










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkInfectio
Print version ISSN 0123-9392
Infect. vol.12 no.3 Bogotá July/Sept. 2008
1 Grupo de Gastrohepatología, Universidad de Antioquia, Medellín, Colombia
2 Ciencias Básicas Biomédicas, Universidad de Antioquia, Medellín, Colombia
3 Facultad de Medicina, Universidad de Antioquia, Medellín, Colombia
Fecha de recibido: 08/02/2008. Fecha de aceptación: 08/05/2008
Resumen
Entre 500.000 y 1000.000 de personas mueren anualmente en todo el mundo a causa de complicaciones relacionadas con la infección por el virus de la hepatitis B. Este virus presenta la tasa de mutación más alta entre los virus ADN que infectan al hombre. Aunque algunos de los genotipos del virus de la hepatitis B se han asociado al desarrollo de complicaciones durante la infección persistente, los estudios recientes sugieren un papel más importante a la aparición de ciertas variantes de este virus durante la infección. Una de las regiones del genoma del virus de la hepatitis B más analizadas y que presentan mayor frecuencia de mutaciones, es la región precore/core y el promotor correspondiente; la doble mutación (A1762T/G1764A) del promotor es la de mayor impacto sobre la replicación del virus de la hepatitis B y la respuesta a la terapia antiviral.
En esta revisión se muestra el papel del genotipo y las variantes de la región precore/core del virus de la hepatitis B en el contexto del curso clínico y el tratamiento antiviral.
Palabras clave: virus de la hepatitis B, genotipos de virus de la hepatitis B, variantes precore/core, cirrosis, carcinoma hepatocelular.
Abstract
It is estimated that among 500,000 to 1 million deaths occurred annually worldwide due to complications associated to hepatitis B virus infection (HBV).
The HBV has the higher mutation rate among DNA viruses that infect humans. Although some HBV genotypes have been associated with clinical complications during chronic infection, recently published data suggest a stronger role of HBV variants.
Precore/core region and its promoter element are one of the HBV genomic segments with a higher mutation rate. The double mutation (A1762T/G1764A) in basal core promoter sequence results in dramatic viral replication changes and poor response to antiviral treatment.
This review summarizes the present knowledge about the genotype and precore/core variants influence in clinical outcome, immune response evasion and antiviral therapy.
Key words: hepatitis B virus, HBV genotypes, precore/core variants, cirrhosis, hepatocellular carcinoma .
Introducción
La infección por el virus de la hepatitis B es uno de los problemas de mayor impacto en salud pública, dada su amplia distribución a nivel mundial y su relación directa con el desarrollo de cirrosis y carcinoma hepatocelular. Se estima que 350 millones de personas están crónicamente infectados y que cada año fallecen entre 500.000 a 1’000.000 de individuos por complicaciones asociadas a la infección por el virus de la hepatitis B1,2.
Según la prevalencia en la población general del antígeno de superficie del virus de la hepatitis B (HBsAg), se han determinado regiones de baja (menos de 2%), intermedia (2 a 8%) y altas (más de 8%) endemias; el suroeste asiático, el áfrica subsahariana y la cuenca amazónica corresponden a las regiones de mayores prevalencias en el mundo2.
En la actualidad, se han caracterizado 8 genotipos, designados con las letras A-H, con una divergencia de 8%. Los genotipos del virus de la hepatitis B presentan un patrón de distribución geográfica más o menos definido. El genotipo F, por ejemplo, circula en América, con diferente prevalencia según el país. El genotipo H, el más recientemente reportado en la literatura, es prevalente en Centroamérica3.
Algunos estudios sugieren que el genotipo viral estaría relacionado con la evolución clínica de la infección y el riesgo de desarrollo de carcinoma hepatocelular asociado al virus de la hepatitis B. Según los resultados de algunos trabajos, los genotipos C y F están relacionados con el desarrollo de complicaciones en el curso de la infección crónica por el virus de la hepatitis B; sin embargo, los estudios recientes sugieren que el factor determinante del curso clínico serían las variantes en la región precore/core del genoma del virus de la hepatitis B, más que el genotipo4.
En esta revisión se describen los aspectos más relevantes de la variabilidad genética del virus de la hepatitis B, en términos de genotipos, subgenotipos y variantes, y las implicaciones en epidemiología, curso clínico y respuesta antiviral.
Virus de la hepatitis B
Genoma. El virus de la hepatitis B pertenece a la familia Hepadnaviridae, género ortohepadnavirus. El virus de la hepatitis B posee un ADN circular de doble cadena parcial, de 3,2 kb de longitud, aproximadamente. Presenta cuatro promotores, dos regiones potencia-doras (enhancer) y cuatro marcos de lectura abierta (open reading frame, ORF) superpuestos, entre ellos la región pre-S1/pre-S2/S, que codifica para los tres tipos, L, M y S del HBsAg, la región P, que codifica la polimerasa viral (Pol), la región X, que codifica la proteína transactivadora X (HBx) y la región pre-core/ core que codifica la proteína core, unidad estructural de la cápside (HBcAg) y el antígeno e (HBeAg)1 (figura 1).
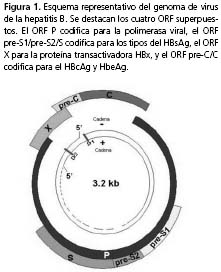
Segmento pre-core/core y su región promotora. El segmento pre-core/core tiene una extensión aproximada de 639 nucleótidos; presenta dos codones de iniciación (ATG) alternativos en las posiciones 1.814 y 1.901, un codón de terminación (TAG) en la posición 2.4505,6, y dos elementos reguladores que se mencionan más adelante (figura 2). Las mutaciones en el segmento pre-core/core y en el promotor, se han relacionado con el desarrollo de complicaciones en el curso de la infección por el virus de la hepatitis B4,6,7.
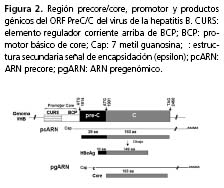
Durante la replicación del virus de la hepatitis B se generan tres transcritos subgenómicos, de 2,4 y 2,1 kb, para la expresión de HBsAg, y de 0,7 kb para HBx. Igualmente, otros dos transcritos genómicos de 3,5 kb, ARN precore (pcARN) y ARN pregenómico (pgARN). El pcARN da lugar a la expresión del HBeAg, antígeno que sufre modificaciones posteriores a la transducción mediadas por proteasas celulares; el pgARN posee doble función: corresponde al molde para la síntesis de nuevos genomas y es traducido a las proteínas HBcAg y Pol (figura 2)1,6.
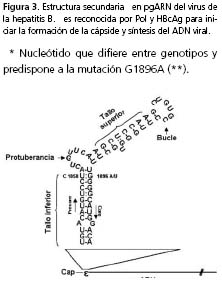
El pgARN sirve como molde debido a la presencia de una estructura generada por plegamiento en su extremo 5’, reconocida por Pol y HBcAg, que permite su información de cápside y el inicio de la síntesis de ADN. Esta estructura secundaria se denomina ε (épsilon); se genera por plegamiento de ciertas regiones del pgARN; se encuentra conformada por dos tallos (inferior y superior) separados por una protuberancia asimétrica y un bucle apical (figura 3). Las mutaciones en la región pre-core/ core no sólo afectan la expresión de HBcAg y HBeAg, sino también la continuidad del ciclo de replicación viral6.
La transcripción de pgARN y pcARN está regulada por un promotor que posee dos ele-mentos “corriente arriba” (upstream) de la región pre-core/core5,8,9: un elemento proximal entre los nucleótidos 1.744-1.851, denominado promotor básico de core (basal core promoter, BCP), y un elemento distal entre los nucleótidos 1.636-1.703, conocido como la secuencia reguladora “corriente arriba” de core (core upstream regulatory sequence, CURS)5,10-12 (figura 2).
Debido al papel fundamental de los transcritos regulados por BCP, las mutaciones en este promotor pueden provocar cambios considerables de la actividad de replicación del virus de la hepatitis B.
Genotipos del virus de la hepatitis B
Clasificación del Virus de la hepatitis B: subtipos y genotipos. Para el virus de la hepatitis B, se han caracterizado 8 genotipos, designados con las letras A-H, con una divergencia genética de 8% o más13,14. Los geno-tipos pueden determinarse por secuenciación del genoma completo o del ORF S y por ensayos de RFLP (restriction fragment lenght polimorfism) de las ORF S o pre-core/core13,15. En la figura 4 se muestra un árbol sin raíz que refleja la relación filogenética entre los virus de la hepatitis B aislados de humanos y de primates (simios). Los genotipos F y H son los más divergentes del grupo de virus que infectan a los humanos; sin embargo, están relacionados con un hepadnavirus que infecta monos del nuevo mundo (mono lanudo, Lagothrix lagotricha)13,14; actualmente, este hallazgo es tema de discusión en la definición del origen del virus de la hepatitis B.
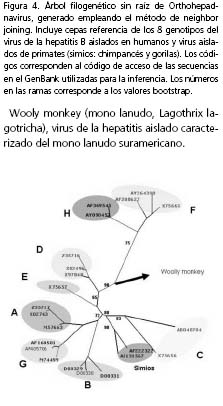
Wooly monkey (mono lanudo, Lagothrix lagotricha), virus de la hepatitis aislado caracterizado del mono lanudo suramericano.
La primera clasificación propuesta para el virus de la hepatitis B se basó en la tipificación de los aislamientos según diferencias serológicas; se utilizan los términos serotipo o subtipo para esta clasificación. Los subtipos se denominan ayw1, ayw2, ayw3, ayw4, ayr, adw2, adw3, adw4q-, adr y adrq-; la primera letra corresponde a la región antigénica muy conservada, presente en los tres tipos de HBsAg denominada determinante a. La designación d/y es mutuamente excluyente y obedece a la presencia del aminoácido lisina (Lis) o arginina (Arg) en la posición 122 del HBsAg. Las letras w/r corresponden al aminoácido Lis/Arg en la posición 16016. La asignación w1-4 se basa en la identidad del aminoácido en la posición 127; prolina (Pro) w1/2, treonina (Tre) w3 y leucina (Leu) w4. El subtipo q- corresponde a los aminoácidos alanina (Ala) y glicina (Gln) en la posición 177 y 1786,17 (tabla 1).
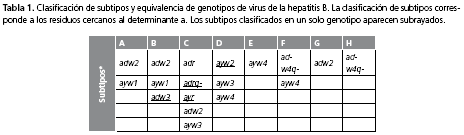
Existe cierta equivalencia entre la designación serológica de subtipos y genotipos del virus de la hepatitis B. Por ejemplo, adw3 se encuentra incluido en el genotipo B, adr, adrq- y ayr pertenecen al genotipo C, ayw2 corresponde al genotipo D; sin embargo, hay subtipos descritos en más de dos genotipos, como es el caso de adw2 que se encuentra en los genotipos A, B, C y G18.
Debido a la gran variabilidad del virus de la hepatitis B, a la mejora de los métodos de inferencia filogenética y a la factibilidad de la secuenciación automatizada, la clasificación por genotipos es la de referencia.
Distribución global genotipos y subgenotipos. Los genotipos del virus de la hepatitis B presentan un patrón de distribución geográfica más o menos específica (figura 5). Los aislamientos pertenecientes al genotipo A provienen de Europa, Norteamérica, áfrica e India, clasificados en dos subgenotipos principales, denominados africano-asiático (A1) y europeo-norteamericano (A2). Recientemente, se ha reconocido un nuevo subgenotipo denominado A3, aislado inicialmente en Camerún y, actualmente, también en Mali y Gambia18-20.
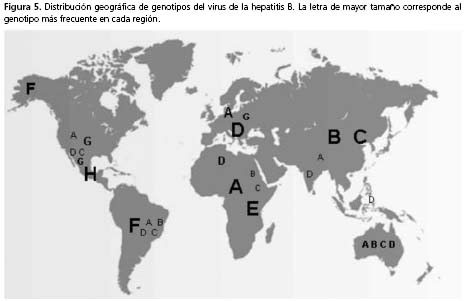
El genotipo B circula principalmente en Asia. Se han caracterizado cuatro subgenotipos, designados B1-B4. En el subgenotipo B1 se encuentran cepas aisladas en Japón; en B2, cepas de China, Vietnam y Tailandia; en B3, cepas de Indonesia, y en B4, cepas provenientes de Vietnam21-23.
Igualmente, los virus agrupados dentro del genotipo C son aislamientos principalmente del continente asiático. Se han descrito cuatro subgenotipos, C1-C4. Al subgenotipo C1 pertenecen virus aislados en Japón, Corea y China; al C2, cepas de Laos, Bangladesh, China y Tailandia; al C3, cepas de Nueva Zelanda, Polinesia y Japón, y al C4 aislados en Australia, de aborígenes16,17,24.
El genotipo D es el de más amplia distribución. En éste se han descrito cuatro subgenotipos, designados D1 a D4. En el subgenotipo D1 se encuentran aislamientos de China y Japón; en el D2, cepas detectadas en Japón e India; en el D3, aislamientos de Alaska y Sudáfrica, mientras que en el D4, virus aislados en islas del Pacífico Sur y Somalia18,25.
África presenta la frecuencia más alta de detección del genotipo E. No se han descrito subgenotipos; se han identificado aislamientos recientes en inmigrantes africanos residentes en países como Francia, Alemania y Japón 26,27.
El genotipo F circula en Suramérica, con diferentes prevalencias entre los países de la región. Numerosos aislamientos caracterizados en tribus amerindias pertenecen a este genotipo; ya que estos grupos de población son cerrados y sus asentamientos se localizan en regiones apartadas, la evidencia sugiere que este geno-tipo es autóctono del continente; sin embargo, esto sigue siendo tema de discusión28,29.
El genotipo F también se ha identificado en pacientes de Francia, Holanda, Italia, España, y Polonia30. En este genotipo se han descrito los subgenotipos F1, F2, F3 y F4, con divergencia de 4,3% al 6,1%. Para F1 se ha propuesto la subdivisión F1a y F1b; F1a es prevalente en el norte de las Américas e incluye aislamientos de Alaska, El Salvador, Honduras, Guatemala, Costa Rica y Nicaragua. F1b predomina en el suroeste del continente y está constituido por cepas de Perú y Argentina. El subgenotipo F2 se ha reportado en Costa Rica, Nicaragua, Venezuela y Brasil. El subgenotipo F3 es prevalente en Panamá y la región norte de Suramérica, conformado por aislamientos de Venezuela y Colombia, mientras que el subgenotipo F4 se ha reportado hacia la parte sur del continente, con reportes en Bolivia y Argentina; a este subgenotipo también pertenecen algunos aislamientos de Francia31,30.
La distribución del genotipo G es menos conocida debido al bajo número de secuencias disponibles. A este genotipo pertenecen cepas caracterizadas en México, Estados Unidos y Francia18.
El genotipo H es el más reciente reportado en la literatura; es muy frecuente en Centroamérica. En éste se agrupan aislamientos de México, Nicaragua y California, Estados Unidos; los aislamientos en este último país están relacionados con inmigrantes centroamericanos30,32,33; a partir de 2005 se ha caracterizado, además, en donantes de sangre e individuos con hepatitis aguda y crónica en Japón34.
Como se mencionó anteriormente, en Centroamérica y Suramérica circulan predominantemente los genotipos H y F, respectivamente; sin embargo, en Brasil y Argentina se ha reportado con frecuencia la detección del genotipo A; aunque también se han descrito los genotipos B, C, D y G en países de la región 13.
Recientemente, en Brasil se clasificó una cepa como genotipo E, proveniente en un paciente originario de Angola; por lo tanto, el caso fue considerado no autóctono35. Este reporte evidencia la diseminación de los genotipos del virus de la hepatitis B que se consideran circunscritos a ciertas regiones del mundo, al igual que la influencia de la actividad humana en la evolución y epidemiología de la infección por virus de la hepatitis B.
En Colombia sólo se ha publicado un estudio de genotipificación del virus de la hepatitis B hasta el momento; éste fue realizado en 47 donantes de sangre de Bogotá y Bucaramanga, y en la mayoría se encontró el genotipo F (86%), seguido por los genotipos D (8%), A (2%), C (2%) y G (2%)36. En 13 aislamientos se determinó el subgenotipo; todos correspondieron a F3. Además de esta reciente publicación, existe otro registro en la literatura del análisis filogenético de cepas de Latinoamérica, en el cual se incluye un aislamiento proveniente de un paciente colombiano; este aislamiento también se clasificó como subgenotipo F313,37.
Considerando que no es claro el papel del genotipo viral en el curso clínico de la infección por virus de la hepatitis B y el reducido número de trabajos realizados en Latinoamérica, son necesarios estudios adicionales para determinar la circulación de genotipos y subgenotipos en Colombia y el impacto de los mismos en la evolución clínica. Actualmente se está realizando un estudio de caracterización de genotipos y variantes pre-core/core en aislamientos de virus de la hepatitis B provenientes de pacientes con diagnóstico de cirrosis, carcinoma hepatocelular o ambos, en un hospital de Medellín.
Impacto del genotipo del virus de la hepatitis B
Curso clínico de la infección. Algunos factores virales son determinantes en el curso de la infección crónica por el virus de la hepatitis B38,39; varios trabajos sugieren que el genotipo del virus de la hepatitis B estaría relacionado con el riesgo de desarrollo de carcinoma hepatocelular40. En regiones donde circulan los genotipos B y C, los individuos infectados con el genotipo C tienen mayor probabilidad de desarrollar carcinoma hepatocelular41 e, igualmente, una menor supervivencia42. Un estudio prospectivo realizado en Taiwán reportó que los pacientes infectados con el genotipo C presentan viremias más altas y desarrollan carcinoma hepatocelular en mayor proporción que lo observado para otros genotipos del virus de la hepatitis B43.
En España, se encontraron diferencias en la evolución clínica de pacientes con hepatitis B crónica, según el genotipo causante de la infección. En 258 pacientes incluidos en el estudio, se reportaron frecuencias de 52%, 35% y 7% para los genotipos A, D y F del virus de la hepatitis B, respectivamente. A pesar de la baja frecuencia de infección por el genotipo F, se encontró mayor mortalidad relacionada con este genotipo en comparación con los pacientes infectados con los genotipos A y D44.
Aunque la infección por los genotipos C y F parece cursar con mayores complicaciones, los resultados de los estudios que establecen relación entre el genotipo y el curso clínico no se pueden extrapolar, principalmente porque la mayoría son trabajos descriptivos, realizados en Asia, donde circulan predominantemente los genotipos B y C, y la población en riesgo posee características genéticas particulares45,46. Es importante realizar estudios a gran escala en diferentes regiones del mundo, donde la infección por el virus de la hepatitis B es un problema de salud pública, para esclarecer el impacto del genotipo sobre el desarrollo de complicaciones.
Seroconversión del HBeAg y curso clínico. Uno de los marcadores utilizados como indicador de alta replicación viral del virus de la hepatitis B es el HBeAg; éste posee un papel en el desarrollo de la tolerancia inmunológica y el ser positivo se asocia con altos niveles de replicación, aunque no es indispensable para la producción de viriones. La seroconversión de este marcador corresponde a la detección de anticuerpos contra el HBeAg (anti-HBe) y se utiliza como indicador pronóstico de la infección, de la respuesta al tratamiento antiviral o de ambos47,48.
La función de modulación inmunológica del HBeAg se ha sugerido a partir de evidencias obtenidas en estudios experimentales y epidemiológicos. Más de 90% de los neonatos nacidos de madres seropositivas para HBsAg y HBeAg desarrollan infección persistente en ausencia del tratamiento antiviral49; en ratones que expresan de manera constitutiva el HBeAg (ratones transgénicos para el HBeAg), se ha demostrado el paso de este antígeno a través de la placenta, induciendo tolerancia a HBcAg y HBeAg47.
Al evaluar la seroconversión para el HBeAg en pacientes infectados con los genotipos C y B, los infectados con C presentan seroconversión tardíamente en comparación con los infectados por genotipo B (6 Vs. 3 años); igualmente, aquéllos con genotipo C presentaron una mayor proporción de reactivación de la infección (36 Vs. 16%)50.
En nativos de Alaska, donde se encontraron los genotipos A, B, C, D y F, se encontró que la población infectada con el genotipo C tenía seroconversión a una edad promedio mayor en comparación con la media de los otros genotipos (44,7 Vs. <20 años)51; esto sugiere un mayor periodo de tolerancia inmunológica en los pacientes infectados por el genotipo C, dada la expresión más prolongada del HBeAg, una mayor tasa de replicación viral y mayor probabilidad de transmisión perinatal.
Respuesta al tratamiento antiviral. Actualmente, no existe tratamiento que permita la resolución definitiva en todos los casos de hepatitis B; de tal modo, los objetivos del tratamiento disponible son controlar la replicación viral para disminuir la progresión y el riesgo del desarrollo de carcinoma hepatocelular52. Cinco medicamentos se utilizan en el tratamiento de los individuos con infección crónica por virus de la hepatitis B: interferón alfa (IFN-α), interferón alfa con polietilén glicos (peg-IFN-α), y medicamentos análogos a nucleótidos y nucleósidos (NuA), como la lamivudina, el adefovir y el entecavir53.
El IFN-α es producido grandes cantidades por las células del sistema inmune; actúa inhibiendo la replicación viral, aumentando el potencial lítico de las células asesinas naturales y el nivel de expresión de moléculas del complejo mayor de histocompatibilidad tipo I54. A diferencia del IFN-α, el peg-IFN-α posee mayor biodisponibilidad debido a la unión covalente con las moléculas de polietilenglicol que disminuye la tasa de excreción renal.
Por otra parte, los NuA son moléculas que ocasionan la terminación temprana de la polimerización de ADN, durante la transcripción inversa del pgARN a ADN55. Aunque los protocolos de tratamiento con NuA se difundieron rápidamente, dada su gran efectividad y mejor tolerancia en comparación con el interferón56, los estudios virológicos recientes han evidenciado la aparición de cepas resistentes57.
Al igual que con el curso clínico de la infección, se presenta cierta asociación entre el genotipo y el éxito de la terapia antiviral. Comparando la eficacia del tratamiento con IFN-ε en un grupo de individuos infectados por los genotipos A o D (46 y 35, respectivamente), se observó mejor respuesta al tratamiento en los infectados con el genotipo A, lo cual se evidencia por mejoría histológica y disminución de la carga viral58.
Igualmente, las personas infectadas por el genotipo C muestran menor respuesta al tratamiento con IFN-α, en comparación con lo observado para el genotipo B. En este mismo trabajo se observó gran frecuencia de mutaciones de pre-core/core en aislamientos del genotipo C59. De la misma forma, los pacientes tratados con lamivudina presentan una respuesta sostenida por más tiempo en individuos infectados con el genotipo B en comparación con pacientes infectados por el genotipo C del virus de la hepatitis B.
Estos resultados indican que una pobre repuesta al tratamiento está más asociada al genotipo C; esto, junto a la evidencia de que este genotipo se relaciona con mayor frecuencia al desarrollo de carcinoma hepatocelular y a mayores periodos positivos del HBeAg, señala la existencia de diferencias genéticas virales que determinan un mayor potencial virulento, lo cual sitúa a la infección por el genotipo C como la de peor pronóstico60.
Variantes de la región pre-core/ core del virus de hepatitis B
Variabilidad genética del virus de la hepatitis B. Uno de los factores más importantes que contribuye a la variabilidad del virus de la hepatitis B es la estrategia de replicación. El virus de la hepatitis B posee una enzima polimerasa con actividad transcriptasa inversa, la cual presenta homología con las transcriptasas inversas de los retrovirus61. La Pol del virus de la hepatitis B presenta una alta tasa de error, ocasionada por la ausencia de actividad correctora en sentido 3’→5’62. Se han descrito variantes del virus de la hepatitis B en individuos con hepatitis B crónica y carcinoma hepatocelular; esta variabilidad se ha propuesto como uno de los mecanismos de persistencia utilizados por el virus de la hepatitis B63,64.
Los estudios moleculares han permitido determinar las regiones del genoma del virus de la hepatitis B que presentan una mayor frecuencia de substituciones, entre ellas, la región correspondiente a pre-core/core65.
Variantes de la región pre-core/core. Se han descrito diversas mutaciones en la región pre-core/core y BCP en la literatura (figura 2). En la secuencia BCP, la doble mutación A1762T/G1764A es frecuente, lo que resulta en la substitución de una timina (T) en la posición 1.762 en reemplazo de una adenina (A), y una A en la posición 1.764 en reemplazo de una guanina (G). Estas mutaciones se han observado en aislamientos del virus de la hepatitis B provenientes de casos con hepatitis crónica, hepatocarcinoma y hepatitis fulminante66.
Los análisis experimentales demuestran que las cepas del virus de la hepatitis B con la doble mutación en BCP presentan aumento del número de moléculas de pgARN y de la síntesis de las proteínas HBcAg y Pol. Estos eventos causan un incremento en la producción del número de partículas virales67.
Para la región pre-core, la mutación de G por A en la posición 1.896 (G1896A) es frecuente. Esta variante tiene impacto en la síntesis del HBeAg y en los eventos tempranos de la replicación, dado que la región coincide con el segmento que genera la señal ε. Por un lado, la síntesis de HBeAg es interrumpida tempranamente, debido a que la substitución G1896A resulta en un codón de parada (UAG), lo que permite la traducción de una proteína truncada no funcional, que corresponde a los primeros 27 aminoácidos del extremo amino terminal. Además, este cambio puede alterar la estabilidad de la estructura secundaria y, por ende, causar ineficiencia en la formación de la cápside de pgARN6.
Papel de las variantes pre-core/core
Seroconversión del HBeAg y terapia antiviral. La disminución o la ausencia de expresión del HBeAg se han descrito en individuos con hepatitis B crónica; este evento se origina por un aumento en el título de anti-HBe. La mutación G1896A, otras substituciones y deleciones/inserciones en el segmento pre-core/core pueden generar codones de parada que podrían también desencadenar la seroconversión de este antígeno6.
Se han realizado estudios in vitro para explicar el papel de las variantes pre-core/core en la resistencia al tratamiento antiviral. En cultivos tratados con IFN-α, las construcciones que poseen la substitución G1896A se replican más eficientemente, observándose aumento de un logaritmo base 10 de la cantidad de ADN viral, mientras que los virus silvestres disminuyen su nivel de replicación en presencia de IFN-α68; resultados similares se obtienen al evaluar el comportamiento de replicación de las variantes BCP en presencia de IFN-α, encontrándose aumento de dos veces el título de virus, en comparación a la construcción sin mutaciones en BCP69. En conjunto, los resultados indican una menor sensibilidad de las cepas que portan substituciones en pre-core/core y BCP a la terapia con IFN-α.
Igualmente, se ha reportado que la variante G1896A restaura la eficacia de replicación in vitro de las construcciones transfectados en células tratadas con lamivudina; resultados similares se reportan en clones que contienen la doble mutación en BCP70.
Un comportamiento biológico distinto de las variantes pre-core/core (G1896A o A1762T/ G1764A), se ha observado ante el tratamiento in vitro con adefovir. Al no observarse disminución en la sensibilidad al adefovir en los las construcciones que portan las mutaciones descritas, el adefovir es considerado un fármaco efectivo para tratar pacientes con infección por virus de la hepatitis B que portan variantes pre-core/core71
La resistencia al tratamiento antiviral in vitro de las cepas con variantes BCP es concordante con la alta frecuencia de estas variantes en el genotipo C, y la menor eficacia de la terapia antiviral en individuos infectados con este genotipo.
Curso clínico de la infección. En el tejido hepático tumoral y no tumoral de pacientes con carcinoma hepatocelular asociado a la infección por virus de la hepatitis B72, se detectó alta proporción de virus con la mutación G1896A en el segmento pre-C; además, la doble mutación en BCP fue reportada en 16 de los 20 tejidos tumorales analizados, y en 18 de los 20 no tumorales. La alta proporción de mutaciones en el segmento pre-C y en BCP en el tejido tumoral y no tumoral, sugiere que la presencia de estas mutaciones podrían potenciar la capacidad oncogénica del virus de la hepatitis B.
C. H. Chen y colaboradores caracterizaron la evolución de las variantes pre-core/core en pacientes infectados por los genotipos B y C73. Las mutaciones G1896A y A1762T/G1764A fueron detectadas en cepas del virus de la hepatitis B genotipo C con una alta frecuencia, con aparición más temprana de las variantes en BCP que en el segmento pre-C (G1896A). En los aislamientos del genotipo B se observó lo contrario; es decir, baja frecuencia de variantes con aparición más temprana de mutaciones en pre-C. Los pacientes infectados por el genotipo C presentaron mayor riesgo del desarrollo de fibrosis hepática; este y otros estudios sugieren que las variantes pre-core/ core podrían tener un valor pronóstico en el desarrollo de carcinoma hepatocelular74-77.
Evasión de la respuesta inmune. En pacientes con hepatitis crónica se ha detectado alta frecuencia de la mutación G1896A, deleciones en la posición 1.937 y mutaciones en regiones importantes para el reconocimiento del HBcAg por los linfocitos, lo que sugiere una asociación entre estas variantes y la presión ejercida por la respuesta inmune de adaptación78.
Algunos investigadores han relacionado la presencia de variantes del segmento C con la evasión de la respuesta inmune y la persistencia de la infección por el virus de la hepatitis B79. Mediante la secuenciación del segmento C de los aislamientos del virus de la hepatitis B provenientes de pacientes con carcinoma hepatocelular, se identificaron siete dominios según la frecuencia de mutaciones80; el dominio I correspondió a mutaciones ubicadas entre los codones 1 y 21; el dominio II entre los codones 33 y 41; el dominio III entre los codones 59 y 66; el dominio IV entre los codones 80 y 121; el dominio V entre los codones 130 y 135; el dominio VI entre los codones 148 y 160, y el dominio VII entre los codones 172 y 183.
Al hacer la deducción de la secuencia de aminoácidos para el HBcAg, se logró establecer que tres de los dominios caracterizados coincidían con regiones antigénicas importantes para el reconocimiento y el establecimiento de la respuesta inmune específica; estas regiones fueron previamente descritas mediante ensayos de proliferación en células de sangre periférica obtenidas de pacientes infectados por el virus de la hepatitis B81-83. Las mutaciones puntuales observadas en la secuencia del segmento C se encontraron predominantemente en los dominios I, IV y V, y menos frecuentemente en los dominios II, III, VI y VII (dominios mayores y menores, respectivamente); las variantes en los dominios I, III y V del HBcAg coincidieron con las regiones peptídicas de reconocimiento antigénico para linfocitos T.
Genotipos del viris de la hepatitis B y frecuencia de variantes pre-C/C
Los genotipos C y F han sido relacionados al desarrollo de complicaciones en el curso de la infección crónica. Algunos estudios sugieren que el factor determinante del curso clínico sería la presencia de variantes pre-core/core más que el genotipo. Un ejemplo es lo reportado para el nucleótido en la posición 1.858 del pgARN; éste establece puentes de hidrógeno con el nucleótido de la posición 1.896 estabilizando el tallo inferior y, por lo tanto, la estructura secundaria de la señal de formación de cápside ε (figura 3).
Si en el pgARN correspondiente a un genotipo, el nucleótido de la posición 1.858 es uracilo (U), como es el caso de los subgenotipos F1 y F431, la mutación G1896A aumenta la estabilidad de la estructura secundaria ε. Por otro parte, si está presente una C en la posición 1.858, la mutación G1896A genera un codón de parada, como en el primer caso, y, además, altera la conformación de la estructura secundaria ε, con un efecto deletéreo en la formación de la cápside de pgRNA y el inicio de la síntesis del ADN viral. Es así como en aislamientos de virus de la hepatitis B que corresponden al genotipo A, C, H y los subgenotipos F2 y F3 que presentan una C en la posición 1858, es menos frecuente encontrar G1896A31,84,85 el desarrollo de la mutante , manteniendo una expresión de HBeAg por un tiempo mayor relacionada con estadios activos de replicación y modulación inmunológica.
Un trabajo realizado por Arauz-Ruiz y colaboradores reportó alta frecuencia de mutaciones en la región pre-C para el genotipo F e, igualmente, mutaciones no sinónimas en los codones que codifican para los aminoácidos 57-68 del HBcAg86; esta región coincide con el dominio III reportado previamente por Ozono y colaboradores80 y que, como se mencionó, es una de las regiones más importantes para el reconocimiento antigénico de linfocitos T.
Resultados similares fueron observados por França y colaboradores en Argentina, donde circulan los subgenotipos F1 y F4; en este estudio se describió alta frecuencia de la mutación G1896A en las cepas pertenecientes a los genotipos F y D, y la presencia de T en la posición 1.958; U si corresponde al pgARN30,87.
Asimismo, algunos investigadores han reportado que el virus de la hepatitis B genotipo B tiene una mayor frecuencia de mutaciones en la región pre-C, mientras que el genotipo C en BCP. Esto se relaciona con la seroconversión más temprana observada en cepas genotipo B y con la mayor tasa de replicación en cepas del genotipo C, lo que sugiere que las substituciones en BCP conllevan a un mayor potencial virulento del genotipo infeccioso, ya sea por pérdida de la regulación de la proporción de transcripción pcARN/pgARN o por el efecto potencial de estas mutaciones en el gen X con el cual BCP se superpone88,89.
Conclusión
Existe un número considerable de hallazgos epidemiológicos y estudios científicos que sugieren una relación entre el genotipo y las variantes pre-core/core del virus de la hepatitis B con el curso clínico de la infección y la respuesta al tratamiento antiviral.
La infección por el genotipo C y la aparición de la doble mutación A1762T/G1764A en BCP se consideran las condiciones de peor pronóstico. La determinación de causalidad se hace difícil debido a que la mayoría de los trabajos publicados son estudios descriptivos y realizados –en su mayoría– en países asiáticos con circulación predominante de ciertos genotipos, características socioculturales particulares y diferencias genéticas de población que dificultan la extrapolación de los resultados.
Es así como se hace necesaria la realización de estudios a gran escala de tipo analítico y experimental, con el fin de identificar el impacto de los genotipos del virus de la hepatitis B y las variantes en la historia natural de la infección y el tratamiento antiviral.
Por último, resulta importante esclarecer la asociación que podría existir entre la infección por el genotipo F, o sus subgenotipos, y el riesgo de desarrollar carcinoma hepatocelular, ya que es el genotipo predominante en Suramérica, y que los estudios clínicos en España y Alaska lo asocian con altas tasas de mortalidad y con el desarrollo de complicaciones.
Estrategia de búsqueda de la bibliografía. Los artículos utilizados para la elaboración del manuscrito fueron obtenidos a través de las bases de datos PubMed y MedLine, utilizando la siguiente combinación de palabras: virus de la hepatitis B, genotipos, variantes precore/core, carcinoma hepatocelular, curso clínico, tratamiento antiviral y eficacia del tratamiento.
Agradecimientos
Los autores del manuscrito agradecen a la Universidad de Antioquia; esta revisión hace parte de los compromisos adquiridos con la Vicerrectoría de Investigaciones de esta institución.
Correspondencia.
Carrera 53 Nº 61-30, Grupo de Gastrohepatología, Laboratorio 434, torre 2 Sede de Investigaciones Universitarias, Universidad de Antioquia. Medellín, Colombia. Teléfono: (574) 210 6573; fax: (574) 210 6565 mcnavasn@gmail.comBibliografía
1. Lee WM. Hepatitis B virus infection. N Engl J Med. 1997;337:1733-45. [ Links ]
2. Lavanchy D. Worldwide epidemiology of HBV infection, disease burden, and vaccine prevention. J Clin Virol. 2005;34(Suppl.1):S1-3. [ Links ]
3. Allain JP. Epidemiology of hepatitis B virus and genotype. Clin Virol. 2006;36(Suppl.1):S12-7. [ Links ]
4. Hunt CM, McGill JM, Allen MI, Condreay LN. Clinical relevance of hepatitis B viral mutations. Hepatology. 2000;3:1037-44. [ Links ]
5. Yuh CH, Chang YL, Ting LP. Transcriptional regulation of precore and pregenomic RNAs of hepatitis B virus. J Virol. 1992;66:4073-84. [ Links ]
6. Kidd-Ljunggren K, Miyakawa Y, Kidd AH. Genetic variability in hepatitis B viruses. J Gen Virol. 2002;83:267-80. [ Links ]
7. Cho SW, Shin YJ, Hahm KB, Jin JH, Kim YS, Kim JH, Kim HJ. Analysis of the precore and core promoter DNA sequence in liver tissues from patients with hepatocellular carcinoma. J Korean Med Sci. 1999;14:424-30. [ Links ]
8. Roossinck MJ, Jameel S, Loukin SH, Siddiqui A. Expression of hepatitis B viral core region in mammalian cells. Mol Cell Biol. 1986;6:1393-400. [ Links ]
9. Bock CT, Malek NP, Tillmann HL, Manns MP, Trautwein C. The enhancer I core region contributes to the replication level of hepatitis B virus in vivo and in vitro. J Virol. 2000;74:2193-202. [ Links ]
10. Galibert F, Mandart E, Fitoussi F, Charnay P, Galibert F. Nucleotide sequence of the hepatitis B virus genome (subtype ayw) cloned in E. coli. Nature 1979;281:646-50. [ Links ]
11. Pasek M, Goto T, Gilbert W, Zink B, Schaller H, MacKay P, Leadbetter G, Murray K. Hepatitis B virus genes and their expression in E. coli. Nature. 1979;282:575-9. [ Links ]
12. Valenzuela P, Medina A, Rutter WJ, Ammerer G, Hall BD. Synthesis and assembly of hepatitis B virus surface antigen particles in yeast. Nature. 1982;298:347-50. [ Links ]
13. Campos RH, Mbayed VA, Pineiro Y, Leone FG. Molecular epidemiology of hepatitis B virus in Latin America. J Clin Virol. 2005;34(Suppl.2):S8-13. [ Links ]
14. Kay A, Zoulim F. Hepatitis B virus genetic variability and evolution. Virus Research. 2007;127:164-76. [ Links ]
15. Zeng GB, Wen SJ, Wang ZH, Yan L, Sun J, Hou JL. A novel hepatitis B virus genotyping system by using restriction fragment length polymorphism patterns of S gene amplicons. World J Gastroenterol. 2004;10:3132-36. [ Links ]
16. Okamoto H, Imai M, Kametani M, Nakamura T, Mayumi M. Genomic heterogeneity of hepatitis B virus in a 54-year-old woman who contracted the infection through materno-fetal transmission. Jpn J Exp Med. 1987;57:231-6. [ Links ]
17. Norder H, Couroucé AM, Magnius LO. Molecular basis of hepatitis B virus serotype variations within the four major subtypes. J Gen Virol. 1992;73:3141-5. [ Links ]
18. Norder H, Couroucé AM, Coursaget P, Echevarria JM, Lee SD, Mushahwar IK, Robertson BH, Locarnini S, Magnius LO. Genetic diversity of hepatitis B virus strains derived worldwide: genotypes, subgenotypes, and HBsAg subtypes. Intervirology. 2004;47:289-309. [ Links ]
19. Kramvis A, Weitzmann L, Owiredu WK, Kew MC. Analysis of the complete genome of subgroup A hepatitis B virus isolates from South Africa. J Gen Virol. 2002;83:835-9. [ Links ]
20. Kramvis A, Kew MC. Epidemiology of hepatitis B virus in Africa, its genotypes and clinical associations of genotypes. Hepatol Res. 2007;37(Suppl.1):S9-19. [ Links ]
21. Sugauchi F, Orito E, Ichida T, Kato H, Sakugawa H, Kakumu S, Ishida T, Chutaputti A, Lai CL, Ueda R, Miyakawa Y, Mizokami M. Hepatitis B virus of genotype B with or without recombination with genotype C over the precore region plus the core gene. J Virol. 2002;76:5985-92. [ Links ]
22. Sugauchi F, Kumada H, Sakugawa H, Komatsu M, Niitsuma H, Watanabe H, Akahane Y, Tokita H, Kato T, Tanaka Y, Orito E, Ueda R, Miyakawa Y, Mizokami M. Two subtypes of genotype B (Ba and Bj) of hepatitis B virus in Japan. Clin Infect Dis. 2004;38:1222-8. [ Links ]
23. Sugauchi F, Kumada H, Acharya SA, Shrestha SM, Gamutan MT, Khan M, Gish RG, Tanaka Y, Kato T, Orito E, Ueda R, Miyakawa Y, Mizokami M. Epidemiological and sequence differences between two subtypes (Ae and Aa) of hepatitis B virus genotype A. J Gen Virol. 2004;85:811-20. [ Links ]
24. Sugauchi F, Mizokami M, Orito E, Ohno T, Kato H, Suzuki S, Kimura Y, Ueda R, Butterworth LA, Cooksley WG. A novel variant genotype C of hepatitis B virus identified in isolates from Australian Aborigines: complete genome sequence and phylogenetic relatedness. J Gen Virol. 2001;82:883-92. [ Links ]
25. Cui C, Shi J, Hui L, Xi H, Zhuoma, Quni, Tsedan, Hu G. The dominant hepatitis B virus genotype identified in Tibet is a C/D Irbid. J Gen Virol. 2002;83:2773-7. [ Links ]
26. Olinger CM, Venard V, Njayou M, Oyefolu AO, Maïga I, Kemp AJ, Omilabu SA, le Faou A, Muller CP. Phylogenetic analysis of the precore/core gene of hepatitis B virus genotypes E and A in West Africa: new subtypes, mixed infections and recombinations. J Gen Virol. 2006;87:1163-73. [ Links ]
27. Bekondi C, Olinger CM, Boua N, Talarmin A, Muller CP, Le Faou A, Venard V. Central African Republic is part of the West-African hepatitis B virus genotype E crescent. J Clin Virol. 2007;40:31-7. [ Links ]
28. Blitz L, Pujol FH, Swenson PD, Porto L, Atencio R, Araujo M, Costa L, Monsalve DC, Torres JR, Fields HA, Lambert S, Van Geyt C, Norder H, Magnius LO, Echevarría JM, Stuyver L. Antigenic diversity of hepatitis B virus strains of genotype F in Amerindians and other population groups from Venezuela. J Clin Microbiol. 1998;36:648-51. [ Links ]
29. Nakano T, Lu L, Hu X, Mizokami M, Orito E, Shapiro C, Hadler S, Robertson B. Characterization of hepatitis B virus genotypes among Yucpa Indians in Venezuela. J Gen Virol. 2001;82:359-65. [ Links ]
.30 Devesa M, Pujol FH. Hepatitis B virus genetic diversity in Latin America. Virus Res. 2007;127:177-84. [ Links ]
31. Huy TT, Ushijima H, Sata T, Abe K. Genomic characterization of HBV genotype F in Bolivia: genotype F subgenotypes correlate with geographic distribution and T(1858) variant. Arch Virol. 2006;151:589-97. [ Links ]
32. Arauz-Ruiz P, Norder H, Robertson BH, Magnius LO. Genotype H: a new Amerindian genotype of hepatitis B virus revealed in Central America. J Gen Virol. 2002;83:2059-73. [ Links ]
33. Alvarado-Esquivel C, Sablon E, Conde-González CJ, Juárez-Figueroa L, Ruiz-Maya L, Aguilar-Benavides S. Molecular analysis of hepatitis B virus isolates in Mexico: predominant circulation of hepatitis B virus genotype H. World J Gastroenterol. 2006;12:6540-5. [ Links ]
34. Kumagai I, Abe K, Oikawa T, Sato A, Sato S, Endo R, Takikawa Y, Suzuki K, Masuda T, Sainokami S, Endo K, Takahashi M, Okamoto H. A male patient with severe acute hepatitis who was domestically infected with a genotype H hepatitis B virus in Iwate, Japan. J Gastroenterol. 2007;42:168-75. [ Links ]
35. Sitnik R, Sette H Jr, Santana RA, Menezes LC, Graça CH, Dastoli GT, Silbert S, Pinho JR. Hepatitis B virus genotype E detected in Brazil in an African patient who is a frequent traveler. Braz J Med Biol Res. 2007;40:1689-92. [ Links ]
36. Devesa M, Loureiro CL, Rivas Y, Monsalve F, Cardona N, Duarte MC, Poblete F, Gutierrez MF, Botto C, Pujol FH. Subgenotype diversity of hepatitis B virus American genotype F in Amerindians from Venezuela and the general population of Colombia. J Med Virol. 2008;80:20-6. [ Links ]
37. Norder H, Couroucé AM, Magnius LO. Complete genomes, phylogenetic relatedness, and structural proteins of six strains of the hepatitis B virus, four of which represent two new genotypes. Virology. 1994;98:489-503. [ Links ]
38. Henry Lik-Yuen Chan, Joseph Jao-Yiu Sung. Hepatocellular Carcinoma and Hepatitis B Virus. Jpn J Exp Med. 2006;26:153-61. [ Links ]
39. Baumert TF, Thimme R, von Weizsäcker F. Pathogenesis of hepatitis B virus infection. World J Gastroenterol. 2007;13:82-90. [ Links ]
40. Wai CT, Fontana RJ. Clinical significance of hepatitis B virus genotypes, variants, and mutants. Clin Liver Dis. 2004;8:321-52. [ Links ]
41. Duong TN, Horiike N, Michitaka K, Yan C, Mizokami M, Tanaka Y, Jyoko K, Yamamoto K, Miyaoka H, Yamashita Y, Ohno N, Onji M. Comparison of genotypes C and D of the hepatitis B virus in Japan: a clinical and molecular biological study. J Med Virol. 2004;72:551-7. [ Links ]
42. Tsubota A, Arase Y, Ren F, Tanaka H, Ikeda K, Kumada H. Genotype may correlate with liver carcinogenesis and tumor characteristics in cirrhotic patients infected with hepatitis B virus subtype adw. J Med Virol. 2001;65:257-65. [ Links ]
43. Yu MW, Yeh SH, Chen PJ, Liaw YF, Lin CL, Liu CJ, Shih WL, Kao JH, Chen DS, Chen CJ. Hepatitis B virus genotype and DNA level and hepatocellular carcinoma: a prospective study in men. J Natl Cancer Inst. 2005;97:265-72. [ Links ]
44. Sánchez-Tapias JM, Costa J, Mas A, Bruguera M, Rodés J. Influence of hepatitis B virus genotype on the long-term outcome of chronic hepatitis B in western patients. Gastroenterology. 2002;123:1848-56. [ Links ]
45. Wang H, Seow A, Lee HP. Trends in cancer incidence among Singapore Malays: a low-risk population. Ann Acad Med Singapore. 2004;33:57-62. [ Links ]
46. Hui P, Nakayama T, Morita A, Sato N, Hishiki M, Saito K, Yoshikawa Y, Tamura M, Sato I, Takahashi T, Soma M, Izumi Y, Ozawa Y, Cheng Z. Common single nucleotide polymorphisms in Japanese patients with essential hypertension: aldehyde dehydrogenase 2 gene as a risk factor independent of alcohol consumption. Hypertens Res. 2007;30:585-92. [ Links ]
47. Milich D, Jones JE, Hughes JL, Price J, Raney AK, McLachlan A. Is a function of the secreted hepatitis B e antigen to induce immunologic tolerance in utero? Proc Natl Acad Sci USA 1990;87:6599-603. [ Links ]
48. Milich D, Liang TJ. Exploring the biological basis of hepatitis B e antigen in hepatitis B virus infection. Hepatology. 2003;38:1075-86. [ Links ]
49. Pongpipat D, Suvatte V, Assateerawatts A. Perinatal transmission of hepatitis B virus in Thailand. Asian Pac J Allergy Immunol. 1985;3:191-3. [ Links ]
50. Chia-Ming Chu, Yun-Fan Liaw. Genotype C hepatitis B virus infection is associated with a higher risk of reactivation of hepatitis B and progression to cirrhosis than genotype B: A longitudinal study of hepatitis B e antigen-positive patients with normal aminotransferase levels at baseline J Hepat. 2005;43:411-7. [ Links ]
51. Livingston SE, Simonetti JP, Bulkow LR, Homan CE, Snowball MM, Cagle HH, Negus SE, McMahon BJ. Prevalence of hepatitis B e antigen in patients with chronic hepatitis B and genotypes A, B, C, D, and F. Gastroenterology. 2007;133:1452-7. [ Links ]
52. Marcellin P, Asselah T, Boyer N. Treatment of chronic hepatitis B. J Viral Hepatitis. 2005;12:333-45. [ Links ]
53. Pardo M, Bartolomé J, Carreño V. Current therapy of chronic hepatitis B. Arch Med Res. 2007;38:661-77. [ Links ]
54. Thomas H, Fostre G, Platis D. Mechanism of interferon and nucleoside analogues. J Hepatology. 2003;39:93-8. [ Links ]
55. Ferreira MS, Borges AS. Advances in the treatment of hepatitis B. Rev Soc Bras Med Trop. 2007;40:451-62. [ Links ]
56. Pardo M , Bartolomé J, Carreño V. Current therapy of chronic hepatitis B. Arch Med Res. 2007;38:661-77. [ Links ]
57. Liu CJ, Kao JH, Chen DS. Therapeutic implications of hepatitis B virus genotypes. Liver Int. 2005;25:1097-107. [ Links ]
58. Erhardt A, Blondin D, Hauck K, Sagir A, Kohnle T, Heintges T, Häussinger D. Response to interferon alfa is hepatitis B virus genotype dependent: genotype A is more sensitive to interferon than genotype D. Gut. 2005;54:1009-13. [ Links ]
59. Kao JH, Wu NH, Chen PJ, Lai MY, Chen DS. Hepatitis B genotypes and the response to interferon therapy J Hepatol. 2000;33:998-1002. [ Links ]
60. Chien RN, Yeh CT, Tsai SL, Chu CM, Liaw YF. Determinants for sustained HBeAg response to lamivudine therapy. Hepatology. 2003;38:1267-73. [ Links ]
61. Miller RH, Robinson WS. Common evolutionary origin of hepatitis B virus and retroviruses. Proc Natl Acad Sci USA. 1986;83:2531-5. [ Links ]
62. Park SG, Kim Y, Park E, Ryu HM, Jung G. Fidelity of hepatitis B virus polymerase. Eur J Biochem. 2003;270:2929-36. [ Links ]
63. Alexopoulou A, Karayiannis P, Hadziyannis SJ, Aiba N, Thomas HC. Emergence and selection of HBV variants in an anti-HBe positive patient persistently infected with quasi-species. J Hepatol. 1997;26:748-53. [ Links ]
64. Hunt CM, McGill JM, Allen MI, Condreay LN. Clinical relevance of hepatitis B viral mutations. Hepatology. 2000;31:1037-44. [ Links ]
65. Baumert TF, Barth H, Blum HE. Genetic variants of hepatitis B virus and their clinical relevance. Minerva Gastroenterol Dietol. 2005;51:95-108. [ Links ]
66. Chan HL, Sung JJ. Hepatocellular carcinoma and hepatitis B virus. Semin Liver Dis. 2006;26:153-61. [ Links ]
67. >Buckwold VE, Xu Z, Chen M, Yen TS, Ou JH. Effects of a frequent double-nucleotide basal core promoter mutation and its putative single-nucleotide precursor mutations on hepatitis b virus gene expression and replication. J Virol 1996:5845-511. [ Links ]
68. Wang Y, Wei L, Jiang D, Cong X, Fei R, Xiao J, Wang Y. In vitro resistance to interferon of hepatitis B virus with precore mutation. World J Gastroenterol. 2005;11:649-55. [ Links ]
69. Wang Y, Wei L, Jiang D, Cong X, Fei R, Chen H, Xiao J, Wang Y. In vitro resistance to interferon-alpha of hepatitis B virus with basic core promoter double mutation. Antiviral Res. 2007;75:139-45. [ Links ]
70. Chen RY, Bowden S, Desmond PV, Dean J, Locarnini SA. Effects of interferon alpha therapy on the catalytic domains of the polymerase gene and basal core promoter, precore and core regions of hepatitis B virus. J Gastroenterol Hepatol. 2003;18:630-7. [ Links ]
71. Tacke F, Gehrke C, Luedde T, Heim A, Manns MP, Trautwein C. Basal core promoter and precore mutations in the hepatitis B virus genome enhance replication efficacy of lamivudine-resistant mutants. J Virol. 2004;78:8524-35. [ Links ]
72. Cho SW, Shin YJ, Hahm KB, Jin JH, Kim YS, Kim JH, Kim HJ. Analysis of the precore and core promoter DNA sequence in liver tissues from patients with hepatocellular carcinoma. J Korean Med Sci. 1999;14:424-30. [ Links ]
73. Chen CH, Lee CM, Hung CH, Hu TH, Wang JH, Wang JC, Lu SN, Changchien CS. Clinical significance and evolution of core promoter and precore mutations in HBeAg-positive patients with HBV genotype B and C: a longitudinal study. Liver Int. 2007;27:806-15. [ Links ]
74. Liu CJ, Chen BF, Chen PJ, Lai MY, Huang WL, Kao JH, Chen DS. Role of hepatitis B viral load and basal core promoter mutation in hepatocellular carcinoma in hepatitis B carriers. J Infect Dis. 2006;193:1258-65. [ Links ]
75. Liu CJ, Chen BF, Chen PJ, Lai MY, Huang WL, Kao JH, Chen DS. Role of hepatitis B virus precore/core promoter mutations and serum viral load on noncirrhotic hepatocellular carcinoma: a case-control study. J Infect Dis. 2006;194:594-99. [ Links ]
76. Tong MJ, Blatt LM, Kao JH, Cheng JT, Corey WG. Precore/ basal core promoter mutants and hepatitis B viral DNA levels as predictors for liver deaths and hepatocellular carcinoma. World J Gastroenterol. 2006;12:6620-6. [ Links ]
77. Jang JW, Lee YC, Kim MS, Lee SY, Bae SH, Choi JY, Yoon SK. A 13-year longitudinal study of the impact of double mutations in the core promoter region of hepatitis B virus on HBeAg seroconversion and disease progression in patients with genotype C chronic active hepatitis. J Viral Hepat. 2007;14:169-75. [ Links ]
78. Lee YI, Hur GM, Suh DJ, Kim SH. Novel pre-C/C gene mutants of hepatitis B virus in chronic active hepatitis: naturally occurring escape mutants. J Gen Virol. 1996;77:1129-38. [ Links ]
79. Liu Z, Luo K, He H, and Hou J. Hot-spot mutations in hepatitis B virus core gene: eliciting or evading immune clearance? J Viral Herat. 2005;12:146-53. [ Links ]
80. Hosono S, Tai PC, Wang W, Ambrose M, Hwang DG, Yuan TT, Peng BH, Yang CS, Lee CS, Shih C. Core antigen mutations of human hepatitis B virus in hepatomas accumulate in MHC class II-restricted T cell epitopes. Virology. 1995;212:151-62. [ Links ]
81. Ferrari C, Bertoletti A, Penna A, Cavalli A, Valli A, Missale G, Pilli M, Fowler P, Giuberti T, Chisari FV, et al. Identification of immunodominant T cell epitopes of the hepatitis B virus nucleocapsid antigen. J Clin Invest. 1991;88:214-22. [ Links ]
82. Penna A, Bertoleti A, Caravali A, Valli A, Missale G, Pilli M, Marchelli S, Giuberti T, Fowler P, Chisari F, Fisccadori F, Ferrari C. Fine specificity of the T cell response to hepatitis B virus core antigen. Arch Virol, 1992;4:23-8. [ Links ]
83. Missale G, Redeker A, Person J, Fowler P, Guilhot S, Schlicht HJ, Ferrari C, Chisari FV. HLA-A31- and HLA-Aw68-restricted cytotoxic T cell responses to a single hepatitis B virus nucleocapsid epitope during acute viral hepatitis. J Exp Med. 1993;177:751-62. [ Links ]
84. Li JS, Tong SP, Wen YM, Vitvitski L, Zhang Q, Trépo C. Hepatitis B virus genotype A rarely circulates as an HBe-minus mutant: possible contribution of a single nucleotide in the precore region. J Virol. 1993;67:5402-10. [ Links ]
85. Rodriguez-Frias F, Buti M, Jardi R, Cotrina M, Viladomiu L, Esteban R, Guardia J. Hepatitis B virus infection: precore mutants and its relation to viral genotypes and core mutations. Hepatology. 1995;22:1641-7. [ Links ]
86. Arauz-Ruiz P, Norder H, Visoná KA, Magnius LO. Molecular epidemiology of hepatitis B virus in Central America reflected in the genetic variability of the small S gene. J Infect Dis. 1997;176:851-8. [ Links ]
87. França PH, González JE, Munné MS, Brandão LH, Gouvea VS, Sablon E, Vanderborght BO. Strong association between genotype F and hepatitis B virus (HBV) e antigen-negative variants among HBV-infected Argentinean blood donors. J Clin Microbiol. 2004;42:5015-21. [ Links ]
88.Watanabe K, Takahashi T, Takahashi S, Okoshi S, Ichida T, Aoyagi Y. Comparative study of genotype B and C hepatitis B virus-induced chronic hepatitis in relation to the basic core promoter and precore mutations. J Gastroenterol Hepatol. 2005;20:441-9. [ Links ]
89. Huang YH, Wu JC, Chang TT, Sheen IJ, Huo TI, Lee PC, Su CW, Lee SD. Association of core promoter/precore mutations and viral load in e antigen-negative chronic hepatitis B patients. J Viral Herat. 2006;13:336-42 [ Links ]

