequence, the other numerals disappeared, and a white square was presented instead. Subjects had to touch the square in which the next number in the sequence had been presented. In both experiments, performance was affected starting from the second non-adjacently presented numeral. Similar results were found by Beran et al. (2004) using Arabic numeral sequences or color sequences. Scarf et al. (2011) found the same effect using 5-item sequences. Zhang et al. (2022) made a comparative study between humans (adults and children) and monkeys (macaques) using an immediate serial recall task of 3, 4, 5 or 6 positions. As the length of the sequence increased, recall accuracy decreased in all cases, being more accentuated in the case of the monkeys in the sequences from 3 positions forward. The main difference was that the recency effect did not occur in monkeys but in humans. These authors suggested that, in the human case, fragments or segments of information (chunks) were possibly established. These segments were formed from the establishment of spatial relationships between consecutive events favoring the participation of working memory. These relationships were stablished by language in the human case. This would explain why monkeys did not improve their performance despite extensive training. The use of sequence segmentation or fragmentation strategies has also been proposed by Botvinick et al. (2009) and Miller (1956).
It is plausible to think that this type of strategy can be favored if the positions are presented in spatial contiguity. For example, Lindsay and Logan (2021) have suggested that the association between events in a sequence is favored if some property of them allows their grouping. Relationships can occur among events, among events and the place where they occur, or with adjacent cues (Hurlstone et al., 2014; Kao et al, 2020, Lindsey, 2019).
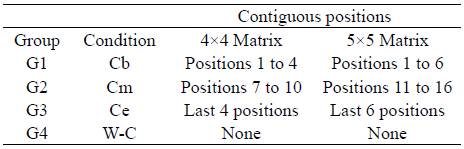
The purpose of this study was to analyze the role of spatial contiguity and the moment of its occurrence within the sequence as a facilitating condition for its learning. To give relevance to the ordinal position of the events presented randomly in a space, the procedure used by Tamayo Tamayo (2019) was modified to consider several rows of positions. Thus, 2 matrices were designed, one with 4 rows x 4 columns (16 positions) and the other with 5 rows x 5 columns (25 positions). The first 4 (in the 4x4 matrix) or 6 positions (in the 5x5 matrix) were presented contiguously for the first experimental group (contiguity at the beginning of the sequence); for a second group, contiguity occurred in the 4 or 6 intermediate positions of the sequence; finally, for a third group, contiguity was present in the final 4 or 6 positions of the sequence. In each group, the other positions were presented in temporal but not spatial contiguity. Due to the primacy effect, it was expected that the condition in which spatial contiguity occurred at the beginning of the sequence would favor sequence learning to a greater degree.
Method
Type of study
The study was experimental. A univariate design of more than two randomized groups according to Castro (1990) was used; or a completely randomized one-factor design according to Myers (1979).
Participants
28 students participated. They were studying electrical mechanical engineering at the University of Guadalajara. There were 24 men and 4 women with an age range of 18 to 26 years (x̅ = 21.5; SD = 1.57). All of them were enrolled in the electromagnetism subject and were invited through their class teacher. Interested students communicated with the experimenters via email. After describing the objective of the study, the conditions in which it would take place and dispelling any doubts regarding the risks of their collaboration, those who agreed to participate expressed it in writing an email. Then, videoconferences were scheduled according to their time availability. In this sense, the only inclusion criterion was that they were enrolled in the subject and there were no explicit exclusion criteria by the researchers. Participants were assigned to each group based on a schedule that specified the date and time of participation. It was generated from their schedule availability. In this way, the first in the list was assigned to G1, the second to G2, the third to G3, the fourth to G4, the 5th to G1 and so on until completing 7 participants per group.
Some students refused to use the TeamViewer tool (see procedure) and, therefore, decided not to participate. For their participation they received extra points for the final grade in the subject of electromagnetism.
Materials
A task designed in a Java environment was used. It consisted of an application that allowed configuring the differential presentation of several matrices with a specific number of cells organized in rows and columns. Each cell corresponded to a position. Each cell measured 1.5 x 1.5 cm. The distance between cells was 0.5 cm.
Inside each matrix, a sequence was configured that consisted of marking one cell at a time with red color until all cells were completed (see procedure). The sequence of positions could be defined randomly or according to the criteria stablished by the experimenter. Given the manipulated conditions, the latter was the case.
The application returned four data files in txt and csv format that included participant's data, the experimental conditions that were set up, selected cells in each trial (each time the participant had the opportunity to reproduce the sequences), time between answers, and the position of the cursor on the computer screen during the presentation of the sequence. For data collection, the TeamViewer tool was used. Excel was used for data analysis.
Design
Participants were assigned to one out of four experimental groups distinguished by where in the sequence a group of positions occurred contiguously. Each participant was presented with two sequences, one in a 4x4 matrix (16 positions) and one in a 5x5 matrix (25 positions). Matrices were selected from a pilot study in which the maximum number of positions that individuals could learn under this procedure was identified. Participants only passed to the 5x5 matrix when they were able to correctly reproduce the position sequence of the 4x4 matrix (see procedure). The number of trials or attempts required by each participant to reproduce the sequence correctly was considered as a dependent variable.
Table 1 presents the design with the conditions to which each group was exposed.
Procedure
Given the confinement conditions derived from the SARS-COV2 virus, the application of the tool was carried out through TeamViewer so that each participant connected remotely to the experimenter's computer. Moreover, participants and the experimenter connected through a videoconference to welcome the participant, answer questions regarding directions, and control what the participant did during the experimental session. Participants were asked to keep the camera on during their participation. However, the researcher's camera was turned off and the microphone was muted to simulate the privacy of the participant's interaction with the experimental task.
To start, a TeamViewer session was initiated, and remote connection was established from the participant's computer to the experimenter's one. Previously, the experimenter had loaded the application for the participant with their data according to the corresponding group condition.
Once the connection was established, the participant would see a screen with the following instruction:
Welcome. We appreciate your participation. You will solve a memory task. You will initially be presented with a 4x4 grid. A sequence will appear in it. After that, your task will be to try to reproduce the sequence as it was presented to you. To do this, click with the mouse on each square of the grid trying to follow the same sequence that was presented to you. You will be told if you are right or wrong. If you make a mistake, don't worry. You can try again several times. If you succeed, then a new 5x5 grid will appear with a new sequence that you must also try to memorize. If at any point you get bored or tired, or think you can no longer remember the sequence, that's fine. The activity will end when you want. During the task you will have the option to quit at any time; just let the person in charge know. However, we invite you to give your best effort.
If you have doubts, you can ask them at this time; otherwise, click start.
After reading instructions and clarifying their doubts with the experimenter, the participant pressed the "start" button. At this point, the experimenter turned off his camera and observed participant's performance both on his computer screen and through the videoconference. A 4x4 grid appeared on the screen. Red color was used to mark the positions. In this way, the sequence began when one of the 16 squares lit up red for one second; after that it stopped being illuminated. One second later, the next square in the sequence was illuminated for one second, and so on until all 16 squares had been illuminated. Once this had happened, all 16 squares appeared on the screen (filling color of squares was white) and the participant had to try to reproduce the sequence as it had been presented. To do this, she or he had to click with the mouse on the first square that had lit up, then on the second, and so on. Once a square was clicked on, it would light up red and stay that way until the participant completed the entire sequence of positions. Next, she or he had to press the "Send" button. At this time, the application provided feedback on participant's performance, indicating the number of errors she or he had made. The application also asked her or him if she or he wanted to try again or if she or he wanted to abandon the task. If she or he pressed the "Try Again" button, she or he was exposed to the same sequence of positions. All this was repeated until the participant was able to reproduce the sequence correctly or until they pressed the "Abandon" button, in which case the application indicated by means of a message that the session had ended and that they were thanked for their participation. The organization of the computer screen during the presentation of the sequence can be seen in Figure 1.

Once participants managed to reproduce the sequence correctly, they were shown the following instruction indicating that they would now be presented with a new sequence, this time 5x5:
"Next, a 5x5 grid is going to appear on the screen and a different sequence is going to be shown to you. The rest is the same. When you're ready, press start."
When the participant pressed the "Start" button, a 5x5 grid now appeared on the screen. The presentation of the sequence and the way of responding, as well as the feedback, were identical to that of the 4x4 matrix. Once the participant was able to reproduce the sequence correctly, the application indicated it to her or him, telling her or him that the session had ended and thanking her or him for her or his participation. It is important to mention that the participant could leave the session by pressing the "Abandon" button after concluding any attempt to reproduce the sequence. Each presentation of the sequence along with its corresponding recall attempt by the participant was counted as one trial.
The initial (G1), intermediate (G2) or final (G3) positions were presented contiguously in space and time. The other positions were contiguous in time, but not in space. In the case of G4, all positions were presented with temporal but not spatial contiguity. Thus, in the case of the 4x4 matrix, the first 4 positions were contiguous in space for G1; 7 to 10 were contiguous for G2; and 13 to 16 were contiguous for G3. In the case of the 5x5 matrix, the first 6 positions were contiguous in space for G1; 11 to 16 were contiguous for G2; and 20 to 25 were contiguous for G3. In these three groups, the other positions occurred in temporal but not spatial contiguity as in the G4 condition. The sequences ofpositions and the contiguity relationships used are presented in Figure 2.
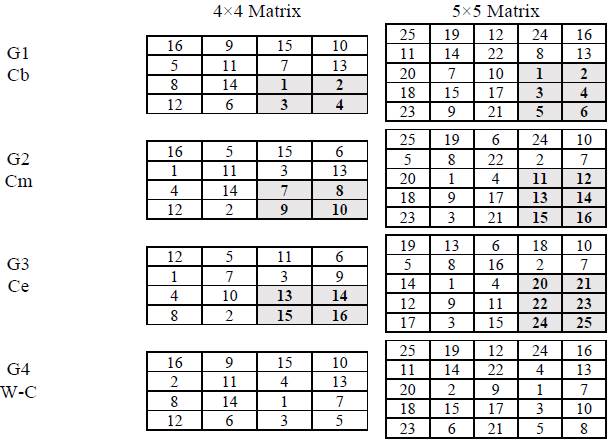
Note: Numbers represent the sequence in which the squares successively lit up red. The bold numbers in gray cells correspond to the positions that occurred in space-time contiguity. Cb: contiguity at the beginning of the sequence; Cm: contiguity in the middle part of the sequence; Ce: contiguity at the end of the sequence; W-C: without spatial contiguity.
Figure 2 Sequence ofpositions used in each of the groups
Ethical aspects
The study was carried out under the guidelines established by the Ethical Principles and the Code of Conduct of the American Psychological Association (2017). The risk level of the study was considered as "low" according to the standards for laboratory research established by the University of Guadalajara. The research protocol was approved by the ethics committee of the University of Guadalajara. During the initial communication via email and during the first part of the videoconference with each participant, they were informed of the aspects considered in an informed consent: conditions of participation, possible risks or inconveniences, right to withdraw without consequences at any time, confidential use of personal data, incentive through extra points in grades and ethical treatment of the data obtained. Before starting the application, each participant was asked if she or he agreed to participate and if she or he allowed her or his performance to be recorded. The sessions were not videotaped and the only record of their performance was the one automatically provided by the task application.
Results
Figures 3 and 4 show the number of trials that each participant needed to reproduce the sequence correctly in each of the groups. Group 1 participants needed an average of 6 trials before successfully reproducing the sequence in the 4x4 matrix (x̅ = 6.4) and 10 trials in the case of the 5x5 matrix (x̅ = 10). The only participant who dropped out of the task was P6 on the second trial of the 5x5 matrix.
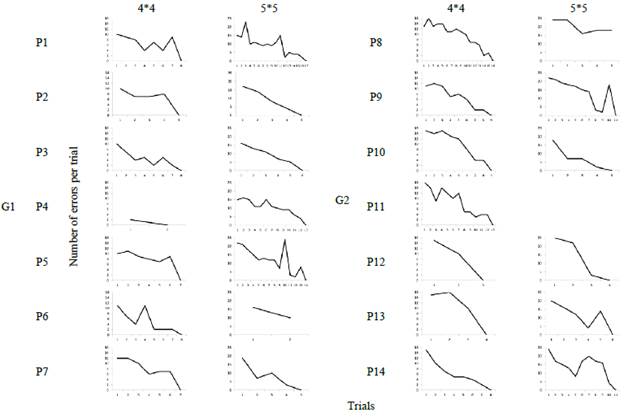
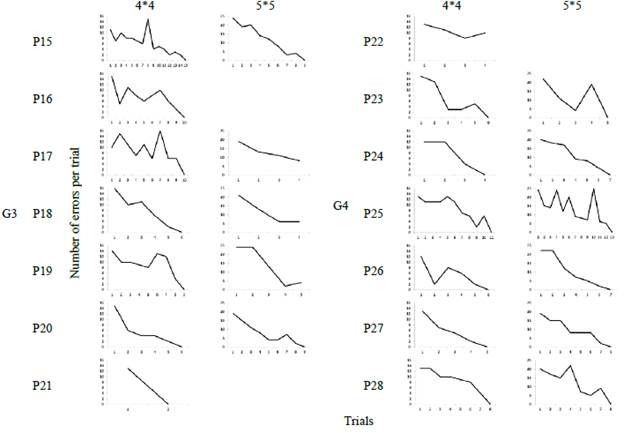
Group 2 participants required around 9 trials to reproduce the sequence correctly (x̅ = 8.8) in the 4x4 matrix. In the 5x5 matrix, participants needed an average of 7 trials (x̅ = 7.4). In this group there were 2 dropouts, P11 before starting the 5x5 matrix and P8 during the 5x5 session.
Group 3 presented most of participant dropouts. P16 abandoned the task before starting the 5x5 matrix, while P17, P18 and P19 did so during the 5x5 session. It should be noted that P21 reproduced the 5x5 sequence in the first attempt. Thus, it does not have a graph indicating the number of trials. In the 4x4 matrix, participants required 8 trials to reproduce the sequence correctly (x̅ = 8.2), while those who completed the 5x5 matrix needed 9 trials (x̅ = 9).
Finally, Group 4 participants required around 6 trials to correctly reproduce the positions of the 4x4 matrix (x = 6.6) and 8 trials in the case of the 5x5 matrix (x = 8). P22 abandoned the task during the 4x4 session.
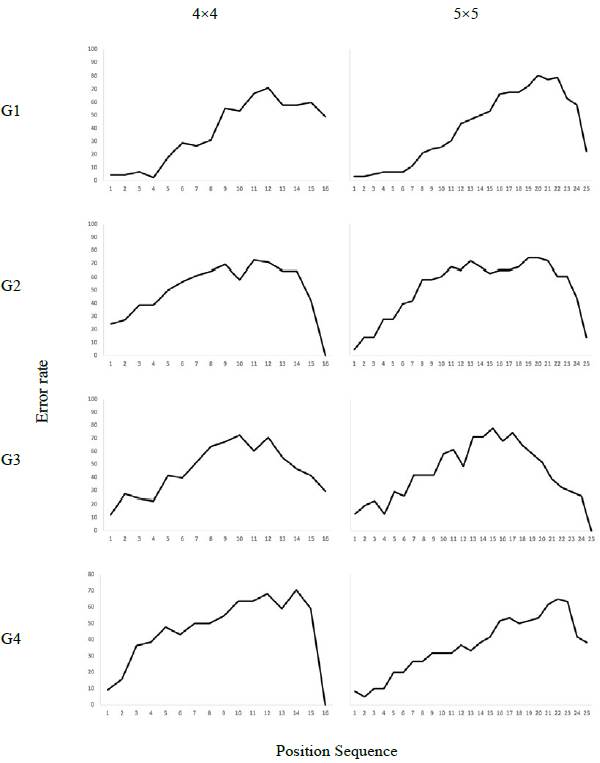
Figure 5 shows error distribution in each position of the sequences in each of the groups. The least number of errors occurred in the first 4 positions of the 4x4 matrix, corresponding to the positions that occurred contiguously in space. Likewise, in the case of the 5x5 matrix, the lowest number of errors occurred in the first 6 positions, which also coincides with the positions that occurred contiguously at the beginning of the sequence. Results of a one-way ANOVA test showed that mean differences in errors during the first 4 positions (of the 4x4 matrix) in G1 was significant (F (3 24) = 5.9, p<.05) compared to groups 2 ((p<.05) 95% CI [-9.02, -9.2]) and 3 ((p=<.05), [-36.1, -0.08]). In the case of the 5x5 matrix, there were significant differences (F (3 20) = 3.5, p<.05) in the average of errors in the first 6 positions of G1 with respect to G2 ((p<.05) CI 95% [- 36.5, -1.6]). In the other groups, no decrease in the percentage of errors in the adjacent positions is observed. The highest number of errors occurred in the middle part of the sequence in all cases with a slight bias to the right in the case of Group 4. This indicates that for Group 4 the highest number of errors occurred in the last positions of the sequence.
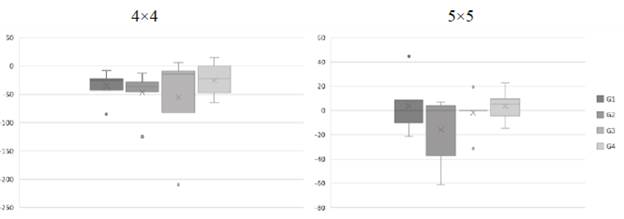
The time it took for each participant to reproduce the sequence on each of the trials was recorded for each of the 16- and 25-position arrays. The difference between recall duration in the last trial of each array (when the sequence was recalled correctly) minus recall duration during the first trial of each of the arrays was calculated. These results are presented in Figure 6. A negative difference indicates that it took participants less time to recall the sequence on the last trial, that is, when they recalled the sequence correctly, compared to the time taken on the first trial; a positive difference indicates the opposite (it took them longer to reproduce the sequence in the last trial than in the first one). As it can be noticed, in the case of the 4x4 matrix, all the differences were negative with a slightly greater dispersion for groups 3 and 4; in the 5x5 matrix, the differences tended to be positive, except for Group 2, which showed a high dispersion. This indicates that it took participants longer to recall the correct sequence on the last trial in the 5x5 matrix compared to the time spent on the first trial of this sequence; in the 4x4 matrix the opposite happened. When comparing groups, it can be noticed that participants in Group 4, although with some variation, tended to spend more time on the last trial (the one in which the sequence was reproduced correctly) than on the first trial, in both sequences.
Discussion
The purpose of the study was to evaluate the effect of spatial contiguity on sequence learning of positions. For this, two sequences of 16 and 25 positions presented in 4x4 and 5x5 matrices, respectively, were used. Within each matrix, the point of the sequence in which the contiguous positions were presented was varied: at the beginning of the sequence, in the middle or at the end. Sequence learning was assessed using a immediate serial recall task.
No differences were observed among groups in terms of the number of trials required to reproduce the sequences correctly, indicating that contiguity did not affect how quickly participants were able to learn them. However, participants made fewer errors in contiguous positions when they were presented at the beginning of the sequence. In the other groups, there was no decrease in the number of errors when the contiguous positions occurred in the middle or at the end of the sequence.
In principle, a possible explanation for this result may be that the fact of presenting the initial positions contiguously in space accentuated the primacy effect that it is usually identified in sequence learning. Solvay et al. (2012) have stated that the primacy effect in sequence learning is identified with the decrease in errors at the beginning of the sequence and its increase as the temporal distance is greater, so the effect is represented as a time gradient. Error distribution curves showed an increase in the number of errors in the intermediate positions and a decrease at the beginning (primacy) and at the end (recency) of the sequences (Farrand et al., 2001; Jones et al., 1995). All error percentage curves obtained in this study show this trend. However, the primacy effect is clearly more accentuated in Group 1 in which the initial positions were presented contiguously in space.
Another possibility is that the contiguous presentation of the positions at the beginning has favored the segmentation in parts or fragments of the sequences as a strategy for their learning (Tamayo Tamayo, 2019). Participants were able to segment the sequence into parts that were learned trial by trial. The repetition of the sequence until its correct reproduction could have favored this progressive learning. The use of fragmentation or segmentation strategies have been proposed by other authors, mainly in humans but also, although to a lesser extent, in non-humans (macaques) (Botvinick et al., 2009; Miller, 1956; Zhang, et al., 2022). Lindsay and Logan (2021) have recently stated that an element of the sequence can be associated with several subsequent components because they are recognized as belonging to the same group of events. The occurrence of contiguity at the beginning of the sequence could have favored the configuration of groups, segments or fragments of positions that shared the same property (being contiguous in space), which caused fewer errors to appear in these positions, although it did not favor learning speed of the complete sequence (that it was learned in a smaller number of trials). However, it is not clear why the same did not occur in the intermediate and final positions that presented contiguity in groups 2 and 3.
Now, why do the position and its contiguity relationships become a relevant variable for learning this type of sequences? The default position is spatially defined (north, south, east, west, up, down, right, left, or nominally as position 1, position 2, etc. which can be combined with the former). When you have a sequence, whose components are presented in different positions in space, the spatial dimension and the relationships that the individual can establish between the events that occur in space ends up being fundamental for learning the sequences. In this sense, Nairne (2015) has argued that the position of an event must necessarily be conceived in a multidimensional model of occurrence (when it occurs, where it occurs, in relation to what other events occur, how these relationships change over time, etc.). Relationships can be established between events, between an event and its place of occurrence, or between these and adjacent or context cues, as suggested by the associative hypotheses presented in the introduction (Hurlstone et al., 2014; Kao et al, 2020, Lindsey, 2019; Lindsey & Logan, 2021). As Zhang et al. (2022) have pointed out, it is possible that this type of relationship in the human case is possible from language, which would explain the differences in performance in this type of task between humans and non-humans. In this study, there was no record or data that could constitute evidence of the participation of language in an eventual process of segmentation of the sequences, so direct evidence would be required in this regard in subsequent investigations.
From findings reported in reproduction tasks with motor responses that implement a visual tracker (Pathman & Ghetti, 2015, 2016), eye movement during the observation of the sequence is related to the precision in the sequence learning. But the observation pattern that is obtained at the moment in which the participant is choosing the event that was presented in a given position may not necessarily reflect the precision but rather different strategies for selecting the possible correct answer. For example, there are very short fixations in trials where there is some automaticity in the choice of response or recurrent fixations on the correct event. In this sense, recording eye tracking could be relevant, differentiating the moments of presentation of the sequence from those of its reproduction.

