











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Shiga toxin-producing Escherichia coli (STEC) or verocytotoxin (VT)-producing E. coli (VTEC) strains are pathogens of public health concern worldwide, and associated with disease in humans and animals. In humans, they can cause severe outbreaks of gastrointestinal illness with clinical symptoms ranging from diarrhea and hemorrhagic colitis to the life-threatening haemolytic uremic syndrome1. STEC strains causing human infections belong to a large number of O:H serotypes. Most outbreaks and sporadic cases of hemorrhagic colitis and haemolytic uremic syndrome (HUS) have been attributed to the STEC O157 strains; the H7 flagellum is frequently but not always present on O157 isolates 2. However, as STEC non-O157 strains are more prevalent in animals and as contaminants in foods, humans are probably more exposed to these strains. Infections with some non-O157 STEC types, such as O26:H11 or H-, O91:H21 or H-, O103:H2, O111:H-, O113:H21, O117:H7,O118:H16, O121:H19, O128:H2 or H-, O145:H28 or H- and O146:H21 are frequently associated with severe illness in humans, but the role of other non-O157 STEC types in human disease needs further examination3.
STEC strains isolated from a variety of animals and cattle are considered the main reservoir4. Nevertheless, recent studies have indicated that small domestic ruminants, including sheep and goats, are also key reservoirs of STEC 5,6. In particular, sheep and their products have been documented as reservoirs for STECs that belong to a diverse set of non-O157 serogroups (O26, O91, O115,O128, and O130) and possess genes encoding key virulence factors that have been implicated in human disease 7. Human infections commonly derived from beef products, un-pasteurized milk and contaminated water, during the processing of the carcasses, fecal contamination and/or transfer of bacteria from the animal's hide to the carcass can facilitate transmission of pathogenic E. coli to the meat 8.
Gulf states, especially Saudi Arabia is a major sheep importer of meat. It represents the world's largest import market for live sheep to meet the strong consumer demand, as sheep meat forms an important component of the Arab diet9.
Shiga toxin-producing E. coli are characterized by the production of one or more types of Shiga toxin (stx1 or stx2 or their variants), which interfere with the protein synthesis of host cells, leading to cell death. These toxins are synonymously either called verocytotoxins because of their activity on Vero cells or Shiga toxins because of their similarity with the toxin produced by Shigella dysenteriae. In addition to toxin production, another virulence-associated factor expressed by STEC is a protein called intimin, which is responsible for intimate attachment of STEC to intestinal epithelial cells, causing attaching and effacing lesions in the intestinal mucosa. Intimin is encoded by the chromosomal gene eae, which is part of a pathogenicity island termed the locus for enterocyte effacement. Severe diarrhea (especially hemorrhagic colitis) and hemolytic uremic syndrome were closely associated with STEC types carrying the eae gene for intimin 10. Additionally, enterohemolysin, expressed by the ehxA gene, liberates hemoglobin from the red blood cells during infection and has been linked to severe disease 11.
The aim of the present study was to identify the serotypes and virulence genes of STEC isolates recovered from human and sheep fecal materials to ascertain if ovine STEC strains possess the same serotypes and virulence profiles as STEC strains that cause human infections. Chromosomal DNAs of O157:H7 isolates were analyzed by pulsed-field gel electrophoresis (PFGE) to determine the isolates potential as human pathogen and the genetic relatedness to obtain a better understanding of the relevant sources of zoonotic O157:H7 isolates. Susceptibility of the isolates to eleven commonly used antibiotics was also investigated.
Materials and methods
Sampling
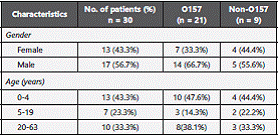
Stool samples were collected from patients with diarrhea from Ouhud Hospital at Al-Madinah Al-Munawarah, demographic data of human patients were depicted in Table 1. Fecal samples were collected from randomly selected sheep at the central sheep market in Al-Madinah Al-Munawarah between June and August 2015. A total of 221 samples; 134 human samples and 87 sheep samples, were collected into appropriate capped tubes. Samples were transferred on ice to Taibah University microbiology laboratory, biology department for immediate processing.
Isolation and identification of STEC
Isolation of E. COLI O157:H7: approximately 1 g of each fecal sample was mixed in 9 ml of Trypticase soya broth (TSB) with 20 mg/L novobiocine and incubated for 6-8 h at 41.5oC for 6 h then transferred and plated onto sorbitol MacConkey agar (SMAC) supplemented with 1 mg/L potassium tellurite and incubated for 18-24 h at 37oC. A pale colony each (sorbitol nonfermenters) was picked as presumptive E. coli O157 per sample and were confirmed to be E. coli biochemically by the indole, methyl red, Voges-Proskauer, and citrate tests (IMViC)12.
Isolation of non-O157 STEC strains: fecal sample were cultured on SMAC agar (Difco) by the streak plate technique. After overnight incubation of the plates at 37oC, colonies were confirmed by the indole, methyl red, Voges-Proskauer, and citrate tests (IMViC).
Somatic and flagellar serotyping
Isolates were grown on nutrient agar plates at 37oC overnight, then the cells were collected and suspended in 0.9% sterile normal saline (154 mEq/L sodium and 154 mEq/L chloride, pH adjusted to 7) then autoclaved at 121oC for 15 min. The cells were concentrated by centrifugation and suspended in an appropriate volume of sterile normal saline. Detection of O-serogroup was performed using a commercially available O-serogrouping Kit13. The Flagellar phase inversion was carried out using the standard Craigie tube technique by passage through semi-solid agar containing the appropriate flagellar antisera14.
Antimicrobial susceptibility testing
The antimicrobial susceptibility phenotypes of Shiga toxinproducing E. coli (STEC) strains from human patients and sheep was performed following the standard disc diffusion method on Muller-Hinton agar plates, using commercially available antimicrobial susceptibility discs (Kirby-Bauer SN DISC) according to the standards and interpretive criteria described by CLSI15. The following antibiotics were used ampicillin (10 μ/disk), oxytetracyclin (30 μ/disk), gentamycin (30 μ/disk), ciprofloxacin (5 μ/disk), cefotaxime (30 μ/disk), streptomycin (15 μ/disk), norofloxacin (30 μ/disk), polymyxin (300 μ/disk), chloromphenicol (30 μ/disk), Kanamycin (30 μ/disk), and trimetoprime-sulfamethoxazole (23.75 μ/disk). The results were recorded on the basis of the zone-size interpretative chart supplied by the manufacturer.
Template DNA preparation
DNA templates for PCR were obtained from overnight E. coli cultures that were harvested, then suspended in 200 ml of sterile distilled water, and boiled for 15 min 16.
Detection of virulence genes
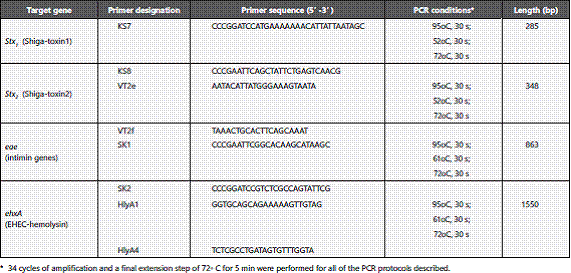
Detection of virulence genes was performed by PCR. Primer sequences and PCR conditions used for the study were listed in Table 2 17. PCR performed in the Takara thermal cycler (Bio-Rad, USA). PCR products were separated and visualized by gel electrophoresis in 1.5% agarose (Wako) in Tris-acetate- EDTA (TAE) buffer at 100 V, where a 100 or 500 bp DNA ladder (one-step ladder, Wako) was included in each agarose run, accordingly the amplified product.
Pulsed field gel electrophoresis (PFGE)
PFGE was performed according to CDC (the PulseNet protocol of the Centers for Disease Control and Prevention). The agarose-embedded bacterial genomic DNA was digested with restriction enzyme XbaI at 37oC for 4 h. Electrophoresis was performed on 1% agarose gel with 0.5 Tris-borate-EDTA buffer. The electrophoretic conditions were optimized for the separation of the 24- to 600-kB XbaI-digested macrore-striction fragments. The following PFGE parameters were applied: voltage of 6 v/cm, initial switch time of 2.2 s, final switch time of 54.2 s, and run time of 19 h. Electrophoresis was conducted by using a CHEF-DRII (Bio-Rad Laboratories, Tokyo, Japan). The gel was stained with ethidium bromide and imaged with the Gel Doc 2000 and Multi-Analyst program (Bio-Rad). Dendrograms were created with a Molecular Analyst (Bio-Rad) using the Dice coefficient, unweighted pair group method with arithmetic means (UPGMA) and a position tolerance of 1.3%.
Results
Shiga toxin-producing E. coli O157 strains
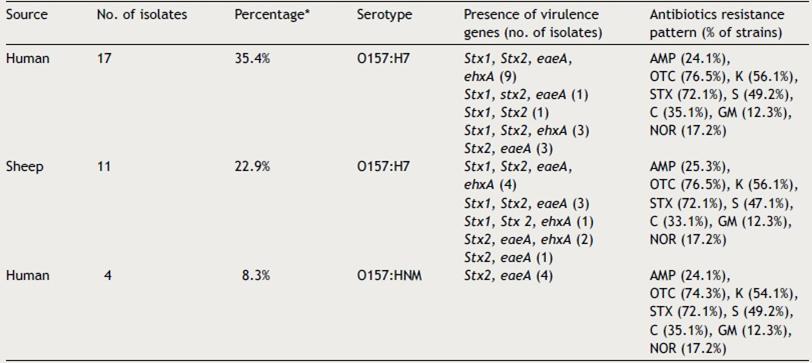
A total of 28 isolates (17 human isolates and 11 sheep isolates) showed typical colony phenotype (sorbitol negative), had IMViC reactions characteristic of E. coli (++ -- ), agglutinated with O157 and H7 antisera and possessed species specific gene (uidA) 18. Four O157:HNM isolates were recovered from humans (Table 3).
Shigatoxin-producing E. coli non-O157 strains
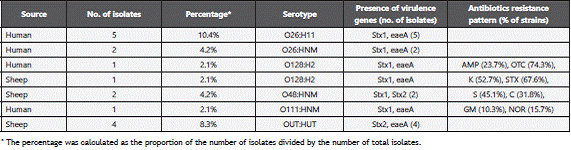
A variety of non-O157 Shiga toxin-producing E. coli reported in the present study belong to 4 (O) serogroups and 6 (O: H) serotypes (Table 4). The most prevalent serotype among human patients was O26:H11 (5/48; 10.4%) and O26: HNM (2/48; 4.2%). Two isolates (2/48; 4.2%) belong to serotype O128: H2; one isolated from human and the other from sheep. Serotype O48: HNM was reported only among sheep isolates (2/48; 4.2%). Single strain (1/48; 2.1%) of serotype O111: HNM was recovered from human patient, while four (4/48; 8.4%) untypable strains were isolated from sheep.
Antibiotics susceptibility of Shiga-toxin E. coli isolates
The phenotypic testing of antibiotics susceptibility of the 48 Shiga toxin-producing E. coli (STEC) isolates, showed high prevalence of multi-resistance to various antimicrobial agents. Human STEC O157:H7 strains exhibited resistance to oxytetracyclin, trimethoprime-sulphamethoxazole, kanamycin, streptomycin, chloramphenicol, ampicillin, norofloxacin and gentamycin (76.5%, 72.1%, 56.1%, 49.2%, 35.1%, 24.1%, 17.2% and 12.3% respectively), while sheep strains of O157:H7 showed the same resistance pattern as human strains except for the resistance to streptomycin, chloramphenicol and ampicillin were (47.1%, 33.1% and 25.3% respectively) (Table 3). STEC non-O157 strains indicated slightly lower resistance to oxytetracyclin, trimethoprim-sulphamethoxazole, kanamycin, streptomycin, chloramphenicol, ampicillin, norofloxacin and gentamycin (74.3%, 67.6%, 52.7%, 45.1%, 31.8%, 23.7%,15.7% and 10.3% respectively) (Table 4). However, all isolates showed 100% susceptibility to polymyxin B, ciprofloxacin and cefotaxime.
Virulence genes distribution among different STECs serotypes
Forty eight confirmed E. coli isolates were analyzed by PCR for the presence of stx1, stx2, eaeA, ehxA; the characteristic virulence genes of STECs. The distribution of the virulence genes showed a variety of virulence profiles among STEC O157 (Table 3) and non-O157 isolates (Table 4). The predominant genotype among the tested E. coli O157:H7 isolates was stx1 , stx2 , ehxA and eae, accounting for 13 of the 28 (46.4%). Of the remaining isolates, four (14.3%) were stx2 and eaeA; four (14.3%) were stx1, stx2 and ehxA; four (14.3%) were stx1, stx2 and eaeA; two (7.1%) were stx1, eaeA, ehxA and one (3.6%) was stx1, stx2 and. Four human O157:HNM isolates possessed stx2 and eaeA genotype. Among the 16 non-O157 STEC isolates, ten (62.5%) were stx1 and eaeA; four (25%) were stx2 and eaeA and two (12.5%%) were stx1 and stx2 (Table 4).
PFGE profiles of E. coli O157:H7 isolates
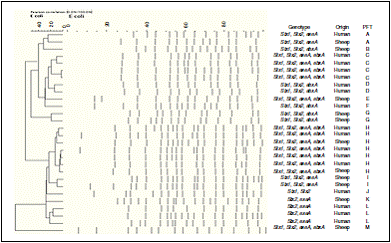
Thirteen different PFGE patterns named A through M, were found among the 28 E. coli O157:H7 isolates (15 human isolates, 13 sheep isolates). As indicated in Fig. 1, the predominant PFGE pattern seen from the human and ovine E. coli O157:H7 isolates was pattern H (25%); including both human and sheep isolates with more than 80% similarity, followed by pattern C (14.3%) and pattern L (10.7%). The observed PFGE pattern was significantly associated with the genotype. Isolates with PFGE patterns H, C, E, and M all had the stx 1, stx 2, ehxA and eaeA genotype. However, E. coli O157:H7 isolates with the other PFGE patterns possessed a variety of genotypes, such as stx1 , stx2 and eaeA (A and I); stx1 , stx2 and ehxA (B, D, F and G); stx1 and stx2 (J) or stx2 and eaeA (K and L).
Discussion
The present study investigated Shiga toxin-producing E. coli (STEC) in human and sheep fecal materials in Al-Madinah Al- Munawarah, Saudi Arabia; where people depend mainly on sheep meat. To our knowledge this study is the first to report the presence of O157 and non O157 STEC in human and sheep fecal materials in Saudi Arabia. A total of 221 fecal samples (134 human and 87 sheep) were collected and analyzed for the presence of STEC strains. A total of 48 STEC isolates confirmed biochemically, serologically and by PCR targeting the characteristic virulence genes stx1 , stx2 , eaeA and ehxA. The highest number of reported STECs among human patients was reported in the youngest age category (46.7%) and in males (63.3%). Higher prevalence of O157 than non-O157 Shiga toxin producing E. coli was reported among human and sheep samples especially E. coli O157:H7 serotype. The high frequency of E. coli O157:H7 serotype can be explained by the fact that the samples were collected during summer when they are known to increase.19. Sheep are a natural reservoir for E. coli O157:H7, in addition, sheep have also been cited as reservoirs for a diverse number of non-O157 Shiga toxin producing E. coli serogroups, which have been implicated in human disease 20,21. A good understanding of the primary animal reservoirs would be crucial for the development of control measures of any risk factors that could lead to human infections with zoonotic STEC, given that sheep are considered relevant reservoirs of zoonotic STEC 5,22. Lack of hygienic practices could potentially contribute to the transmission of pathogens from sheep meat to human. Previous studies had suggested that STEC prevalence in feces was correlated with the pathogen's prevalence on carcasses 23-25.
Diarrhea caused by E. coli requires antimicrobial therapy; however, antibiotic-resistant strains cause longer and more severe illnesses than their antibiotic-susceptible counterparts. Several studies have shown that antibiotic resistance in E. coli has increased over time. 26-27. Investigation of antimicrobial resistance revealed a very high level of multiple antimicrobial resistances among the isolates and the most common resistance was to tetracycline which was often used as a first-line antimicrobial and growth promoter in food animals and its widespread use has contributed to high rates of resistance 28. The frequency of tetracycline resistance among the E. coli isolates was 76.5- 74.3% which is within the range of values described in previous reports (68-93%) 29-30. STECs isolates of the present study also showed high resistance to trimethoprime-sulphaethoxazole, kanamycine, streptomycin, chloromphenicol and Ampicillin.
Knowledge of recent regional patterns of antimicrobial resistance is critical to therapeutic decision-making. To our knowledge this is the first report on STEC isolates from human and sheep in Saudi Arabia. Therefore, no previous studies were reported to compare our results, but we can compare the results with other countries. In Iraq a study on children revealed resistance of O157:H7 strains to erythromycin, polymyxin B and vancomycin and susceptibility to cephalexin, ciprofloxacin, gentamicin and nalidixic acid 31. A study on STEC isolated from ruminants, reported a high resistance (>65%) to tetracycline, streptomycin, erythromycin, and sulfamethoxazole. Moreover, a high resistance (30%) to ampicillin, chloramphenicol, trimethoprim, and trimethoprim-sulfamethoxazole and susceptibility (>90%) to gentamicin and colistin was reported.32 O157:H7 Human strains recorded a resistance to tetracycline, sulphamethoxazole and erythromycin.33 A high resistance of STEC strains to penicillin (100%), followed by tetracycline (86.88%), gentamicin (62.29%) and streptomycin (54.91%) was also reported 27. In this regard, the use of antimicrobials in food producing animals resulting in the transmission of resistant bacteria by the food vehicle.
STEC human and sheep isolates possessed various combinations of the virulence-associated genes. The most common virulence genotype among STEC O157 strains in which 40.6% of the isolates are stx1 , stx2 , eaeA, ehxA. Many reports attempted to correlate the presence of specific recognized virulence factors with disease or severity of disease and concluded that no single factor is responsible for the virulence of STEC.34,35Previous studies have shown a certain degree of homogeneity for the presence of virulence factors within STEC serotypes 36. The present study showed that stx1 was significantly more frequent in isolates from O157 and non- O157 serotypes, followed by stx2 and none of the strains lack either stx1 or stx2 . ehxA was observed to be frequent between isolates from O157 serotypes, but completely absent in non-O157 serotypes. eaeA also seems to be restricted to O157 serotypes except serotype O26:H11. Our results are in agreement with previous studies which reported that ehxA,37eae,38 and stx2 39 are found more frequently in O157 isolates from patients with severe disease than in other STEC populations. Interestingly, STEC isolates from sheep showed the same pattern of distribution of the virulence genes among different serotypes in parallel with human isolates.
PFGE remains an effective tool for detecting variation between closely related strains. Analysis of E. coli populations by use of PFGE has shown that serotype is a good marker for the genetic background of STEC in terms of unknown virulence factors involved in the pathogenesis of STEC-associated diseases 1.This approach was followed in the present study to describe genetic links between virulence and serotype. PFGE and genotyping are complementary methods as PFGE primarily detects insertion/deletion variation within genomic regions specific to STEC O157 40 .Thus, genotyping combined with PFGE could be very useful in assessing strain diversity and evolutionary relatedness between epidemiologically unrelated strains. On the basis of combined genotype and PFGE profile, we are able to distinguish human and sheep O157:H7 isolates obtained in the present study. The isolates were revealed in two main clusters, each cluster included human and sheep isolates in a high percentage of homology. The most prevalent PFGE pattern among the isolates was (H), which including 5 human isolates and 2 sheep isolates with over 80% homology percentage indicating that the strains were almost the same. Together with almost equal distributions of virulence genes found among ovine and human isolates indicate localized transmission between sheep and humans. The observed clustering of isolates was explained by their genotypes. Together with the significant associations between human and sheep isolates, provide further evidence of an animal/environmental-associated pathway of sporadic STEC infection.
The present study supports the concept that sheep are a natural reservoir for potentially virulent E. coli O157:H7 and non-O157 STEC strains. In addition sheep strains of E. coli O157:H7 are closely related to human isolates. Our results cannot be used to establish a direct cause-and-effect relationship between pathogen prevalence in sheep fecal materials and human pathogenicity. Further studies are needed to determine the relevance of these implications.
Ethical responsibilities
Protection of human and animal subjects: The authors declare that the procedures followed were in accordance with the regulations of the responsible Clinical Research Ethics Committee and in accordance with those of the World Medical Association and the Helsinki Declaration
Confidentiality of data: The authors declare that they have followed the protocols of their work center on the publication of patient data
Right to privacy and informed consent: The authors must have obtained the informed consent of the patients and/or subjects mentioned in the article. The author for correspondence must be in possession of this document

