










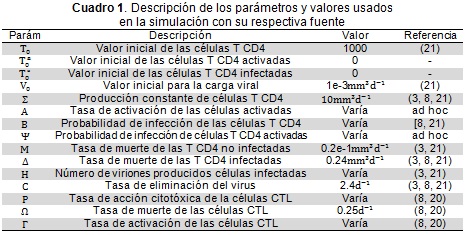
Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en
SciELO
Similares en
SciELO -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkRevista de Salud Pública
versión impresa ISSN 0124-0064
Rev. salud pública vol.16 no.1 Bogotá ene./feb. 2014
https://doi.org/10.15446/rsap.v16n1.37530
http://dx.doi.org/10.15446/rsap.v16n1.37530
Artículo/Investigación
Modelo de simulación para la infección por VIH y su interacción con la respuesta inmune citotóxica
A simulation model for HIV infection and its interaction with a cytotoxicimmune response
César A. Londoño-González,Hernán D. Toro-Zapata y Carlos A. Trujillo-Salazar
Universidad del Quindío. Armenia, Colombia. cesarlond@gmail.com; hdtoro@uniquindio.edu.co; catrujillo@uniquindio.edu.co
Recibido 19 Marzo 2013/Enviado para Modificación 10 Julio 2013/Aceptado 16 Agosto 2013
RESUMEN
Con base en ecuaciones diferenciales no lineales se formula un modelo matemático que describe la dinámica de interacción del VIH con células T CD4, que se considera que pueden ser activadas o no activadas en el reconocimiento de partículas virales; en cualquiera de los dos casos, son susceptibles a la infección con el virus. Se encuentran los puntos de equilibrio del sistema y en el caso particular del equilibrio trivial o de ausencia de infección, se determina su estabilidad local con base en el número básico de reproducción. Se efectúa además la simulación numérica del modelo para establecer el comportamiento que presentan las células infectadas y la carga viral, frente a variaciones de algunos de los parámetros. Finalmente, se reformula el modelo considerando respuesta inmune celular de tipo citotóxico y se realiza la simulación numérica.
Palabras Clave: VIH, sistema inmunológico, dinámica de población, linfocitos t (fuente: DeCS, BIREME).
ABSTRACT
Nonlinear differential equations were used for formulating a mathematical model describing the dynamics of HIV interaction with CD4 T-cells which are considered to be activated or not activated during recognition of viral particles; in either case they are susceptible to HIV infection. The system's equilibrium points were found and local stability was determined for trivial equilibrium or the absence of infection based on the basic reproduction number. The model was used for numerical simulation to show infected cell and viral load patterns regarding the variations of some parameters. The model was then reformulated, considering a cytotoxic cellular immune response and numerical simulation was run again.
Key Words: HIV, immune system, population dynamics, T-cell, mathematical model (source: MeSH, NLM).
La infección por VIH se transmite principalmente por relaciones sexuales sin medidas preventivas,entre otras vías de transmisión. Una vez el virus ingresa a las paredes mucosas empieza el ciclo infeccioso. El virus tiende a atacar las células que expresan el receptor CD4 y la interacción se da a través de los correceptores y se fundamenta en el tipo de tropismo del VIH: T-trópico (X4) que infecta a linfocitos T CD4, M-trópico (R5) que infecta a macrófagos y el dual-trópico (R5/X4) que posee la habilidad de infectar tanto a macrófagos como a linfocitos T CD4 (1-3).
Las dendríticas (presentadoras de antígenos) hacen parte de la primera línea de defensa y se encuentran tanto en las membranas mucosas como en el flujo sanguíneo, actúan como centinela del sistema inmune. Las células dendríticas presenta proteínas como las lectinas DC-SIGN y L-SING a las que se adhieren con alta afinidad numerosos virus; por lo tanto,estás células llevanproteínas virales (y viriones) en la superficie de su membranay las presentan a los linfocitos T CD4, lo que dauna señal de alarma de la presencia del VIH invasor. Las interacciones entre las células dendríticas y los linfocitos T CD4 se lleva a cabo por medio de las proteínas del complejo mayor de histocompatibilidad (CMH) de clase II, presentes en las membranas de las células dendríticas, las cuales se enlazan fuertemente al complejo TcR/(CD3 o CD4) de los linfocitos T,con la finalidad de estimular suactivación y proliferación clonal,lo que los lleva a su especialización en el reconocimiento de proteínas virales. Las células T ya activadas, se encargan de activar las células T efectoras de acción citotóxica(células T CD8) y las T de memoria (linfocitos B); estas últimasfavorecen el marcaje de las células infectadaspor el VIH para que lascélulas citotóxicas (T CD8) hagan el reconocimiento y realicen el proceso de eliminación de las células infectadas (1,3). El empalme de los viriones con las células T y células T CD4 especializadas es posible por la glicoproteína gp120 ubicada en la envoltura del virus y las moléculas CD4 en la superficie de las células T.El empalme produce cambios conformacionales en la gp120 y facilita la unión con los receptores CXCR4 o CCR5, lo cual le permite al VIH fusionarse con la membrana plasmática de la célula del huésped e iniciar el proceso de infección de la célula. Una vez en el interior de la célula, el virión se desensambla y utiliza sus herramientas proteicas (principalmente integrasa, proteasa y transcriptasa inversa) para el control total de la célula y el ensamble de nuevos viriones. En los linfocitos infectados, la replicación masiva ocasiona la destrucción de las células debido al ensamblaje y salida del virus generando lesiones irreparables a la membrana celular (1).
Cada día se generan entre 109 y 1010 partículas virales con una vida media de 6horas, las cuales son responsables de continuar el ciclo de infección a nuevos linfocitos T,lo que lleva a una reducción progresiva de la subpoblación de células T. El conteo de linfocitos T, la situación clínica, y la carga viral plasmática (CVP) constituyen los elementos básicos para establecer las decisiones terapéuticas en las distintas situaciones clínicas y para el tipo de terapia antirretroviral (TAR) a emplearse.De acuerdo con lo anterior, la relación entre el virus, las células T CD4 activas y la respuesta inmune celular, parece ser determinante en la evolución futura de la infección, debido a que si la concentración de células T CD4 activas es baja, la respuesta inmune celular no será desplegada efectivamente y el cuerpo no contará con las herramientas necesarias para controlar la infección, lo que llevaría al uso temprano de medicamentos antirretrovirales; por lo tanto, el objetivo de este artículo es estudiarmatemáticamente el efecto de la activación y especialización de las células T en la dinámica de infección del VIH y su relación con la respuesta inmune citotóxica. La bibliografía alrededor del modelado matemático de la transmisión del VIH y del SIDA es amplia, pero algunos modelos planteados en la literatura se han enfocado en el modelado de la infección y el tratamiento se citan algunos trabajos interesantes (4-15).
Análisis del modelo
- Formulación
El modelo se propone de manera que incluye el proceso de activación de las células T CD4 y su intervención en la dinámica de infección por VIH. Sea T=T(t) el número promedio de células T CD4 en un tiempo t, Tα=Tα(t) el número promedio de células T CD4 activadas y especializadas en identificar proteínas virales, T*=T*(t) el número promedio de células T CD4 infectadas en un tiempo t y V=V(t) la concentración promedio de VIH en un tiempo t. Asuma que las células se incrementan a una tasa constante σ y que mueren en proporción a su tamaño con tasa de muerte μ, por lo tanto el número promedio de células que mueren en un tiempo t está dado por μT. De manera semejante, las células T CD4 especializadas se activan a un promedio αTV, término que describe el encuentro de las células T con las partículas virales infecciosas (por ejemplo, durante la presentación de antígenos, lo que lleva a la activación y especialización de la célula T). Se asume que las células T CD4 especializadas mueren también a un promedio μTα. Si la población de células T es infectada por el virus con una probabilidad β, el principio de acción de masas permite establecer que el número promedio de células que se infectan en un tiempo t es βTV, que corresponde a una consideración clásica en este tipo de modelos. Del mismo modo las células TCD4 especializadas Tα son susceptibles a la infección y se infectan a un promedio de ψTαV en un tiempo t.
Finalmente, la concentración promedio de partículas virales producidas es proporcional al promedio de células infectadas que mueren por lisis celular δT* si esta tasa de proporcionalidad es η, se tiene un total de partículas virales ηδT* nuevas por unidad de tiempo. La tasa de eliminación natural del virus c da origen a que se eliminen cV partículas virales por unidad de tiempo. En el Cuadro 1 se muestran las descripciones de los parámetros usados en el modelo,

Con condiciones iniciales T(0)=T0, Tα(0)=T0α, T*(0)=T0* y V(0)=V0, donde los parámetros son todos positivos y β, ψ, ∈ [0,1].
La región 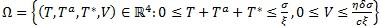 es un compacto positivamente invariante para el sistema (1 a-d). Esta región tiene importancia matemática ya que garantiza que las soluciones del sistema ingresan en la región y se mantienen en ella para todo t≥0, y además es importante desde el punto de vista inmunológico, ya que se garantiza que las poblaciones no se hagan negativas ni que crezcan indefinidamente.
es un compacto positivamente invariante para el sistema (1 a-d). Esta región tiene importancia matemática ya que garantiza que las soluciones del sistema ingresan en la región y se mantienen en ella para todo t≥0, y además es importante desde el punto de vista inmunológico, ya que se garantiza que las poblaciones no se hagan negativas ni que crezcan indefinidamente.
- Puntos de equilibrio
Los puntos de equilibrio son soluciones constantes del sistema y representan valores numéricos para cada población en los cuales el sistema se puede mantener de manera indefinida.
Proposición 1. (Puntos de equilibrio). El sistema dinámico (1 a-d) tiene dos puntos de equilibrio: 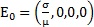 que corresponde a la ausencia de infección y
que corresponde a la ausencia de infección y 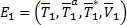 que corresponde al equilibrio en presencia de infección. Las coordenadas
que corresponde al equilibrio en presencia de infección. Las coordenadas 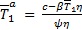 para
para 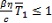 ,
, 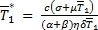 y
y 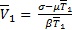 para
para 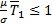 . La coordenada
. La coordenada  viene dada por,
viene dada por,
Demostración. Para determinar los puntos de equilibrio del sistema se resuelve el sistema algebraico obtenido de igualar cada ecuación a cero, de donde se obtiene la solución trivial, que corresponde a la ausencia de infección y el equilibrio no trivial, , con , y y donde es solución positiva de la ecuación,
ημ(β - ψ)(β + α)T2 + cμ(ψ + (β + α)(R1 - 1))T - ψcσ = 0 (2)
Donde  y cuyas soluciones son,
y cuyas soluciones son,
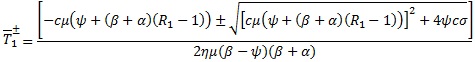
Para β > ψ es evidente que 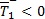 y que
y que 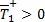 siempre. Los roles se invierten cuando β < ψ ya que en este caso
siempre. Los roles se invierten cuando β < ψ ya que en este caso 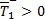 y
y 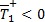 . De este modo se muestra que el sistema (1 a-d) tiene solo un equilibrio no trivial descrito por los casos i y ii de la Proposición 1. Para el caso β = ψ, el coeficiente del término cuadrático en la ecuación (2) se hace cero y queda la ecuación lineal,
. De este modo se muestra que el sistema (1 a-d) tiene solo un equilibrio no trivial descrito por los casos i y ii de la Proposición 1. Para el caso β = ψ, el coeficiente del término cuadrático en la ecuación (2) se hace cero y queda la ecuación lineal,
cμ(ψ + (β + α)(R1 - 1))T - ψcσ = 0 (2)
Cuya única solución es 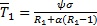 . Note que una condición suficiente para que tenga sentido inmunológico (
. Note que una condición suficiente para que tenga sentido inmunológico (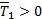 ) es R1 < 0, mientras que una condición necesaria es
) es R1 < 0, mientras que una condición necesaria es 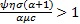 .
.
Lo interesante es conocer cuál es el comportamiento del sistema cuando no se encuentra en equilibrio, sino en valores cercanos; para analizar esa situación se determina el número básico de reproducción y con base en él la estabilidad del sistema en el equilibrio trivial.
- Número básico de reproducción
El número básico de reproducción, denotado con R0, establece el número de células secundarias que son infectadas por una sola célula infectada a partir de la carga inicial de virus. Para determinarlo, se definen los vectores,
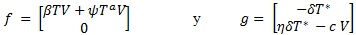
Que están formados por las ecuaciones de las poblaciones infecciosas y a partir de los cuales se determinan las matrices Jacobianas F = Dƒ y G = Dg respecto a las variables T* y V. La matriz de la siguiente generación está dada por M = -FG-1, evaluada en el equilibrio en ausencia de infecciones,
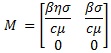
Cuyo radio espectral define el R0, de este modo, 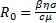 . El valor de R0 es de suma importancia, pues determina el aumento o la disminución de la producción de viriones en el sistema inmune, situación que se hace manifiesta en en el siguiente resultado.
. El valor de R0 es de suma importancia, pues determina el aumento o la disminución de la producción de viriones en el sistema inmune, situación que se hace manifiesta en en el siguiente resultado.
Proposición 2. (Estabilidad local del punto de equilibrio trivial). El equilibrio en ausencia de infección E0 es local y asintóticamente estable si y solo si R0 < 1.
Demostración. Se determina la matriz Jacobiana del sistema (1 a-d) se evalúa en el punto de equilibrio en ausencia de infección. El polinomio característico correspondiente es,
p(λ) = (μ + λ)2(λ2 + (c + δ)λ - δc(R0 - 1))
Para el cual, un valor propio evidente es λ1,2 = -μ de multiplicidad algebraica 2. Y para el factor cuadrático del polinomio se aplica el criterio de Routh-Hutwitz con n = 2. Para ello se hacen a1 = (c + δ) y a2 = -δc(R0 - 1), los cuales son positivos si y solo si R0 < 1. Bajo estas condiciones, los cuatro valores propios tienen parte real negativa con lo que se prueba la proposición.
- Simulación numérica
La simulación es hecha con los parámetros que se muestran en el Cuadro 1 y con el fin de revelar el efecto que tienen β, η y σ en la dinámica; de este modo es posible determinar cuáles de ellos son susceptibles de ser controlados.
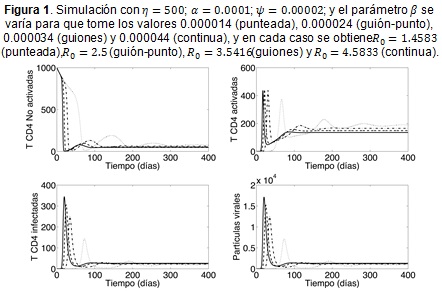
En la Figura 1 se está variando el parámetro β y allí se observa cómo responde el modelo a diferentes probabilidades de infección (virulencia). Evidentemente las probabilidades menores conllevan a brotes de infección con menor intensidad, además el brote tarda más tiempo en presentarse; sin embargo, en estos casos las oscilaciones que presentan todas las poblaciones tienen mayor frecuencia y tardan más tiempo en estabilizarse. Nótese que en todos los casos simulados, se tendría una población final de células T (activadas y sin activar) inferior a las 350 cel/mm3, lo que implica que la terapia antirretroviral es fuertemente recomendable (16).
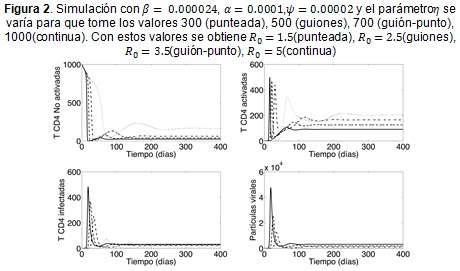
La Figura 2 muestra cómo reacciona la dinámica frente a variaciones del parámetro η. Las mayores tasas de producción implican que los niveles celulares desciendan con mayor rapidez y además se presenta el brote de infección en muy poco tiempo después del contagio. Por otro lado, las tasas de producción más bajas conllevan a un brote de menor intensidad y retrasan su ocurrencia en el tiempo, pero por otro lado, incrementan la amplitud de las oscilaciones y hacen que tome más tiempo al sistema llegar al estado de equilibrio. Note que solo con la tasa de producción más baja (η = 300), el paciente tendría un conteo de células T sanas superior a las 350 cel/mm3, en los demás casos el conteo celular es inferior, situación en la que algunos autores consideran necesaria la terapia antirretroviral (16).
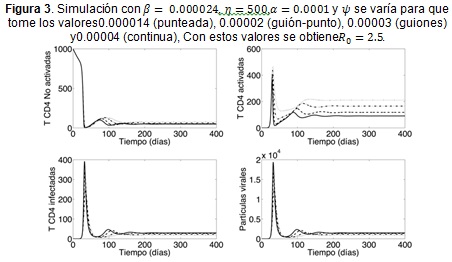
El comportamiento del modelo al variar el parámetro ψ es semejante al que se obtiene al variar a β, aunque la amplitud de las oscilaciones es menor, como puede evidenciarse en la Figura 3. En efecto, valores pequeños de la probabilidad de infección conllevan a brotes más pequeños pero con oscilaciones más amplias.
Una observación importante que se obtiene de las Figuras 1 y 3 es que los parámetros β, η y ψ, son los que tienen mayor efecto sobre la población de células infectadas y la carga viral. Por otro lado, el parámetro α es importante en el modelo que se estudia debido a que las células activadas y especializadas en el reconocimiento de partículas virales Tα son las encargadas de desplegar la respuesta inmune celular y la humoral, que son procesos que influyen directamente sobre el control de la infección. Para poner esto de manifiesto, se propone hacer una modificación al modelo (1 a-d) para incluir la respuesta inmune celular de tipo citotóxico.
Modelo con respuesta citotóxica
La respuesta inmune se clasifica en humoral y celular. La respuesta humoral específica se enfoca en la producción de anticuerpos, mientras que en la respuesta celular específica se distingue la acción de los linfocitos T CD4 activados y se pone de manifiesto por una intensa expansión clonal de linfocitos T CD8 con actividad citotóxica (CTL). La respuesta de estas células aparece temprano en la evolución de la infección y puede constituir un mecanismo de depuración de la viremia plasmática inicial. Aunque la principal célula que media la actividad citotóxica es la célula CD8, también se ha inducido dicha actividad en CD4. Debido a que el VIH no ataca directamente las células CTL, se ha reportado clínicamente que los individuos que mantienen niveles altos de éstas células se mantienen sanos por más tiempo. El objetivo es elevar los niveles de células CTL en el individuo con el fin de mantener niveles altos de células T y bajas cargas virales.
Tras el contacto con el VIH se produce la primoinfección; esta se caracteriza por un período ventana de 4 a 12 semanas durante el cual no es posible detectar anticuerpos específicos con pruebas serológicas, aunque existe una viremia muy alta. Durante este período es posible detectar actividad citotóxica frente al VIH mediante la caracterización clonal de los linfocitos CD8 del paciente.La actividad celular antiviral en ausencia de anticuerpos sugiere, que la respuesta celular es más precoz e importante en el control inicial de la replicación viral que la respuesta humoral (1,8,17,18).
Teniendo en cuenta lo anterior, sea Tc = Tc(t) el número promedio de células de respuesta inmune con efecto citotóxico debido a la infección por VIH. Se asume que estás células son producidas de manera proporcional al número promedio de células Tα disponibles, con tasa de proliferación γ y con tasa de eliminación natural ω (no asociada con la infección por VIH), de manera que en un tiempo t se producen γTα células citotóxicas y se eliminan en promedio ωTc células. El efecto citotóxico se refleja en una disminución de las células infectadas a razón de ρT*Tc células en promedio. Con estas consideraciones el modelo toma la forma,
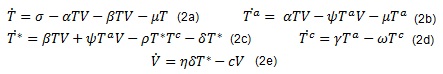
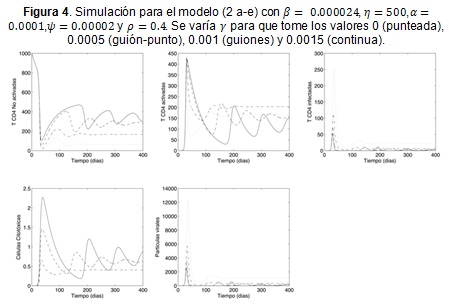
Se hace una simulación preliminar de este modelo considerando los mismos parámetros del cuadro 1 y para β = 0.000024, η = 500, α = 0.0001, ψ = 0.00002 que fueron escogidos a partir de la simulación del modelo original (1 a-d) y que se corresponden con los parámetros usados por (3, 8, 19). Los parámetros asociados con la respuesta citotóxica se determinaron con base en los comentarios de (8,20),es decir ρ = 0.3 y ω = 0.2; el valor de se varía para ilustrar el efecto que la respuesta citotóxica tiene en el control del virus.
La Figura 4 permite observar que efectivamente la respuesta citotóxica tiene efectos positivos en cuanto disminuye notablemente el promedio células T infectadas y la carga viral, por lo menos en las fases iniciales de infección. Sin embargo, debe tenerse en cuenta que las poblaciones de células sanas presentan grandes oscilaciones, lo que podría traducirse en la susceptibilidad del paciente frente a las enfermedades oportunistas.
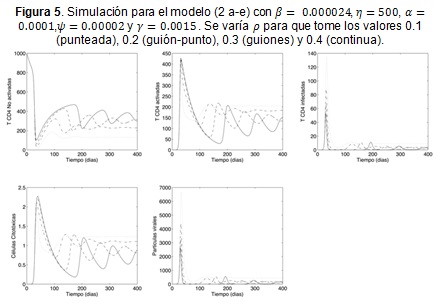
En la Figura 5 se utilizan β = 0.000024, η = 500, α = 0.0001, ψ = 0.00002 y γ = 0.0015 y se varía el parámetro ρ lo que permite observar el efecto que la acción citotóxica tiene en la dinámica. Se observa que para valores pequeños el comportamiento de las poblaciones es más uniforme, en el sentido que las variables alcanzan el estado de equilibrio en poco tiempo. Por el contrario, para valores altos de ρ, se presentan amplias oscilaciones en las células T no activadas para VIH, en las células Tα activadas y especializadas para VIH, y en las mismas células Tc citotóxicas. Tales oscilaciones ponen de manifiesto que un aumento en la acción citotóxica ρ conlleva a un incremento en las células T, y aun decremento de las células Tα; lo que se debe a la reducción que se da en las células infectadas y las partículas virales.
El R0 en epidemiología se interpreta como un indicador de riesgo; en este caso, determina la posibilidad que tiene un individuo de adquirir la infección a partir de una situación de riesgo. En este trabajo se verifica que bajo la condición R0 < 1 la infección no logra establecerse en el individuo, ese hecho también se puede constatar mediante la simulación.
Para controlar la infección es conveniente disminuir las probabilidades de infección β, ψ ó ambas, ya que se observó que al disminuir esos parámetros se reduce el impacto del brote de infección inicial y esto tiene efectos en el futuro de la infección. Una situación semejante se presenta con la reducción del parámetro η.
La respuesta citotóxica en la fase inicial de la infección por VIH juega un papel determinante en la reducción de la infección, y puede decirse que un paciente que presenta alta respuesta citotóxica en la fase inicial de la infección, puede mantener cargas virales bajas y conteos de células T más altos. Es necesario que bajo estas circunstancias haya una vigilancia frecuente del estado inmunológico del paciente y de su respuesta a enfermedades oportunistas dado el alto nivel de variabilidad que presenta su sistema inmune (amplias oscilaciones).
Agradecimientos: El presente trabajo fue financiado por la Universidad del Quindío, a través de la Vicerrectoría de Investigaciones. Proyecto de investigación 573.
REFERENCIAS
1. Alcami J. Avances en la inmunopatología de la infección por el VIH. Enferm Infecc Microbiol Clin. 2004;22(8):486-96. [ Links ]
2. Kirschner DE, Stewart T. Chang, Thomas W. Riggs, Nicolas Perry, Jennifer J. Linderman. Toward a multiscale model of antigen presentation in immunity. Immunological Review. 2007; Vol. 216: 93-118. [ Links ]
3. Orellana JM. Optimal drug scheduling for HIV therapy efficiency improvement.Biomedical Signal Processing and Control. 2011; (6):379-386. [ Links ]
4. Adam A, Wu H. Relationships between antiviral treatment effects an biphasic viral decay rates in modeling HIV dynamics. Mathematical Biosciences. 1999; 160. 63-82. [ Links ]
5. Adams B.M. Banks H.T. Davidian M. Hee-Dae K. Tran H.T. Wynne S.N. Rosenberg E.S. HIV dynamics: Modeling, data analysis, and optimal treatment protocols. Journal of Computational and AppliedMathematics. 2005;184:10-49. [ Links ]
6. Cazelles B, Chau N.P. Using the Kalman Filter and Dynamic Models to Assess the Changing HIV/AIDS Epidemic. MathematicalBiosciences. 1997; 140: 131-154. [ Links ]
7. Cleary S, Mooney G, McIntyre D. Claims on health care: a decision-making framework for equity, with application to treatment for HIV/AIDS in South Africa. HealthPolicy and Planning. 2011; 26:464-470. [ Links ]
8. Culshaw RR, Ruan S, Spiteri RJ. Optimal HIV treatment by maximising immune response. J. Math. Biol. 2004;48:545-562. [ Links ]
9. Del Valle S., Morales Evangelista A, Velasco MC, Kribs-Zaleta CM, HsuSchmitz SF. Effects of education, vaccination and treatment on HIV transmission in homosexuals with genetic heterogeneity. Mathematical Biosciences. 2004; 187(2):111-33. [ Links ]
10. Nelson P, Murray J, Perelson A. A model of HIV-1 pathogenesis that includes an intracellular delay. Mathematical Biosciences. 2000; 163:201-215. [ Links ]
11. Revilla T, García G. Fighting a virus with a virus: a dynamic model for HIV-1 therapy. MathematicalBiosciences. 185. 2003; 191-203. [ Links ]
12. Hyungbo S, Seung-Ju H, Chung Choo Chung, Sang Won N, and Jin Heon S. Optimal Scheduling of Drug Treatment for HIV Infection: Continuous Dose Control and Receding Horizon Control. International Journal of Control, Automation, and Systems. 2003; (1)3:282-288. [ Links ]
13. Stengel R. Mutation and control of the human immunodeficiency virus. Mathematical Biosciences. 2008; 213: 93-102. [ Links ]
14. Wandeler G. Keiser O, Hirschel B, Günthard HF, Bernasconi E, Battegay M. et al. A Comparison of Initial Antiretroviral Therapy in the Swiss HIV Cohort Study and the Recommendations of the International AIDS Society-USA. PLoS ONE. 2011; 6(12). [ Links ]
15. Zaric GS, BayoumiAM, Brandeau ML, Owens DK. The Cost Effectiveness of Counseling Strategies to Improve Adherence to Highly Active Antiretroviral Therapy (HAART) Among Men Who Have Sex with Men. Med Decis Making. 2008; 28(3): 359-376. [ Links ]
16. Panel on Antiretroviral Guidelines for Adults and Adolescents. Guidelines for the use of antiretroviral agents in HIV-1-infected adults and adolescents. Department of Health and Human Services. January 10, 2011; 1-166. [ Links ]
17. Montoya CJ, Moreno ME, Rugeles MT. Reacciones y alteraciones del sistema inmune durante la infección por el VIH-1.Infectio. 2006; 10(4): 250-265. [ Links ]
18. Wodarz D, Hamer DH. Infection dynamics in HIV-specific CD4 T cells: Does a CD4 T cell boost benefit the host or the virus? Mathematical Biosciences. 209 (2007) 14–29. [ Links ]
19. Srivastava PK, Chandra P. Modeling the dynamics of HIV and CD4+ T cells during primary infection. Nonlinear Analysis: Real World Applications. Differential Equations and Dynamical Systems, Vol. 16, Nos. 1& 2; 2008 pp. 77-100. [ Links ]
20. P. Kr. Srivastava P. Chandra. Modeling the dynamics of HIV and CD4+ T cells during primary infection, Nonlinear Analysis: Real World Applications (2008), doi:10.1016/j.nonrwa.2008.10.037 8 Elsevier Ltd. All rights reserved. [ Links ]
21. Stilianakis NI, Dietz K, Schenzle D. Analysis of a Model for the Pathogenesis of AIDS. Mathematical Biosciences.1997; 145:27-46. [ Links ]
22. Perelson AS. Nelson PW. Mathematical analysis of HIV-1 dynamics in vivo. SIAM/REWIEW. 1999; 41(1):3-44. [ Links ]
