Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Los paisajes de los Andes han sido reconocidos a nivel mundial por su diversidad de aves (Herzog & Kattan, 2011), además de sus altos niveles de endemismo y amenaza (Myers et al., 2000). La alta heterogeneidad ambiental generada por la variabilidad topográfica, climática y edáfica en los paisajes andinos ha tenido efectos importantes sobre la diversificación de la biota regional (Kattan et al., 2006). Sin embargo, estos paisajes han sido transformados por la ocupación humana, eliminando y fragmentando el hábitat de muchas especies, por lo que este proceso es considerado una de las principales amenazas a la biodiversidad a escala global (Sala et al., 2000).
Varios autores sugieren que la transformación de estos paisajes ha tenido un efecto negativo sobre la composición de la avifauna, y se han descrito algunos patrones de extinción de especies en los que las aves más dependientes de los bosques se extinguen primero (frugívoros grandes, insectívoros de sotobosque, entre otros), teniendo como consecuencia unos ensamblajes de aves empobrecidos y compuestos principalmente de especies generalistas (Kattan et al., 1994; Renjifo,1999; Renjifo, 2001; Sekecioglu, 2012). Varios estudios han comparado la avifauna entre los bosques y las áreas agrícolas, encontrando en general una mayor riqueza de especies en los bosques (Estrada et al., 1997; Daily et al., 2001).
En contraste con lo anterior, diferentes autores sostienen que la heterogeneidad de los paisajes transformados podría tener una relación positiva con la riqueza de especies (Bennett et al., 2006; Benton et al., 2003), en la medida que permanezcan en el mosaico parches de vegetación natural y el manejo de los agroecosistemas favorezca la conectividad entre los parches (Burel & Baudry, 2005; Harvey et al., 2008). Estas características podrían considerarse “típicas” del paisaje rural cafetero de Colombia, conformado por diversos tipos de vegetación en donde además de remanentes de vegetación natural, se integran árboles en los agroecosistemas (Rodríguez et al., 2008) y se mantienen corredores de vegetación riparia que favorecen la conectividad en el paisaje (Marín et al., 2008).
Aunque muchos estudios han documentado una pérdida dramática de especies de interior de bosque en los hábitats transformados (Renjifo, 1999), también se ha demostrado que muchas de las especies de bosque pueden usar otros tipos de hábitat en el paisaje (Gascon et al., 1999; Daily et al., 2001; Renjifo, 2001; Hughes et al., 2002; Luck & Daily, 2003). Adicionalmente, varios estudios sugieren que la composición de los ensamblajes de aves difiere significativamente entre tipos de vegetación, teniendo como consecuencia una mayor riqueza de especies en el paisaje por efecto de la complementariedad de los hábitats (Naidoo, 2004).
Los ensamblajes de aves en los paisajes rurales de las regiones cafeteras de Colombia han sido estudiados en varias localidades por diversos autores (Verhelst et al., 2001; Rojas-Díaz et al., 2004; Kattan et al., 2006; Botero et al., 2012; Lentijo & Botero, 2013; Henao-Isaza et al., 2014; Botero et al., 2014; Carranza-Quiceno & Henao-Isaza, 2015; García & Loaiza, 2016; López, 2018) y todos reconocen en alguna medida su aporte a la conservación de las aves. No obstante, la mayoría de los estudios se han realizado en localidades puntuales que solo permiten el registro de una porción de la riqueza total de especies; por lo que la información disponible sobre la riqueza de aves en los paisajes rurales del oriente de Risaralda es a escala local.
Tradicionalmente, las diferencias en la composición de los ensamblajes de especies han estado asociadas a la distancia geográfica entre las localidades (Kattan et al., 2006). El propósito de este trabajo es reconocer la riqueza de aves de los paisajes rurales estudiados y explorar el aporte de los diferentes tipos de vegetación en el mosaico a la diversidad alfa, beta y gamma del ensamblaje de especies, partiendo de la hipótesis de que existe una relación positiva entre la heterogeneidad del mosaico y la riqueza regional de especies (Benton et al., 2003). Finalmente, planteamos la necesidad de incluir los paisajes rurales dentro de las estrategias de conservación de la biodiversidad a escala local y regional.
Materiales y métodos
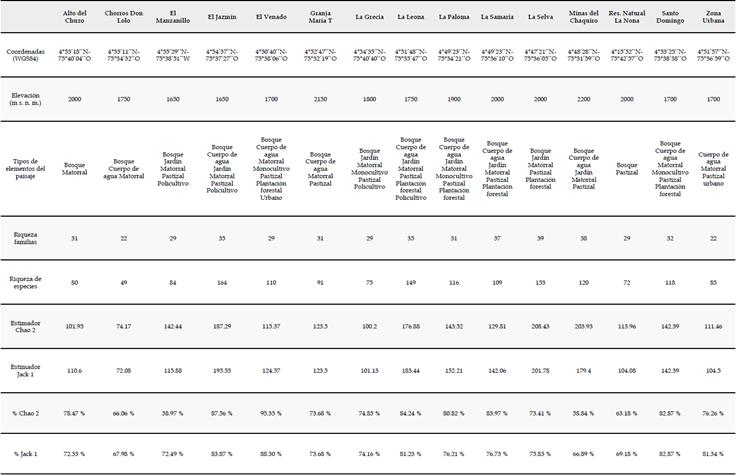
Área de estudio. Describimos la avifauna presente en el paisaje rural del oriente de Risaralda con base en registros de aves acumulados entre los años 2013 y 2017 en 15 localidades de los municipios de Santa Rosa de Cabal y Marsella: Alto del Chuzo (Chu), Chorros de Don Lolo (Cho), El Manzanillo (Man), El Jazmín (Jazm), El Venado (Ven), Granja María T (MT), La Grecia (Gre), La Leona (Leo), La Paloma (Pal), La Samaria (Sam), La Selva (Sel), Minas del Chaquiro (Chaq), La Nona (Non), Santo Domingo (SDom) y la zona urbana de Santa Rosa de Cabal (ZUrb) (Figura 1, ver detalles en el Anexo 2), abarcando un rango altitudinal entre 1600 y 2200 m s. n. m.
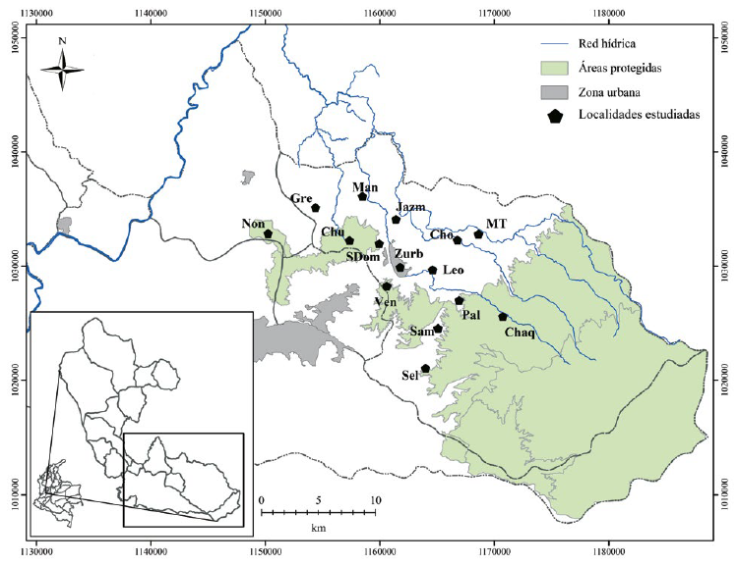
Figura 1 Ubicación del paisaje rural y las localidades donde se estudió la avifauna en Risaralda, Andes centrales de Colombia.
Todas las localidades que estudiamos incluyen varios tipos de vegetación entre las que conforman el mosaico de paisaje: bosque, plantación forestal, matorral, policultivo, monocultivo, pastizal, jardín, zona urbana y cuerpo de agua. Estos tipos de vegetación difieren notablemente en su estructura y en su composición florística. Además, varían en su representatividad en cada localidad, configurando así un mosaico muy heterogéneo espacialmente (Figura 2).

Figura 2 Diversas configuraciones del mosaico en el paisaje rural andino del oriente de Risaralda, Colombia, en las que se encuentran bosques, potreros, cultivos forestales y viviendas humanas. A) La Leona, B) La Grecia, C) Granja María T, D) El Manzanillo, E) Minas del Chaquiro.
Muestreo de aves. Documentamos las aves mediante recorridos de observación, registros auditivos y capturas en redes de niebla. En total acumulamos datos de 421 recorridos de observación de distancia variable, realizados en la mañana (6:00 a 10:00 h) o en la tarde (15:00 a 18:00 h), totalizando aproximadamente 1250 horas de observación. En cada recorrido anotamos las diferentes especies de aves observadas o escuchadas y registramos el tipo de vegetación en el que obtuvimos cada registro. Además, capturamos aves con redes de niebla (12 x 2.5 m) en la mayoría de las localidades estudiadas, acumulando un total de 6350 horas-red. Adicionalmente, para completar los datos de riqueza de especies, también incluimos observaciones ocasionales realizadas por fuera de los recorridos. Los registros de aves en sobrevuelo solo los tuvimos en cuenta en los análisis de diversidad por localidades, pero no fueron asignados a ningún tipo de hábitat.
La identidad de las especies la determinamos usando las guías de campo de Hilty & Brown (2001) y McMullan (2011). Para la nomenclatura taxonómica seguimos la propuesta de la SACC (Remsen et al., 2018; en: http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm) y para la información sobre el status de conservación revisamos la base de datos de IUCN (2018; https://www.iucnredlist.org/search) y el Libro rojo de las aves de Colombia (Renjifo et al., 2014). El endemismo de cada especie lo determinamos con base en la revisión de Avendaño et al. (2017).
Análisis de datos. Calculamos la representatividad del muestreo de aves a escala de paisaje y a escala local con base en los estimadores Chao2 y Jack1, obtenidos mediante el software EstimateS 8.2 (Colwell, 2009; en: http://viceroy.eeb.uconn.edu/EstimateS). Para evaluar las diferencias en la composición de especies entre localidades, calculamos la similitud de Jaccard (S) como la proporción de especies compartidas entre pares de localidades. Para evaluar el recambio espacial del ensamblaje de aves, usamos una regresión lineal entre la distancia geográfica (km en línea recta) y la similitud de Jaccard (S) entre pares de localidades. De igual forma, usamos una regresión lineal para evaluar si existe alguna relación entre la riqueza de tipos de vegetación y la riqueza de aves en cada localidad. Para estos análisis usamos el software Infostat disponible en http://www.infostat.com.ar (Di Rienzo, 2016)
Finalmente, evaluamos el aporte de cada tipo de hábitat a la riqueza regional del ensamblaje, teniendo como base el modelo aditivo de la diversidad regional (γ), en el que γ = α + β (Veech et al., 2002), que puede estar más relacionado con los componentes espaciales de la diversidad (Kattan et al., 2006), y en el que α = al número de especies presentes en cada tipo de hábitat, β = especies ausentes en cada tipo de hábitat y γ = al número total de especies del ensamblaje; de esta forma, calculamos la proporción de diversidad alfa (α) de cada localidad con respecto a la diversidad regional (γ). Adicionalmente, evaluamos la complementariedad de los ensamblajes en cada tipo de vegetación usando las medidas de disimilitud de Jaccard (1 - S), a partir de las cuales construimos un dendrograma, usando el método de ligamiento promedio.
Resultados
En el paisaje rural cafetero del oriente de Risaralda obtuvimos un total de 11732 registros, que corresponden a 22 órdenes, 50 familias y 309 especies de aves. El listado representa un 85 % del total de especies calculadas por el estimador Chao 2 y un 85.5 % de las calculadas por el estimador Jack 1. En las localidades que estudiamos, alcanzamos una representatividad promedio de 75.90 % ± 10.61 % para el estimador Chao 2 y de 76.21 % ± 6.24 % para el estimador Jack 1 (Anexo 2).
A nivel taxonómico, las familias con mayor riqueza de especies dentro del ensamblaje fueron Thraupidae (49 spp.), Tyrannidae (45 spp.), Trochilidae (27 spp.), Parulidae (19 spp.), Furnariidae (14 spp.) y Picidae (10 spp.) (Anexo 1). Adicionalmente, hacen parte del ensamblaje 29 especies migratorias boreales pertenecientes a 10 familias, que se establecen en el paisaje rural entre los meses de septiembre y marzo de cada año; una especie introducida (Columba livia) y 22 especies que pueden considerarse errantes que incursionan en el paisaje desde otras franjas altitudinales y tienen muy pocos registros (<3). Por otro lado, entre las especies residentes, los registros más frecuentes fueron de las especies Thraupis episcopus, Turdus ignobilis, Tyrannus melancholicus, Stilpnia vitriolina y Bubulcus ibis, los cuales suman el 11.49 % de los registros totales.
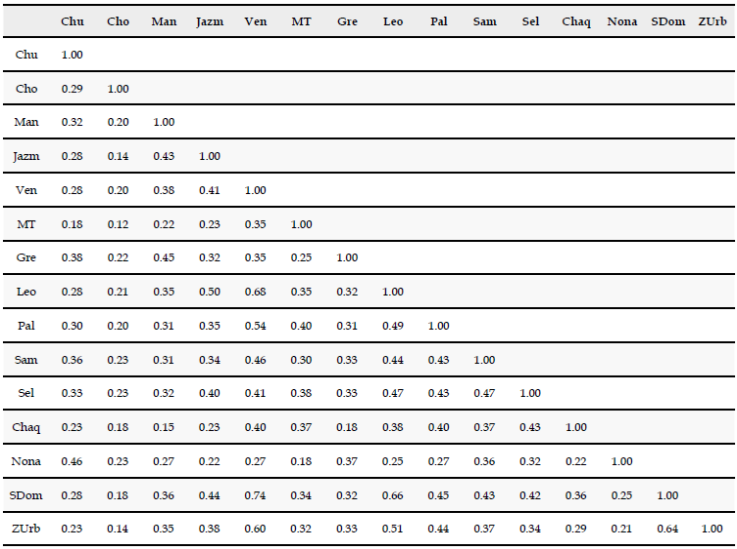
Entre las diferentes localidades que estudiamos, las que mayor riqueza de especies presentaron fueron El Jazmín (164 spp.), La Selva (154 spp.), La Leona (149 spp.) y Minas del Chaquiro (120 spp.). Las localidades que presentaron mayor afinidad en la composición de la avifauna fueron El Venado, Santo Domingo, La Leona, zona urbana Santa Rosa de Cabal y la Paloma, con valores de similitud superiores a 0.5 (Anexo 3). Encontramos una relación inversa, aunque no significativamente fuerte, entre la distancia geográfica y la similitud en la composición de la avifauna de cada localidad (R2 aj = 0.22; gl = 103; p<0.001), pero el modelo explica una baja proporción de la variabilidad de los datos (Figura 3). Por otro lado, encontramos que además existe una relación entre la riqueza de tipos de vegetación y la riqueza de aves en cada localidad (R2 aj = 0.40; gl = 14; p = 0.006) (Figura 4). Estos resultados sugieren que el recambio en la composición de los ensamblajes locales de aves en el paisaje estudiado podría estar influenciado tanto por la distancia geográfica como por la composición del paisaje en cada localidad.
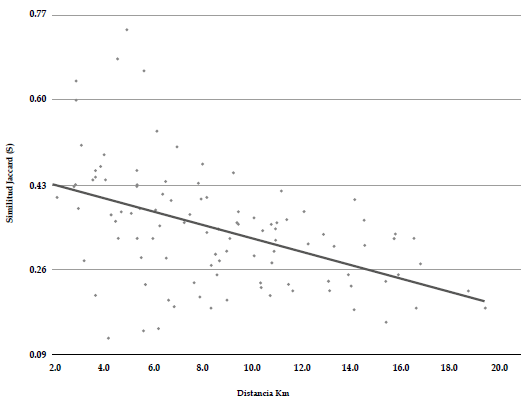
Figura 3 Relación entre la similitud en la composición de aves y la distancia geográfica entre 15 localidades del paisaje rural del oriente de Risaralda, Colombia.
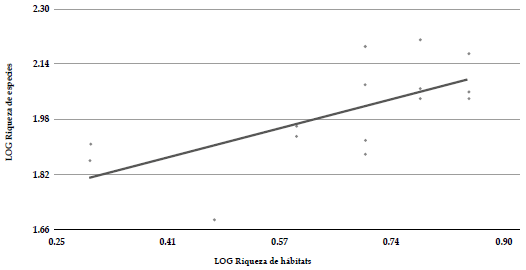
Figura 4 Relación entre la riqueza de tipos de vegetación por localidad y la riqueza de especies de aves en 15 localidades del paisaje rural del oriente de Risaralda, Colombia.
Los tipos de vegetación que presentan mayor afinidad en la composición de su avifauna, corresponden a aquellos con mayor diversidad en su estructura vertical (policultivo, matorral, bosque y jardín), mientras que los cuerpos de agua, las zonas urbanas y los monocultivos contienen conjuntos de especies más diferenciados respecto a los demás hábitats (Figura 5). Sin embargo, todos los tipos de vegetación en el paisaje rural aportan a la diversidad. En el ensamblaje de aves del paisaje estudiado, destacamos la importancia de los bosques (ca. 80 % de las especies) y los matorrales (más del 50 %); además observamos 103 especies (33.1 %) usando los pastizales y 41 especies (13.2 %) usando el hábitat urbano (Figura 6).
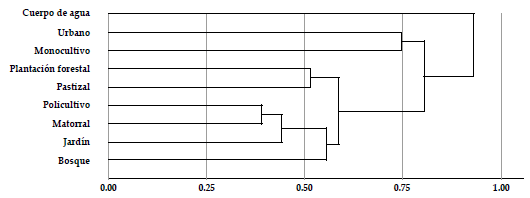
Figura 5 Afinidades en la composición de la avifauna de los tipos de vegetación presentes en el paisaje rural del oriente de Risaralda, Colombia.
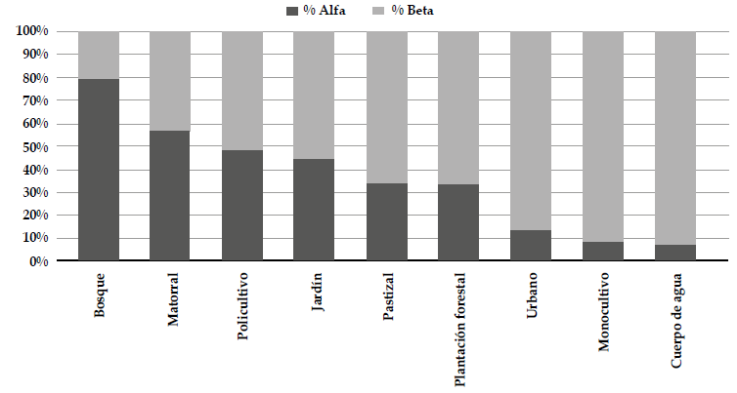
Figura 6 Componentes alfa y beta de la diversidad de aves en cada tipo de vegetación observado en el paisaje rural el oriente de Risaralda, Colombia, con base en el modelo aditivo de la diversidad regional en el que γ = α + β (Veech et al., 2002).
Discusión
Los paisajes rurales del oriente de Risaralda, heterogéneos en su configuración espacial, albergan una representación importante de la riqueza avifaunística de los Andes centrales de Colombia. Las aves en el paisaje que estudiamos representan un 71.5 % de las 435 especies reportadas por Kattan et al. (2006) a escala regional, en un rango altitudinal más amplio. La composición taxonómica del ensamblaje de aves coincide con los patrones descritos como “típicos” de ambientes andinos intervenidos y con una alta oferta de recursos (Naranjo, 1994; Botero et al., 2014) y las especies más abundantes son principalmente omnívoras, tolerantes a la transformación del hábitat o que prefieren los hábitats transformados; sin embargo persisten también especies de interés para la conservación.
Un total de 49 especies (15.7 % del total) aparecen con solo un registro en este estudio; algunas de ellas pueden ser consideradas errantes en el ensamblaje de aves. Estas especies fueron observadas en sobrevuelo (Buteo brachyurus, Chondrohierax uncinatus, Geranoaetus albicaudatus) o corresponden a especies distribuidas en otros rangos altitudinales que incursionaron en el paisaje estudiado, tal como Buthraupis montana, Diglossa humeralis, Dixiphia pipra, Porzana flaviventer, Pachyramphus cinnmomeus, Cantorchilus nigricapillus, entre otros (Hilty & Brown, 2001). Sin embargo, aquellas especies cuya presencia en el ensamblaje está confirmada, podrían sugerir que existen poblaciones con bajas densidades y de distribución fragmentada en el paisaje (Pearman, 2002), lo que incrementa su vulnerabilidad a la extinción (Kattan, 1992). Por estas razones, es necesario incrementar el esfuerzo de muestreo para evaluar el estado de estas poblaciones, tal como lo sugirió Renjifo (2001).
Es importante resaltar la presencia de 29 especies migratorias (9.33 % del total) en el ensamblaje de aves del paisaje rural. Esta proporción es menor que en ensamblajes locales tal como la zona urbana de Pereira (Londoño, 2011), la reserva de Planalto de Cenicafé (Botero et al., 2012), localidades cafeteras de Manizales (Lentijo & Botero, 2013) y una localidad rural del paisaje estudiado (Henao-Isaza et al., 2014), donde se reportan porcentajes de 11 % -14 %. Las migratorias más abundantes en el paisaje fueron Setophaga fusca, Mniotilta varia, Piranga rubra, Cardellina canadensis y Catharus ustulatus. Entre este grupo se destacan los registros de Passerina cyanea, la cual ha sido considerada como un visitante errático durante la migración, y la mayoría de los registros en Colombia se ubican en la costa Caribe (Avendaño et al., 2017).
El ensamblaje de aves observado en el paisaje cafetero incluye siete taxa endémicas, de las cuales una está en peligro de extinción (Penelope perspicax) y cinco de ellas son consideradas vulnerables (Leptosittaca bra- nickii, Chlorochrysa nitidissima, Dacnis hartlaubi, Sericossypha albocristata y Setophaga cerulea). También dos de las especies son consideradas casi-amenazadas (Aburria aburri y Contopus cooperi). Resaltamos la presencia del endémico carpinterito punteado (Picumnus granadensis), registrado frecuente en los matorrales del paisaje estudiado; la presencia de esta especie en áreas agrícolas fue reportada previamente por Verhelst et al. (2002).
Kattan et al. (2006) describieron los componentes espaciales de la riqueza de aves en la región del oriente de Risaralda, resaltando la alta diversidad beta asociada a la variación altitudinal. Encontraron que los ensamblajes de aves fueron más similares entre sí en localidades dentro de un mismo rango de elevación. No obstante, estos mismos autores sugieren la existencia de un componente importante de diversidad beta, representado en la diferenciación de los ensamblajes de aves entre localidades en un mismo rango altitudinal, lo cual podría estar asociado a la fragmentación de los bosques y el aislamiento de las poblaciones.
En el paisaje rural del oriente de Risaralda, es notable el recambio espacial en la riqueza de aves, a pesar de que el área abarcada sea relativamente pequeña (aprox. 400 km2) y en un rango altitudinal estrecho (600 m). Sin embargo, los resultados sugieren que las diferencias en la composición de los ensamblajes locales también pueden estar asociadas a la riqueza de tipos de vegetación en cada localidad (Haslem & Bennett, 2008). A escala de paisaje, la heterogeneidad del mosaico (evidenciada en la diversidad de tipos de vegetación) tiene un efecto positivo sobre la riqueza regional de especies, en la medida en que todos los tipos de vegetación aportan algunas especies únicas al ensamblaje de aves (Lee & Martin, 2017; Tews et al., 2004; Benton et al., 2003).
En cuanto al aporte a la diversidad de aves de los diferentes tipos de vegetación en el paisaje, es notable la riqueza de especies que albergan los bosques (Renjifo, 2001), y entre las cuales está la mayor proporción de aves especialistas de hábitat. De igual forma, si consideramos a los matorrales como estados iniciales de la sucesión de los bosques, luego del abandono de las prácticas productivas, la riqueza de aves presentes en hábitats no manejados asciende a 280 especies (90 % del ensamblaje), lo que respalda la importancia de mantener parches de vegetación natural en diferentes estados de sucesión en el mosaico, tal como lo sugieren Estrada-Carmona et al. (2014). Es importante reconocer que algunos elementos del paisaje favorecen el establecimiento de aves típicas de áreas abiertas o que son muy tolerantes a la transformación de los hábitats; de hecho, 168 especies (54.02 % del ensamblaje) fueron registradas en más de 3 tipos de vegetación diferentes por lo que pueden ser consideradas generalistas de hábitat.
En los paisajes rurales, aunque los hábitats transformados no sustituyen a los bosques, el mantenimiento de diferentes tipos de vegetación en el mosaico ha permitido el sostenimiento de una avifauna diversa por el efecto de complementariedad en la composición de los ensamblajes de cada tipo de vegetación (Haslem & Bennett, 2008). De acuerdo con el modelo aditivo de la diversidad regional (Veech et al., 2002), todos los tipos de vegetación en el mosaico aportan al mantenimiento de la riqueza regional, tanto por las especies especialistas de hábitat como por el número total de especies que usan cada tipo de vegetación, por lo que la riqueza regional (309 especies) es casi tres veces mayor que el promedio de riqueza de aves en cada tipo de vegetación (110 especies). Estas características del modelo aditivo resaltan su valor como herramienta para evaluar el aporte de la heterogeneidad del paisaje a la diversidad (Kattan et al., 2006).
Sugerimos que el éxito de las estrategias de conservación en los paisajes rurales depende del diseño y manejo de paisajes que puedan albergar tanto como sea posible de la diversidad nativa de una región a través del mantenimiento de diferentes tipos de vegetación, principalmente bosques, que incrementen la heterogeneidad del paisaje (Daily et al., 2001). Estas iniciativas son aún más importantes si se considera que existen limitaciones para la conservación exclusivamente en áreas protegidas (Chazdon et al., 2009); algunos autores incluso consideran que los paisajes rurales manejados con criterios de sostenibilidad ambiental, son la única alternativa de conservación posible en alrededor del 70 % de los Andes de Colombia (Lozano-Zambrano et al., 2006).
Existen en Latinoamérica varias experiencias de manejo integrado de paisajes que destacan la necesidad de incrementar la multifuncionalidad de los paisajes agrícolas para la producción de alimentos, el mejoramiento de la calidad de vida de las comunidades y la conservación de la biodiversidad (Estrada-Carmona et al., 2014). El aporte de los agroecosistemas a la conservación de la biodiversidad en el paisaje rural del oriente de Risaralda puede estar influenciado por su heterogeneidad, tanto a escala del mosaico (Bennett et al., 2006) como a escala local (Henao-Isaza et al., 2014), la cual se incrementa cuando se integran al sistema productivo varios estratos de vegetación (Fajardo et al., 2009; Sánchez-Clavijo et al., 2009) y podrían favorecer la presencia de especies de interés (Botero & Verhelst, 2001). En síntesis, sugerimos la necesidad de involucrar los paisajes rurales dentro de las iniciativas de conservación a nivel regional y nacional, para promover el diseño y manejo de agroecosistemas diversos e implementar estrategias de manejo del paisaje orientadas a mantener no solo los remanentes de vegetación natural, sino también la heterogeneidad y la conectividad del paisaje.