










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkInfectio
versão impressa ISSN 0123-9392
Infect. v.11 n.1 Bogotá jan./mar. 2007
REVISIÓN DE TEMA
Amebiasis intestinal
Intestinal amebiasis
JULIO CÉSAR GÓMEZ1, JORGE ALBERTO CORTÉS2,SONIA ISABEL CUERVO1, MYRIAM CONSUELO LÓPEZ3
1 Unidad de Infectología, Facultad de Medicina, Universidad Nacional de Colombia, Bogotá, D.C., Colombia.
2 Departamento de Medicina, Facultad de Medicina, Universidad Nacional de Colombia, Bogotá, D.C., Colombia.
3 Laboratorio de Parasitología, Departamento de Salud Pública y Tropical, Universidad Nacional de Colombia, Bogotá, D.C., Colombia.
Fecha de recepción: 25/04/2006; fecha de aceptación; 02/02/2007
RESUMEN
Entamoeba histolytica es el patógeno intestinal más frecuente en nuestro medio –después de Giardia lamblia–, una de las principales causas de diarrea en menores de cinco años y la cuarta causa de muerte en el mundo debida a infección por protozoarios. Posee mecanismos patogénicos complejos que le permiten invadir la mucosa intestinal y causar colitis amebiana. El examen microscópico es el método más usado para su identificación pero la existencia de dos especies morfológicamente iguales, una patógena ( E. histolytica) y una no patógena ( Entamoeba dispar), ha llevado al desarrollo de otros métodos de diagnóstico. El acceso al agua potable y los servicios sanitarios adecuados, un tratamiento médico oportuno y el desarrollo de una vacuna, son los ejes para disminuir la incidencia y mortalidad de esta entidad.
Palabras clave: amebiasis, infecciones parasitarias, Entamoeba histolytica.
ABSTRACT
Entamoeba histolytica is the most frequent intestinal pathogen seen in our country, after Giardia lamblia, being one of the main causes of diarrhea in children younger than five years of age, and the fourth leading cause of death due to infection for protozoa in the world. It possesses complex pathogenic mechanisms that allow it to invade the intestinal mucosa, causing amoebic colitis. Microscopy is the most used method for its identification, but the existence of two species morphologically identical, the pathogen one ( E. histolytica), and the non pathogen one ( E. dispar), have taken to the development of other methods of diagnosis. The access to drinkable water and appropriate sanitary services, an opportune medical treatment, and the development of a vaccine are the axes to diminish the incidence and mortality of this entity.
Key words: amebiasis, parasitic infections, Entamoeba histolytica.
William Petri Jr. resume los alcances logrados en la investigación sobre este protozoario de la siguiente manera:
"Es un momento emocionante en el estudio de la Entamoeba histolytica. En los últimos dos años la historia natural y el riesgo de la enfermedad en humanos se han redefinido, se han identificado las respuestas inmunes en mucosas asociadas a protección y se han descrito los procesos reguladores del desarrollo de la exquistación. Los avances en la biología celular proveen nuevas luces sobre la invasión a través del epitelio intestinal" (1).
La diarrea dejó de ser la causa más importante de muerte en menores de cinco años debido al advenimiento de las sales de rehidratación oral y el mayor acceso al agua potable. Conocemos muchos de los mecanismos que causan dichas infecciones, avanzamos cada vez más en aras de su control, el papel de Entamoeba histolytica es mejor comprendido y desarrollamos nuevos métodos para su diagnóstico (2). A continuación se revisan los aspectos más importantes de este patógeno, haciendo énfasis en los factores de virulencia y los factores del hospedero que facilitan la infección.
AGENTE ETIOLÓGICO
E. histolytica fue descubierta por Friedrich Lösch en 1873 en Rusia, quien la aisló de un paciente con disentería e, incluso, estableció la relación existente entre la enfermedad y el protozoario en perros infectados experimentalmente con quistes amebianos procedentes de humanos (3). En 1925, Brumpt propuso que existían dos especies distintas, una patógena y una no patógena, aunque morfológicamente iguales (4). En 1961, Diamond logró el primer cultivo axénico (5) y, en 1978, Sargeaunt diferenció la variante patógena de E. histolytica de una no patógena mediante patrones electroforéticos de isoenzimas (6). En 1993, Diamond demostró que realmente eran dos especies diferentes (7) y, en 1997, la OMS recomendó que se diferencien las dos especies para efectos de diagnóstico y tratamiento, y, cuando el diagnóstico se lleve a cabo por microscopía óptica, se reporte como E. histolytica/dispar (8).
Es un protozoario que presenta dos formas en su ciclo vital: quiste y trofozoíto (figura 1). Los quistes son estructuras redondeadas de 10 a 16 µm, con una cubierta gruesa y que presenta en su interior 1 a 4 núcleos. El trofozoíto tiene un diámetro de 20 a 40 µm y es móvil, gracias a su ectoplasma que le permite formar un seudópodo; su núcleo presenta un cariosoma compacto central y cromatina en gránulos uniformes en tamaño y localización (1, 2, 9). En el 2005 se publicó el genoma de E. histolytica (10), con un tamaño de 23.751 kilobases y 9.938 genes. No hay una clara conformación cromosómica y se confirmó la ausencia de mitocondrias; sin embargo, es necesario obtener el genoma de E. dispar para compararlos y hallar el origen de capacidad patógena.
CICLO DE VIDA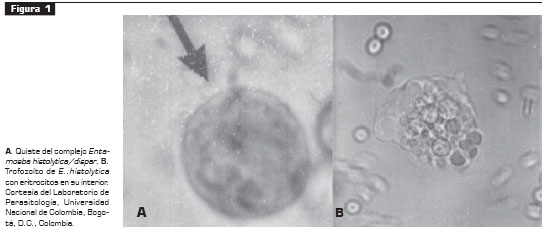
El ciclo de vida se inicia con la ingestión de un quiste infeccioso. Gracias a su dura cubierta no es dañado, aunque si reblandecido, por la acción de los jugos gástrico y pancreático, lo cual permite su exquistación en el intestino grueso, donde puede desarrollar infección invasora o no hacerlo. Al romperse el quiste, el trofozoíto termina su proceso de división y da lugar a cuatro trofozoítos metaquísticos (9). Si el trofozoíto continúa avanzando por el colon, inicia su proceso de enquistación con la formación de un prequiste mononuclear, en el cual se inicia un proceso de división celular que da lugar a un quiste tetranuclear, en el cual termina el proceso de formación de la pared del quiste, el cual es expulsado con la materia fecal (2). Este quiste es muy resistente, sobre todo en climas húmedos, y puede infectar si es ingerido. Este proceso lo apreciamos resumido en la figura 2. El proceso que lleva a la infección intestinal se expondrá más adelante.
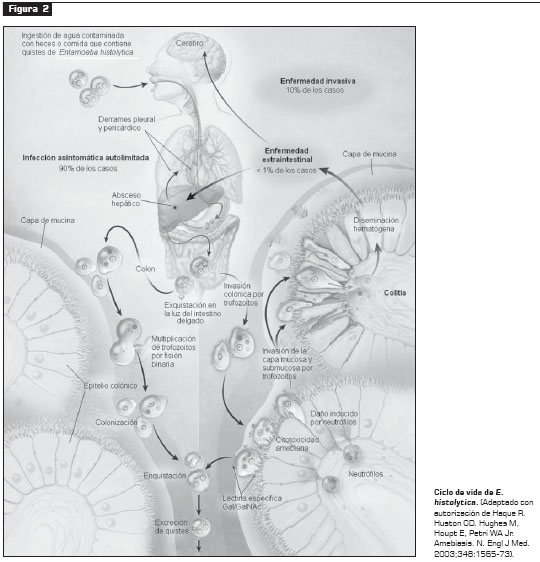
EPIDEMIOLOGÍA
La amebiasis es una de las parasitosis más tempranamente reconocidas y universales. Es la cuarta causa de muerte en el mundo debida a infección por protozoarios después del paludismo, la enfermedad de Chagas y la leishmaniasis, y la tercera causa de morbilidad después del paludismo y la tricomoniasis (11). Centro y Suramérica, África y Asia se consideran zonas endémicas para amebiasis.
Los factores de riesgo en las naciones desarrolladas son: ser inmigrante de un país en vías de desarrollo, haber viajado a los trópicos, tener compromiso inmune, ser residente de una institución para retardados mentales o ser un hombre que tiene sexo con hombres. Vale la pena anotar que, actualmente, este último factor de riesgo puede deberse en parte a un sesgo por la búsqueda de la homosexualidad como factor de riesgo de muchas entidades, lo cual lleva a un enfoque que podría llegar a ser discriminatorio (12). En 1986, Walsh calculó que E. histolytica infecta al 10% de la población mundial (13) y se estima en 50 millones el número de casos anuales, lo que resulta en 40 a 110 mil muertos por año (2, 11,13). Sin embargo, estas cifras son previas a la redescripción de las dos especies por Diamond y se basan en el diagnóstico microscópico o por seroprevalencias, lo que lleva a problemas de estimación (11). En México, la seroprevalencia es de 8,4%, pero los estudios moleculares muestran cifras de infección asintomática por E. histolytica de 13,8% y de 9,6% por E. dispar (14), así que desconocemos la verdadera incidencia y prevalencia de la enfermedad.
La amebiasis puede transmitirse a través del consumo del agua, incluso en naciones desarrolladas, dado que no se usan marcadores parasitológicos para determinar la pureza del agua; por lo tanto, en toda nación con amebiasis endémica pueden presentarse brotes de infección por consumo de agua contaminada (11, 15, 16).
Aunque la amebiasis no es una infección oportunista, no es clara la relación entre inmunosupresión e amebiasis. Se ha observado una incidencia creciente de la amebiasis en pacientes con VIH, lo que ha alertado sobre la disminución en la práctica de sexo seguro y sobre la posibilidad de que el VIH sea un factor de riesgo independiente (17-20). Sin embargo, no se ha demostrado que la infección por VIH aumente la seriedad de la infección amebiana (9), aunque algunos autores lo proponen (17).
En un modelo experimental con ratones se describió que la disminución de los linfocitos T CD4+ disminuye la gravedad de la enfermedad, posiblemente por la disminución de las interleucinas (IL) proinflamatorias 4 y 13, las cuales activan de forma menos eficaz los macrófagos y neutrófilos que el factor de necrosis tumoral alfa (FNTalfa) y el interferón gamma (IFNgamma) (21). Al parecer, los diabéticos se encuentran en mayor riesgo de amebiasis (22) y se ha reportado la aparición de amebiasis en pacientes que reciben quimioterapia, receptores de trasplantes de médula ósea y de órgano sólido (23). En un estudio en pacientes con compromiso inmune, que incluía, neoplasias hematológicas e infección por VIH, la incidencia de infección por E. histolytica/E. dispar fue de 9,91% (24). En el Instituto Nacional de Cancerología (Bogotá, D.C., Colombia) se reportó una prevalencia de amebiasis de 7% en pacientes con neutropenia febril (25), de 24,7%, en pacientes que fueron llevados a trasplante autólogo de médula ósea (26) y de 14,3%, en pacientes con colitis neutropénica (27). En los pacientes con inmunosupresión o aquéllos que residen en zonas tropicales, no hay variables que permitan estratificar el riesgo de presentar infección o de no hacerlo, a pesar de ser los sitios más afectados por esta situación, por lo que es necesario comenzar a controlar los factores de riesgo conocidos (sanidad pública) e investigar otros factores que puedan favorecer la infección.
PATOLOGÍA
Desde Lafleur, los aportes a la descripción patológica son pocos. La amebiasis intestinal puede presentar un espectro de alteraciones anatómicas, de acuerdo con su localización, que puede ir de una colitis ulcerativa a una apendicitis amebiana, pasando por el megacolon tóxico y el ameboma.
El hallazgo anatomopatológico típico son las úlceras localizadas en el colon, predominantemente en el ciego, el sigmoide y el recto. Estas úlceras presentan dos patrones claramente definidos: nodular e irregular. Las úlceras nodulares son redondeadas, de un diámetro entre 1 y 5 mm, con áreas de mucosa ligeramente elevadas y áreas necróticas, deprimidas o hemorrágicas, rodeadas por un borde de tejido edematoso. A menudo estas áreas están llenas de un material mucoso y amarillento, denominadas "úlceras en botón de camisa", en las cuales pueden verse trofozoítos (figura 3). En ocasiones, estas lesiones pueden llegar a cubrir la mayor parte de la mucosa del colon, causando edema y eritema en las áreas mucosas que no se encuentran comprometidas. Las úlceras irregulares o serpiginosas tienen 1 a 5 cm de longitud y se encuentran habitualmente en el ciego y el colon ascendente. Sus márgenes suelen ser elevados y edematosos, y la úlcera se llena de fibrina. Cuando las úlceras son grandes, las áreas de mucosa sin compromiso se encuentran congestivas y edematosas. Ambos cambios morfológicos pueden encontrarse en un mismo paciente, con extensas áreas de denudación superficial (28).
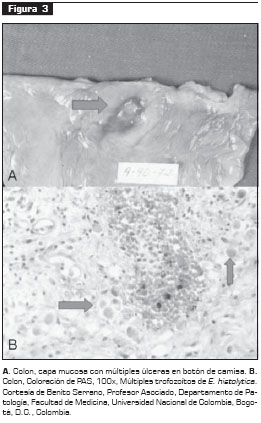
Los hallazgos microscópicos se describen en cinco etapas: lesión inespecífica, depresión mucopénica, lesión invasiva temprana con ulceración superficial, lesión invasiva tardía con ulceración profunda y la úlcera en granulación. Dado que estas etapas son la manifestación de cada una de las fases de la patogénesis, se las describirá junto con ellas.
FASES Y MECANISMOS DE LA PATOGÉNESIS
El proceso de invasión desarrollado por E. histolytica es uno de los más elaborados entre los patógenos intestinales. La capa de mucina que recubre el epitelio del colon es el principal obstáculo para invadirlo, por lo que desarrolló sofisticados mecanismos para penetrarla. En 1970, Prathap y Gilman publicaron una clasificación de las úlceras amebianas de acuerdo con su apariencia microscópica (28). Posteriormente, se vio que esta descripción coincidía adecuadamente con las fases de la patogénesis, con la intervención de diversos mecanismos moleculares en cada fase, tanto del hospedero como del patógeno. Las fases de la patogénesis son tres: colonización, disminución o disrupción de la mucosa y enlace, y lisis de las células epiteliales del colon del hospedero (29). Si bien la infección es un proceso dinámico que involucra factores del hospedero y del agresor, la revisión se centrará predominantemente en el papel del agresor y sus mecanismos moleculares.
Lesión inespecífica. Se observa engrosamiento de la mucosa, hiperplasia glandular y edema del estroma. Durante esta fase ocurre la etapa de colonización y se inicia la fase de disrupción de la mucosa. La adherencia del trofozoíto es difícil debido a las cargas negativas de la capa mucosa, pero, por medio de la lectina galactosa–N-acetil-Dgalactosamina se adhiere a ésta (30). Una vez adherida, secreta una serie de proteínas, entre ellas las proteasas de cisteína, por medio de las cuales causa un efecto paradójico: promueve la producción de mucina y su secreción a la luz intestinal, lo que lleva a hiperplasia glandular (28, 30, 31).
Depresión mucopénica. En esta etapa se aprecian lesiones mucopénicas, pequeñas depresiones asociadas a pérdida de mucosa, con adherencia de los trofozoítos a ellas. El mecanismo propuesto es que las células caliciformes encargadas de la producción de mucina agotan su capacidad de reponer la mucina destruida por el trofozoíto (28). Si bien la lectina permite la adherencia del trofozoíto, ésta puede ser inhibida por la misma mucina, por lo que, al parecer, es necesaria la alteración de las mucinas por un mecanismo a distancia a cargo de proteasas y otras sustancias (28-30). Se conoce la capacidad de E. histolytica de secretar glucosidasa, manosidasa, galactosidasa, fucocidasa, xilosidasa, glucuronidasa y N-acetil-D-galactosaminidasa, entre otras, y, en asociación con las proteasas de cisteína, de abrirse paso a través de la capa mucosa. Así se completa la fase de reducción/disrupción de la mucosa.
Lesión invasiva temprana con invasión superficial. Durante esta fase clave se presentan tres fenómenos: erosión epitelial superficial, focos de microinvasión interglandular e infiltración de la lámina propia (28), completándose así el trío de acciones que implican la patogenia clásica.
Durante la erosión epitelial superficial, los trofozoítos alcanzan la superficie de la luz de los enterocitos e inician un proceso de lisis que depende del contacto (28, 30). In vitro se determinó que este complejo proceso se desarrolla en varios pasos: adhesión, daño dependiente del contacto ( hit and run) a la membrana plasmática, fagocitosis y degradación intracelular de las células ingeridas (28). Estos eventos se relacionan con tres tipos de moléculas: la lectina galactosa–N-acetil-D-galactosamina (Gal-GalNac lectin), los amebaporos y las proteasas de cisteína (32).
La lectina Gal-GalNac se liga a los residuos galactosa–N-acetil-D-galactosamina de las glucoproteínas de la célula blanco. Otras moléculas que participan en la adhesión son una lectina de 220 kd, una adhesina de 112 kd y un lipofosfoglucano. La lectina Gal-GalNac tiene otras funciones, como se describe más adelante. Es uno de los principales factores de virulencia y su bloqueo interfiere en la capacidad de adherirse e invadir (28, 30).
Una vez se ha logrado la adhesión y el contacto con la célula blanco, ocurre una rápida lisis celular (1, 28, 30), lo que lleva a edema, vacuolización de la membrana y lisis de la célula blanco, la cual puede ser linfocitos, neutrófilos o macrófagos. La proteína amebaporo se ha encontrado en lisados de trofozoítos de E. histolytica y se han determinado muchas de sus características químicas y físicas. In vitro se ha apreciado su efecto citolítico contra una gran variedad de líneas celulares humanas (28) y, actualmente, se sabe que se ubica dentro de las vesículas citoplasmáticas; sin embargo, no se ha logrado demostrar que participe en los procesos de lisis celular. No se han logrado obtener indicios de su secreción extracelular y, al parecer, su función es la de lisar las bacterias fagocitadas como mecanismo de defensa. Las proteasas de cisteína son un grupo de proteínas con habilidad de degradar la matriz extracelular y que participan en los procesos de lisis celular.
Una vez es penetrada la capa de enterocitos luminales, se establecen unos focos de microinvasión interglandular que se aprecian como zonas de erosión epitelial superficial con invasión de trofozoítos entre las glándulas del colon. En este punto, los trofozoítos penetran y se trasladan hacia capas más profundas; tienen que lisar los enterocitos circundantes y la matriz extracelular para seguir avanzando, llevando a cabo una lisis celular continua, una invasión a través de la locomoción y la degradación de la matriz extracelular.
No se ha explicado completamente la lisis celular continua de esta fase. Se evidenció que, aparte de la habilidad de lisar por sí misma a las células blanco, por un mecanismo no del todo conocido, E. histolytica induce la caspasa humana 3 que lleva a apoptosis de las células (33) pero, al parecer, es inducido por la lectina Gal-GalNac y por los mecanismos de señalización basados en cinasas transmembrana (34). Además, la fagocitosis de células apoptósicas es más rápida que la de células previamente sanas, lo que demuestra que ésta es una forma importante para que el trofozoíto consiga nutrientes.
Por otra parte, el trofozoíto atrae neutrófilos que infiltran la lámina propia, fenómeno mediado principalmente por la interleucina 8, cuya expresión y secreción por las células epiteliales intestinales son inducidas por los trofozoítos (28, 30); los trofozoítos también bloquean otras rutas quimiotácticas, aparentemente con el fin de atraer sólo polimorfonucleares. ¿Qué papel juegan los neutrófilos? Al ser atraídos por el trofozoíto se exponen a ser lisados por éste y así aumentan la lisis tisular, lo que permite a E. histolytica continuar su expansión por la mucosa y la submucosa. El trofozoíto resiste la acción tóxica del neutrófilo gracias a una ferro-superóxido dismutasa y el peróxido producido se elimina a través de una proteína rica en cisteína, de 29 kd, que suple la catalasa que no contienen los neutrófilos (1, 28, 30).
El citoesqueleto de E. histolytica le permite realizar dos funciones importantes para la patogénesis: la locomoción y la fagocitosis. Esto se logra mediante la polimerización de la actina, la cadena pesada de la miosina II y un conjunto de proteínas ligadoras de calcio, además de otros mecanismos de señalización intracelular (35). La lectina Gal- GalNac, a través de una serie de cinasas transmembrana relacionadas (36), induce cambios estructurales en el trofozoíto y en la célula blanco a través de mecanismos no identificados, para formar una placa de anclaje (37). Después, la miosina pesada II y la actina interactúan "contrayendo" el trofozoíto, que de esta manera avanza. Durante el proceso de fagocitosis, el reconocimiento de la célula blanco depende nuevamente de la lectina (34). Así, el trofozoíto penetra rápidamente por la mucosa y la submucosa, extendiendo la invasión a capas más profundas.
Las proteasas de cisteína juegan un papel importante en esta etapa, dado que permiten degradar casi todos los componentes de la matriz extracelular y su actividad depende de la virulencia del trofozoíto. Los principales componentes destruidos por las proteasas son laminina, colágeno tipo I y IV, y fibronectina.
Lesión invasiva tardía con ulceración profunda. A medida que avanza la invasión, la úlcera se extiende profundamente en un área mayor de la submucosa, dado que el tejido subyacente al epitelio interglandular ofrece menor resistencia, lo que da lugar a las típicas úlceras en botón. Para obtener el hierro necesario para su supervivencia, el trofozoíto procesa los eritrocitos fagocitados por medio de hemoglobinasas. En este momento se aprecia una notable infiltración por células como neutrófilos, macrófagos y algunos linfocitos. Llama la atención la pasividad de los macrófagos, secundaria a la alteración del perfil de citocinas inducido por el parásito, con disminución de la secreción de IL5, IL6, IFNgamma, y FNTalfa (38). El macrófago, previamente activado con IFNgamma, es la única célula capaz de enfrentar con éxito al trofozoíto y fagocitarlo (28).
Úlcera en granulación. La proliferación de pequeños vasos indica curación de la herida y es el marcador de esta etapa. No se tienen reportes que permitan saber si esta etapa ocurre espontáneamente o depende del tratamiento (28).
MECANISMOS MOLECULARES
Lectina galactosa–N-acetil-D-galactosamina (Gal-GalNac lectin). Se trata de una molécula compuesta por un heterodímero entre una cadena pesada (170 kd) y una liviana (35/31 kd) (2, 30). Su principal papel es la adhesión celular, la cual es uno de los mecanismos patógenos más importantes, dado que si es bloqueado, el trofozoíto pierde su capacidad invasora (39). No se ha determinado si participa directamente en el proceso de lisis, aunque puede estar implicada en la activación de la caspasa humana 3 e inducir apoptosis (33). Es parte fundamental de los mecanismos de locomoción y fagocitosis del trofozoíto (32, 34-37). Dado su parecido estructural, presenta reactividad antigénica cruzada con CD-59, por lo cual inhibe la formación del complejo de ataque a membrana. E. dispar también presenta esta molécula (28) que, por lo tanto, si bien es indispensable para el proceso de invasión, no es la única responsable del mismo.
Amebaporo. Esta proteína está formada por 4 hélices alfa y existen 3 isoformas denominadas A, B, y C, de las cuales la C es la más activa. Forma canales que permiten el paso de iones, agua y moléculas pequeñas, lo cual altera el equilibrio osmótico de la célula y lleva a su lisis. Ejercen actividad citolítica in vitro contra varias líneas celulares humanas y se localizan en vesículas citoplasmáticas del trofozoíto de ambas especies. No se ha demostrado que sean secretadas por los trofozoítos, así que su actividad es intracelular para digerir las células previamente fagocitadas (1, 28, 30). Su estructura es muy similar a la de otras proteínas porinas (40).
Proteasas de cisteína . Se trata del más abundante tipo de proteasas presente en E. histolytica (1); son seis proteasas de cisteína (ehcp 1-6) y la de mayor importancia es la ehcp-5 (28, 30, 31, 41). Son muy importantes en la adquisición de nutrientes y el desarrollo del ciclo de vida, así como en la invasión del hospedero. Es conocida su actividad contra diversos componentes de la matriz extracelular, pero también se ha descrito un efecto citopático directo. Se sabe que la ehcp-5 rompe la capa de mucina en su dominio carboxi-terminal, lo cual favorece la invasión (42). Se han descrito, además, sus efectos en la evasión de la inmunidad, por degradación directa de la IgA, y en la disrupción de la IgG que confiere resistencia a la lisis mediada por complemento e interrumpe la amplificación de la respuesta inmune por degradación de anafilo-toxinas C3a y C5a. Existen diferencias entre las proteasas de cisteína de E. histolytica y de E. dispar, en lo cual puede estar la clave de la capacidad patógena de la una y la inocuidad de la otra (31).
EVASIÓN DE LA RESPUESTA INMUNE
La respuesta inmune frente a la infección por E. histolytica está dada, principalmente, por inmunidad innata inespecífica. El principal mecanismo de defensa es la secreción de mucina y, una vez éste es sorteado, se facilita el proceso de invasión. Los neutrófilos infiltran la mucosa luego de la liberación de IL8 por las células epiteliales intestinales, la cual es inducida por el trofozoíto. El reconocimiento del parásito se logra por medio de receptores similares a Toll (TLR) 2 y 4 (43). Algunos autores proponen que E. histolytica puede variar sus patrones moleculares asociados al patógeno y no ser detectada (44). El complemento es inactivado por medio de reactividad cruzada de la lectina Gal-GalNac con el CD 59, lo cual lleva al bloqueo en la formación del complejo de ataque a membrana, y las proteasas de cisteína degradan las anafilotoxinas (45). La lectina de 220-kd de E. histolytica causa una disminución importante de IL5, IL6, IFNgamma, y FNTalfa, con lo cual se genera un efecto antiinflamatorio, atrayendo macrófagos inactivos o inactivando los más cercanos (38). Esto lleva a que el macrófago no pueda fagocitar adecuadamente los trofozoítos y a que no se pueda iniciar una respuesta inmune específica, la cual, si ocurre, lleva a la producción de IgA de mucosas y de IgG sérica dirigida a la lectina Gal-GalNac. La inmunidad adquirida es de corta duración, con protección de 86% a un año (4) pero inferior a 43% a los cuatro años (46).
MANIFESTACIONES CLÍNICAS
La infección amebiana presenta un amplio espectro de manifestaciones clínicas: colonización asintomática que ocurre hasta en 90% de los casos; la amebiasis intestinal (entre 9% y 9,9% de los casos), que va desde la colitis amebiana hasta la peritonitis; o las amebiasis extraintestinales, entre las cuales se destacan la amebiasis cutánea y el absceso hepático que ocurre en menos de 1% de los casos (9).
En la tabla 1 se presenta un resumen de las características asociadas a la colitis amebiana y cuáles debemos buscar en la anamnesis y en las pruebas de laboratorio (47).

Entre los diagnósticos diferenciales infecciosos es importante considerar la infección por Shigella, Salmonella, Campylobacter, Escherichia coli o Balantidium coli y, entre los no infecciosos, la enfermedad inflamatoria intestinal, la colitis isquémica, la diverticulitis y las malformaciones arteriovenosas. En el caso particular de sospecha de ameboma se debe descartar el cáncer de colon.
DIAGNÓSTICO
Durante mucho tiempo la confirmación parasitológica del diagnóstico se realizó por estudios microscópicos para visualizar la morfología del protozoario. Sin embargo, el descubrimiento de E. dispar, la cual es idéntica morfológicamente a E. histolytica, resaltó las deficiencias de este método. En su lugar están surgiendo un número importante de pruebas basadas en la detección de antígenos y anticuerpos, y algunas de ellas diferencian entre las dos especies.
La tabla 2 compara la sensibilidad y la especificidad de los diversos métodos, dejando en claro que para nuestras condiciones la prueba de detección de antígenos en heces sería la de mayor utilidad, dado que diferencia E. histolytica, es fácil de realizar y es la de más bajo costo. La serología podría ser útil para determinar la prevalencia y las demás son costosas o son complicadas (2). La reacción en cadena de la polimerasa (PCR) presenta un excelente rendimiento, con una sensibilidad mayor para la PCR de tiempo real; además, puede diferenciar infección por E. dispar específicamente (14, 48).
TRATAMIENTO
El tratamiento de la infección depende del diagnóstico clínico. Un paciente con un cuadro de colitis amebiana no requiere el mismo tratamiento que un portador asintomático, debido a los sitios y mecanismos de acción de los medicamentos empleados. Éstos se suelen dividir en luminales, como las 8-hidroxiquinolinas halogenadas (yodoquinol) y las amidas (teclozán, etofamida, quinfamida, etc.), o tisulares, como los nitroimidazoles (metronidazol, secnidazol, ornidazol). El metronidazol presenta una acción mixta, es decir, tanto luminal como tisular (9, 39).
Los medicamentos disponibles en Colombia y los esquemas de tratamiento se resumen en la tabla 3. Se recomienda el tratamiento de los pacientes asintomáticos como una medida para controlar la transmisión, pero no existen estudios que validen esta recomendación.
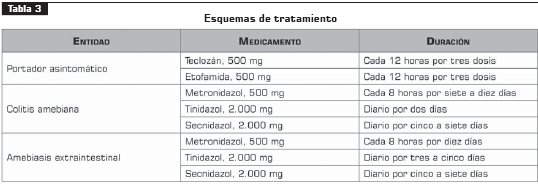
Algunos casos requieren manejo quirúrgico, como en la colitis amebiana fulminante (hemicolectomías o, incluso, colectomías totales), apendicitis amebiana (cuyo diagnóstico suele ser posquirúrgico), la perforación intestinal y el ameboma. Aunque se han descrito mecanismos de resistencia al metronidazol (49), la respuesta clínica suele ser adecuada.
INMUNIDAD Y PREVENCIÓN
Dada la morbilidad y la mortalidad asociadas a la amebiasis, se ha investigado la posibilidad de desarrollar una vacuna. La mayoría de los esfuerzos se centra en generar una respuesta humoral contra la lectina Gal-GalNac por medio de la IgA, dado que disminuye hasta en 86% la incidencia de amebiasis hasta dos años después de su aplicación (50). Hay avances en este campo, pero todavía no hay una vacuna disponible para ser evaluada en seres humanos (51).
Agradecimientos
Le agradecemos al doctor Benito Serrano del Departamento de Patología por las imágenes facilitadas para la figura 3.
Correspondencia:
Jorge Alberto Cortés, Universidad Nacional de Colombia, Facultad de Medicina, oficina 318, Bogotá, D.C., Colombia.
Carrera 30 Nº 45-03, Teléfono: 615 3546.
jorgecortes@yahoo.com
REFERENCIAS
1. Petri WA Jr. Pathogenesis of amebiasis. Curr Opin Microbiol. 2002; 5:443-7. [ Links ]
2. Tanyuksel M, Petri WA Jr. Laboratory diagnosis of amebiasis. Clin Microbiol Rev. 2003;16:713-29. [ Links ]
3. Lösch FA. Massive development of amebas in the large intestine. Translation from the original in Russian, 1875. Am J Trop Med Hyg. 1975;24:383-92. [ Links ]
4. Brumpt E. Etude sommaire de l " Entamoeba dispar" n. sp. Amibe a`kystes quadrinucleés, parasite de lhomme. Bull Acad Med. (Paris) 1925;94:943-52. [ Links ]
5. Diamond LS. Axenic cultivation of Entamoeba histolytica. Science. 1961;134:336-7. [ Links ]
6. Sargeaunt PG, Williams JE, Greene JD. The differentiation of invasive and non-invasive Entamoeba histolytica by isoenzyme electrophoresis. Trans R Soc Trop Med Hyg. 1978;72:519-21. [ Links ]
7. Diamond LS, Clark CG. A redescription of Entamoeba histolytica Schaudinn, 1903 (Emended Walker, 1911) separating it from Entamoeba dispar Brumpt, 1925. J Eukaryot Microbiol. 1993;40:340-4. [ Links ]
8. WHO/PAHO/UNESCO report. A consultation with experts on amoebiasis. Mexico City, Mexico 28-29 January, 1997. Epidemiol Bull. 1997;18:13-4. [ Links ]
9. Haque R, Huston CD, Hughes M, Houpt E, Petri WA Jr. Amebiasis. N Engl J Med. 2003;348:1565-73. [ Links ]
10. Loftus B, Anderson I, Davies R, Alsmark UC, Samuelson J, Amedeo P et al. The genome of the protist parasite Entamoeba histolytica. Nature. 2005;433:865-8. [ Links ]
11. Petri WA Jr, Haque R, Lyerly D, Vines RR. Estimating the impact of amebiasis on Health. Parasitol Today. 2000;16:320-1. [ Links ]
12. Meyer IH. Smearing the queer: medical bias in the medical care of gay men (commentary). Int J Epidemiol. 2002;31:500-3. [ Links ]
13. Walsh JA. Problems in recognition and diagnosis of amebiasis: estimation of the global magnitude of morbidity and mortality. Rev Inf Dis. 1986;8:228-38. [ Links ]
14. Ramos F, Moran P, González E, García G, Ramiro M, Gómez A et al. High prevalence rate of Entamoeba histolytica asymptomatic infection in a rural Mexican community. Am J Trop Med Hyg. 2005;73:87-91. [ Links ]
15. Ashbolt NJ. Microbial contamination of drinking water and disease outcomes in developing regions. Toxicology. 2004;198:229-38. [ Links ]
16. Barwick RS, Uzicanin A, Lareau S, Malakmadze N, Imnadze P, Iosava M et al. Outbreak of amebiasis in Tbilisi, Republic of Georgia, 1998. Am J Trop Med Hyg. 2002;67:623-31. [ Links ]
17. Lewthwaitea P, Gillb GV, Hartb CA, Beechingb NJ. Gastrointestinal parasites in the immunocompromised. Curr Opin Infect Dis. 2005; 18:427-35. [ Links ]
18. Flórez AC, García DA, Moncada L, Beltrán M. Prevalencia de microsporidios y otros parásitos intestinales en pacientes con infección por VIH, Bogotá, 2001. Biomédica. 2003;23:274-82. [ Links ]
19. Lowther SA, Dworkin MS, Hanson DL. Entamoeba histolytica/ Entamoeba dispar infections in human immunodeficiency virus–infected patients in the United States. Clin Infect Dis. 2000;30:955-9. [ Links ]
20. Tsai JJ, Sun HY, Ke LY, Tsai KS, Chang SY, Hsieh SM et al. Higher seroprevalence of Entamoeba histolytica infection is associated with human immunodeficiency virus type 1 infection in Taiwan. Am J Trop Med Hyg. 2006;74:1016-9. [ Links ]
21. Houpt ER, Glembocki DJ, Obrig TG, Moskaluk CA, Lockhart LA, Wright RL et al.The mouse model of amebic colitis reveals mouse strain susceptibility to infection and exacerbation of disease by CD4 T Cells. J Immunol. 2002;169:4496-503. [ Links ]
22. Bredin C, Margery J, Bordier L, Mayaudon H, Dupuy O, Vergeau B, Bauduceau B. Diabetes and amoebiasis: a high risk encounter. Diabetes Metab. 2004;30:99-102. [ Links ]
23. Perret C, Harris P, Rivera M, Vial P, Duarte I, Barriga F. Refractory enteric amebiasis in pediatric patients with acute graft- versus-host disease after allogeneic bone marrow transplantation. J Pediatr Gastroenterol Nutr 2000;31:86-90. [ Links ]
24. Botero J, Castaño A, Montoya M, Ocampo N, Hurtado MI, Lopera MM. A preliminary study of the prevalence of intestinal parasites in immunocompromised patients with and without gastrointestinal manifestations. Rev Inst Med Trop S Paulo. 2003:45:197-200. [ Links ]
25. Gómez CA, Cortés JA, Cuervo S, Bermúdez D, Martínez T, Arroyo P. Hallazgos clínicos, epidemiológicos y desenlace de los pacientes con neutropenia febril, Instituto Nacional de Cancerología, Bogotá, Colombia, 2003. Infectio. 2006;10:105. [ Links ]
26. Reyes P, Cortés JA, Potdevin G, Urdaneta AM, Rosales J, Cuervo SI et al. Infecciones en pacientes con trasplante de médula ósea en el trópico. Infectio. 2006;10:101. [ Links ]
27. Cardona AF, Combariza, JF, Ospina EG, Poveda CM, Ruiz CA, Ramos P, Aponte DM. Características clínicas y microbiológicas de la colitis neutropénica en adultos con neoplasias hematológicas del Instituto Nacional de Cancerología de Bogotá, D.C. (Colombia). Enferm Infecc Microbiol Clin. 2004;22:462-6. [ Links ]
28. Espinosa-Cantellano M, Martínez-Palomo A. Pathogenesis of intestinal amebiasis: from molecules to disease. Clin Microbiol Rev. 2000; 13:318-31. [ Links ]
29. Moncada D, Keller K, Chadee K. Entamoeba histolytica cysteine proteinases disrupt the polymeric structure of colonic mucin and alter its protective function. Infect Immun. 2003;71:838-44. [ Links ]
30. Huston CD. Parasite and host contributions to the pathogenesis of amebic colitis. Trends Parasitol. 2004;20:23-6. [ Links ]
31. Que X, Reed SL. Cysteine proteinases and the pathogenesis of amebiasis. Clin Microbiol Rev. 2000;13:196-206. [ Links ]
32. Ackers JP, Mirelman D. Progress in research on Entamoeba histolytica pathogenesis. Curr Opin Microbiol. 2006;9:367-73. [ Links ]
33. Huston CD, Boettner DR, Miller-Sims V, Petri WA Jr. Apoptotic killing and phagocytosis of host cells by the parasite Entamoeba histolytica. Infect Immun. 2003;964-72. [ Links ]
34. Marion S, Guillen N. Genomic and proteomic approaches highlight phagocytosis of living and apoptotic human cells by the parasite Entamoeba histolytica. Int J Parasitol. 2006;36:131-9. [ Links ]
35. Meza I, Talamas-Rohana P, Vargas MA. The cytoskeleton of Entamoeba histolytica: structure, function, and regulation by signaling pathways. Arch Med Res. 2006;37:234-43. [ Links ]
36. Beck DL, Boettner DR, Dragulev B, Ready K, Nozaki T, Petri WA Jr. Identification and gene expression analysis of a large family of transmembrane kinases related to the Gal/GalNAc lectin in Entamoeba histolytica. Eukaryot Cell. 2005;4:722-32. [ Links ]
37. Tavares P, Rigothier MC, Khun H, Roux P, Huerre M, Guillen N. Roles of cell adhesion and cytoskeleton activity in Entamoeba histolytica pathogenesis: a delicate balance. Infect Immun. 2005;73:1771-8. [ Links ]
38. Bekker-Méndez VC, Pérez-Castillo VL, Rico-Rosillo MG, Pérez- Rodríguez M, Arellano-Blanco J, Kretschmer-Schmid RR, Talamas-Rohana P. Downregulation of selected cytokines in amebiasis. Arch Med Res. 2006;37:556-8. [ Links ]
39. Guerrant RL, Van Gilder T, Steiner TS. Thielman NM, Slutsker L, Tauxe R, et al. practice guidelines for the management of infectious diarrhea. Clin Infect Dis. 2001;32:331-50. [ Links ]
40. Hecht O, Van Nuland NA, Schleinkofer K, Dingley AJ, Bruhn H, Leippe M, Grotzinger J. Solution structure of the pore-forming protein of Entamoeba histolytica. J Biol Chem. 2004;279:17834-41. [ Links ]
41. Tillack M, Nowak N, Lotter H, Bracha R, Mirelman D, Tannich E, Bruchhaus I. Increased expression of the major cysteine proteinases by stable episomal transfection underlines the important role of EhCP5 for the pathogenicity of Entamoeba histolytica. Mol Biochem Parasitol. 2006;149:58-64. [ Links ]
42. Lidell ME, Moncada DM, Chadee K, Hansson GC. Entamoeba histolytica cysteine proteases cleave the MUC2 mucin in its C-terminal domain and dissolve the protective colonic mucus gel. Proc Natl Acad Sci USA. 2006;103:9298-303. [ Links ]
43. Maldonado-Bernal C, Kirschning CJ, Rosenstein Y, Rocha LM, Rios- Sarabia N, Espinosa-Cantellano M et al. The innate immune response to Entamoeba histolytica lipopeptidophosphoglycan is mediated by toll-like receptors 2 and 4. Parasite Immunol. 2005;27:127-37. [ Links ]
44. Campos-Rodríguez R, Jarillo-Luna A. The pathogenicity of Entamoeba histolytica is related to the capacity of evading innate immunity. Parasite Immunology. 2005;27:1-8. [ Links ]
45. Evering T, Weiss LM. The immunology of parasite infections in immunocompromised hosts. Parasite Immunol. 2006;28:549-65. [ Links ]
46. Haque R, Mondal D, Duggal P, Kabir M, Roy S, Farr BM et al. Entamoeba histolytica infection in children and protection from subsequent amebiasis. Infect Immun. 2006;74:904-9. [ Links ]
47. Petri WA Jr., Singh U. Diagnosis and management of amebiasis. Clin Infect Dis. 1999;29:1117-25. [ Links ]
48. Haque R, Petri WA Jr. Diagnosis of amebiasis in Bangladesh. Arch Med Res. 2006;37:273-6. [ Links ]
49. Bansal D, Sehgal R, Chawla Y, Malla N, Mahajan RC. Multidrug resistance in amoebiasis patients. Indian J Med Res. 2006;124:189-94. [ Links ]
50. Houpt E, Barroso L, Lockhart L, Wright R, Cramer C, Lyerly D, Petri WA Jr. Prevention of intestinal amebiasis by vaccination with the Entamoeba histolytica Gal/GalNac lectin. Vaccine. 2004;22:611-7. [ Links ]
51. Snow MJ, Stanley SL Jr. Recent progress in vaccines for amebiasis. Arch Med Res. 2006;37:280-7. [ Links ]

