











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Vanellus chilensis se distribuye en Centroamérica y Suramérica, desde el sur de El Salvador hasta el sur de Argentina y Chile (Blake, 1977; Santos, 2010; Wiersma & Kirwan, 2018). Habita pastizales, bancos y márgenes de ríos, manglares, áreas abiertas con poca vegetación y áreas perturbadas (Antas, 2004; Ridgely & Grenfield, 2001a; McMullan & Navarrete, 2013). En Ecuador se distribuye en la Amazonia baja, las estribaciones de la región interandina del norte (Ridgely & Greenfield, 2001a; MacMullan & Navarrete, 2013) y centro, y el norte hasta la costa de Esmeraldas (https://ebird.org/home). Es una especie poco común en los pastizales de las tierras bajas del este y común en las tierras altas (McMullan & Navarrete, 2013). Esta especie ha expandido su área de distribución en Ecuador desde 1980, debido a la deforestación (Ridgely & Greenfield, 2001b; Santos, 2010).
Vanellus chilensis no ha sido estudiada en Ecuador, mientras que en otros lugares de su distribución se ha descrito las características de los huevos, nidos, polluelos, supervivencia, mortalidad (e.g., Naranjo, 1991; Liker & Székely, 1999; Marín, 2014), comportamiento parental y de grupo (Gallegos, 1984; Maruyama et al., 2010; Santos & Macedo, 2011; Serpa, 2016), uso del espacio (Gallegos, 1984), defensa y conflictos (Milléo 2002; Kis et al., 2000).
Se describen aquí algunos aspectos de la historia natural de V. chilensis, como la época de reproducción, comportamiento reproductivo, alimentación, cuidado parental, crecimiento y muda de las crías, además de las amenazas, con base en observaciones de tres parejas que habitan los alrededores del lago San Pablo, en la provincia de Imbabura, en Ecuador.
Materiales y métodos
Área de estudio. San Pablo es un lago de alta montaña ubicado en la región interandina del norte de Ecuador, en el cantón Otavalo, de la provincia de Imbabura (Gunkel, 2003; Gunkel & Casallas, 2002). Ecológicamente corresponde al herbazal inundado lacustre montano de los Andes (MAE, 2013). Tiene 3.8 km de largo por 2.4 km de ancho y 35 m de profundidad, es alimentado permanentemente por el río Itambi y varios nacimientos (Gunkel, 2003). En los alrededores del lago se encuentran asentados cerca de 20000 habitantes (Gunkel & Casallas, 2002), cuya actividad primaria es la agricultura y la ganadería (Aguilar-Consultores, 2016), y un gran porcentaje realizan actividades agropecuarias con el uso de agroquímicos. El lago es considerado eutrófico (Gunkel, 2003; Gunkel & Casallas, 2002).
Las observaciones se realizaron en tres lugares de los alrededores del lago San Pablo, cada uno con un territorio de la Vanellus chilensis, de manera directa, utilizando binoculares Redfield 12x50 (Figura 1).
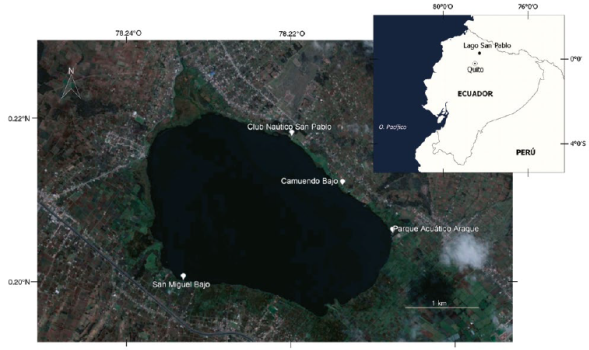
Figura 1 Ubicación de los sitios de estudio de Vanellus chilensis en el lago San Pablo, Imbabura, Ecuador: San Miguel Bajo (SMB), Parque Acuático Araque (PAA), y la planicie entre el Club Náutico San Pablo y Camuendo Bajo (CAB).
El primer sitio estuvo ubicado en la Comunidad San Miguel Bajo (SMB) (0º11´59.78´´N 78º13´58.81´´O) a 2680 m s. n. m. Es un área pantanosa situada en la margen suroccidental del lago, permanece casi todo el año inundada, pero en verano la parte norte se seca hasta la llegada de las primeras lluvias de septiembre. Otra parte se mantiene seca y cubierta por pasto kikuyo (Pennisetum clandestinum) y limita con cultivos de maíz, fréjol, frutillas, habas y papas. El pantano junto al lago está cubierto con totora (Schoenoplectus californicus y S. americanus) y entremezcladas con jacinto de agua (Eichhornia crassipes) y otras plantas herbáceas como el botoncillo (Bidens laevis) (Figura 2A). Al sur de este pantano se encuentran varias piscinas de tratamiento de aguas servidas, donde crece también el jacinto de agua, que termina por cubrir las piscinas, las cuales permanecen así durante varias semanas hasta que son limpiadas.

Figura 2 Sitios de estudio. A) San Miguel Bajo, B) Parque Acuático Araque, y C) Camuendo Bajo. Fotografías: Patricio Mena-Valenzuela.
El segundo sitio se encuentra al suroriente del lago y al borde del mismo, en el denominado Parque Acuático Araque (PAA) (0º12´22.34´´N 78º12´26.52´´O) a 2685 m s. n. m. Al sur y al borde norte del lago existe un pantano cubierto con jacinto de agua y totora, el cual limita con un área seca y cubierta con kikuyo. Este pantano permanece húmedo, pero se seca parcialmente en verano. En las cercanías del pantano se realizan actividades recreativas (Figura 2B).
El tercer sitio se encuentra en la orilla noreste del lago, es una gran planicie de aproximadamente 850 m de longitud y corresponde a la comunidad Camuendo Bajo (0º13´04.48´´N 78º13´12.13´´O y 0º12´43.03´´N 78º 12´ 49.19´´O) a una altitud de 2670 m s. n. m. Se extiende desde las instalaciones del Club Náutico San Pablo hasta el extremo sur oriental de la Comunidad Camuendo Bajo (CAB). Las partes secas están cubiertas con kikuyo, las partes húmedas con jacinto de agua, totora, Polygonum hydropiperoides y en algunos sitios sobresalen pequeños islotes de carrizo (Arundo donax) (Figura 2C).
Los pobladores de las cercanías del lago mantienen perros como mascotas, y algunos de ellos están acostumbrados a cazar aves y buscarlas en sitios de postura y crianza de aves acuáticas silvestres, como Fulica ardesiaca, Anas georgica, Gallinula galeata y Rallus aequatorialis. Las localidades de CAB y PAA son visitadas por turistas; los fines de semana hay una gran afluencia y en ocasiones los jóvenes y niños molestan a Vanellus chilensis y a otras especies.
Observación de comportamiento. Los territorios escogidos fueron visitados 11 fines de semana consecutivos con el fin de observar a las crías recién eclosionadas y determinar la fecha de eclosión de los huevos, considerando que la cría puede permanecer un día en el nido antes de salir a buscar el alimento (Marín, 2014).
El comportamiento, crecimiento y muda de las crías se observó en una cría sobreviviente de las dos nacidas de la pareja de SMB, y se complementa con algunas observaciones de los individuos de los tres territorios.
En cada visita las observaciones duraron en promedio 1h y 45m, y fueron hechas en diferentes momentos del día (mañana, medio día y tarde). En el PAA iniciaron el 5 de julio de 2015; el área fue visitado 16 veces y acumula un total de 28 horas de observación; en SMB inició el 12 de diciembre de 2015 y fue visitado 40 veces, acumulando 100 horas de observación. La superficie de los territorios de las parejas observadas, así como el área de crianza en SMB fue calculada en polígonos formados con las coordenadas de los registros periféricos de las parejas de cada territorio y colocados en Google Earth.
En cada observación, las aves fueron registradas mediante fotografías y ocasionalmente en video. La terminología para la descripción del comportamiento fue tomada de Gallegos (1984).
Resultados
Se observó un comportamiento territorial, en el que los tres territorios se encuentran cerca al agua, y los individuos están en parejas o con su prole hasta que se dispersan. Los adultos tienen un comportamiento defensivo frente a los intrusos.
Cópula. La cópula fue observada en SMB el 29 de abril de 2017. Ocurrió en el pantano adyacente a la totora de la orilla del lago. En 20 minutos (10:50-11:10) se observó a esta pareja copular tres veces, a las 10:50, las 11:07 y las 11:10. La cópula inicia con el acercamiento mutuo de la pareja, luego la hembra se coloca a un lado del macho a aproximadamente 55 cm, los dos miran a su alrededor moviendo la cabeza de un lado al otro y a veces se miran entre ellos, el macho se acerca por detrás ligeramente agazapado y se coloca detrás, luego camina hacia ella, que se agacha para que el macho suba a su espalda, una vez arriba la hembra se incorpora con la cabeza hacia abajo y las alas ligeramente abiertas, el macho se mantiene con las patas en la espalda de la hembra sacudiendo la cola, hasta que la hembra levanta la cabeza y el macho cae al suelo agitando las alas para mantenerse en equilibrio. Una vez en el suelo se pueden ver los espolones en el álula. Finalmente, el macho mira al lado opuesto de la hembra y ella se aleja unos pasos, para continuar en la búsqueda de alimento. Mientras dura la monta (2.42 segundos) los dos emiten un sonido vibrante y bajo (http://www.xeno-canto.org/423697).
Eclosión de los huevos. La cría de la pareja del PAA fue observada por primera vez el 28 de noviembre de 2015. Se presume que la eclosión del huevo ocurrió alrededor del 22 de noviembre de 2015. La segunda nidada (una cría) de la misma pareja del PAA eclosionó el 9 de septiembre de 2016 y los huevos de la tercera nidada (dos crías) eclosionaron el 9 de marzo de 2017.
La pareja de SMB tuvo en su primera nidada dos crías, que fueron observadas por primera vez el 5 de febrero de 2016, y se presume que la eclosión del huevo ocurrió alrededor del 28 de enero de 2016; la segunda nidada, de una sola cría, fue observada el 21 de enero de 2017 y la eclosión del huevo ocurrió posiblemente entre la segunda y tercera semanas de octubre de 2016. La pareja de CAB no tuvo crías durante todo el periodo de estudio. Las eclosiones de los huevos en los dos lugares ocurrieron en meses de baja (14.45-58.15 mm) y alta precipitación (72.75-95.15 mm).
Supervivencia de las crías y dispersión. Un día después de eclosionados los huevos, las crías son capaces de seguir a sus padres caminando en búsqueda de alimento, sin capacidad de vuelo, por lo que están expuestas a potenciales predadores que habitan el área como el águila pechinegra (Geranoaetus melanoleucus), el gavilán alicastaño (Parabuteo unicinctus), el cernícalo americano (Falco sparverius) y la comadreja andina (Mustela frenata).
La cría de la primera pareja (PAA) se desarrolló y creció junto a sus padres durante 7 meses y 17 días. La última vez fue observada el 30 de junio de 2016. Luego a los padres se los vio solos. Se presume que se alejó en busca de un nuevo territorio.
La segunda pareja tuvo dos crías; una de ellas fue observada hasta el 13 de marzo de 2016, y se presume que murió después de cuatro semanas y media. La segunda creció junto a sus padres, para separarse después de aproximadamente ocho meses; la última vez junto a ellos fue el 24 de septiembre de 2016.
Tamaño del territorio, superficie del área de crianza y alimentación. Las parejas observadas mostraron un comportamiento territorial. El tamaño de los territorios fue variable: el más pequeño cubre una superficie de 6.9 ha (CAB); le sigue el PAA con 7.11 ha y el de SMB con 28.7 ha. La superficie del área de crianza de SMB fue de 3.8 hectáreas. En este lugar nacieron dos crías y se mantuvieron allí en la fase de crianza. En los territorios de SMB y PAA, las parejas y sus crías permanecieron en los pantanos durante todo el desarrollo de la cría. Sin embargo, cuando la cría vuela ya son capaces de explorar otros lugares del territorio.
En los pantanos, los polluelos buscan el alimento por sí mismos bajo el cuidado de sus padres, allí proliferan lombrices de las que se alimentan los padres y los polluelos, aunque también comen otros invertebrados (artrópodos) y larvas de coleópteros.
Vanellus chilensis detecta lombrices y larvas de coleópteros caminando lentamente sobre el pantano golpeando el suelo con las patas para mover a las lombrices que se encuentran cerca y debajo de ellos. Una vez detectada, la atrapan con el pico, para nuevamente dar varios pasos y golpear de nuevo el suelo con una pata y luego con la otra y de esta manera continuar buscando alimento.
En el PAA se observó a una pareja usando el pantano como sitio de anidación y crianza; la misma pareja usó el mismo pantano en una segunda nidada inmediatamente después de que el juvenil se dispersó, y de igual manera la tercera nidada. Lo mismo ocurrió en SMB: una vez que se separó el juvenil de los padres, tuvieron una nueva cría. Esta pareja fue observada el 21 de enero de 2017 con un juvenil de aproximadamente cuatro meses. Sin embargo, durante la segunda nidada de la pareja de SMB ocupó un sitio más cubierto con totora en el mismo pantano. Allí se mantuvieron los primeros cuatro meses y después aprovecharon un sitio más descubierto. Las tres parejas se mantuvieron fieles a sus territorios y a sus parejas.
Crecimiento y muda de las crías. Cuando nacen, las crías son crípticas, están cubiertas de plumón de color gris oscuro con manchas negras en la espalda y la cabeza, detrás de los ojos presentan una línea gruesa oscura; el cuello y la región inferior abdominal blanco y el pecho ya presenta una gran mancha negra como en los adultos. Las plumas de la nuca aún no aparecen (Figura 3A). Mientras van creciendo mantienen el mismo patrón de color, que se va definiendo, las plumas del dorso café con sus bordes más claros con aspecto de escamado. Llama la atención la línea café oscura que va desde la frente y rodea la cabeza (Figura 3B); la cabeza y el dorso café con el borde de las plumas más claras da un aspecto escamado, las plumas de la nuca son evidentes y de color café oscuro más corta que en los adultos, la frente con una franja café oscura hasta detrás de los ojos, las mejillas blancas, garganta y cuello café claro, en la primera porción de la garganta una franja negra que termina en punta en la mitad de ella, una mancha negra en el pecho, el vientre y la primera porción de la cola blanco, la parte inferior de la cola negra con borde delgado café claro, iris ligeramente rojizo (Figura 3C); mantiene el aspecto anterior, pero casi todas las plumas escapulares definitivas son café, así como las plumas de la cola, las coberteras mayores aún con plumas con aspecto escamado, las terciarias son blancas y las rectrices negras (Figura 3D); la porción escapular con más plumas de color café y algunas del hombro castaño como en los adultos, sobre la franja negra de la frente algunas plumas blanquecinas, las coberteras mayores como en la fase anterior pero con algunas definitivas de café (Figura 3E); las plumas de la cabeza, cuello y dorso con plumas definitivas son café claro, el manto con plumas café claro, coberteras mayores con plumas definitivas en crecimiento, en el hombro con plumas castaño y azul oscuro metálico, arriba de la franja negra de la frente plumas blanquecinas, la punta del pico negro, iris rojizo sin llegar a la intensidad de los adultos (Figura 3F); bajo las alas las plumas blancas y las rectrices negras, en los hombros presenta pequeños espolones rojos (Figura 3G); casi está completa la muda, aunque las plumas de la porción escapular (hombro) no están completas, las plumas de la nuca no tan largas como en los adultos, y el cuerpo aún menor y más delgado (Figura 3H); las plumas del dorso (coberteras mayores) café verdosas, las plumas del hombro mayormente castaño, la franja negra de la frente más ancha y más obscura, las plumas de la nuca un poca más largas, las plumas del hombro no terminan de crecer, iris más rojo que en la fase anterior casi como un adulto, las plumas de la porción sobre la franja negra de la frente ancha y blanquecina, franja vertical negra de la garganta desciende hasta la mancha negra del pecho, franja superciliar café oscuro (Figura 3I); el juvenil ya es similar al adulto pero de menor tamaño, las plumas del hombro no terminan de crecer, las plumas de la nuca oscuras casi del tamaño de los adultos, el ojo aunque rojo no es como en los adultos que es rojo intenso (Figura 3J); la muda está completa y con los colores definitivos, el tamaño aún no alcanza el tamaño de los adultos y las plumas de la nuca aún más pequeñas que en los adultos (Figura 3K). En la última figura (3L) se muestra las características de uno de los padres, en donde es evidente las diferencias con la cría y el juvenil. En general los juveniles desarrollados se diferencian de los adultos por el tamaño, estos son ligeramente más pequeños y delgados, la intensidad del rojo del iris, menor cantidad de tonalidades de color de las plumas escapulares, la frente ligeramente blanca y las plumas de la nuca de menor tamaño.

Figura 3 Crecimiento y muda de Vanellus chilensis en el lago San Pablo, Ecuador. A-K) Crecimiento desde los primeros días hasta la separación de los padres. L) Padre del juvenil. Fotografías: Patricio Mena-Valenzuela.
Cuidado parental. Las crías, en las primeras semanas, debido a que aún no pueden volar, caminan confiadas por el pantano buscando alimento, mientras que los dos adultos se mantienen cerca, atentos a lo que ocurre a su alrededor, y en caso necesario ahuyentan rápidamente a cualquier intruso, aunque por su tamaño no representen ningún peligro para las crías. Los adultos persiguen a las aves pequeñas o grandes para repelerlas a picotazos. En varias ocasiones los adultos ahuyentaron a aves migratorias boreales de la familia Scolopacidae (Actitis macularius, Tringa flavipes, T. melanoleuca), y otras especies residentes como la garza bueyera (Bubulcus ibis) y la paloma doméstica (Columba livia). Los adultos también alertan del peligro a las crías o juveniles mediante gritos estridentes. En ciertos momentos del día, las crías buscan abrigo bajo las alas del adulto.
Comportamiento de defensa. Los adultos siempre se mantienen cerca al nido con huevos o con pichones para protegerlos. Al acercarse el intruso (por ej. personas o perros) Vanellus chilensis se levanta del suelo y vuela con la intención de golpearlo con los espolones de las alas, hasta ahuyentarlo. Si el intruso está muy cerca de la cría, los adultos lo atacan de manera persistente y agresiva hasta alejarlo. Mientras un adulto hace esto el otro se mantiene alerta en el suelo junto a las crías. Cuando la pareja no tiene crías que proteger, se apartan volando ante la persistencia del intruso. Mientras atacan están emitiendo su típica vocalización keh, keh, keh, keh (Ridgely & Greenfield, 2001a; http://www.xeno-canto.org/287588).
En el suelo adoptan diferentes posturas y comportamientos con el fin de engañar al intruso o alejarlo de las crías. Estas posturas son las denominadas paradas de ataque y de falso nido. La parada de ataque consiste en dirigirse al intruso de frente, caminar de un lado al otro gritando sin dejar de mirar el objetivo, agacharse, estirar la cabeza, igualando el cuello con la línea del lomo, las patas flexionadas. Cuando está de frente se ve la gran mancha negra en el pecho, las alas están ligeramente abiertas, mostrando los espolones rojos de los hombros, la cola ampliamente abierta. En ese momento vuela hacia el intruso tratando de golpearle con los espolones, da un giro para aterrizar y ponerse nuevamente de frente y volver al ataque.
La parada del falso nido se produce cuando el intruso está cerca al nido o a la cría. Entonces el adulto camina apresurado a otro lugar, para detenerse y bajar el cuerpo al suelo doblando las patas, posiblemente tratando de desorientar al intruso sobre la verdadera ubicación del nido o la cría. Esta postura también lo hacen los individuos que no ponen huevos o no tienen crías y los juveniles.
En dos ocasiones (9 de septiembre de 2016 y 9 de octubre de 2016) se observó el acercamiento de un perro por detrás de un adulto; este lo dejó acercarse hasta menos de 1 m, para luego alejarse saltando, agitando las alas y mostrando los espolones, alejándolo así de las crías. Los adultos y las crías se protegen de los perros colocándose en los sitios más anegados; desde esa posición gritan y vuelan bajo sobre los intrusos y se sitúan en lugares alejados de las crías para que los perros los persigan y así alejarlos. En 15 minutos de insistente ataque de los perros, los adultos de Vanellus chilensis los alejaron 20 veces aplicando la estrategia descrita. Después de unos 45 minutos los perros desistieron y se alejaron emitiendo alaridos. Estas maniobras de distracción, escape y ataque ejecutadas por los padres son muy efectivas al momento de ahuyentar y alejar al intruso de las crías o de ellos mismos.
Para atacar, V. chilensis también adopta varias posturas particulares en el suelo y el aire, como la de inicio del vuelo (Figura 4A), postura de ataque en el vuelo (Figura 4B-D), postura de aterrizaje (Figura 4E) y postura final (Figura 4F). En el suelo adopta la postura de vigilancia (Figura 4G), si el intruso continúa en su territorio se eleva nuevamente y ataca haciendo giros a su alrededor (Figura 4H, 4I).

Figura 4 Posturas para el vuelo de ataque de Vanellus chilensis. A) Inicio de vuelo; B, C y D) postura de ataque en vuelo; E) postura de aterrizaje; F) postura final; G) en el suelo se para a vigilar; H e I) luego vuela y persigue al intruso dando varios giros en el aire y ataca nuevamente. Fotografías: Patricio Mena-Valenzuela.
En los territorios de CAB y PAA, el macho y la hembra de Vanellus chilensis fueron muy agresivos: al acercarse la gente, gritan y atacan hasta alejarlos o alejarse. En cambio, en la localidad SMB, la pareja fue poco agresiva: ante la presencia de gente gritaban un momento y luego continuaban buscando alimento. Aquí, la pareja permitió observarla desde distancias cortas (aproximadamente 8 m) sin que se produjera el ataque, pero siempre se mantuvo vigilantes. El comportamiento agresivo de las dos primeras parejas posiblemente se debía a que sus territorios están en lugares visitados por personas, por lo que las aves se sienten amenazadas y se alteran adoptando un comportamiento defensivo.
Discusión
La reproducción de Vanellus chilensis en el lago San Pablo aparentemente no tiene una época marcada, sino que ocurre en cualquier mes del año, como sucede en otras especies de aves (Fulica ardesiaca, Gallinula galeata, obs. pers.), y como también observó Naranjo (1991) en otros lugares de su distribución.
Las respuestas defensivas de V. chilensis fueron observadas en las parejas con o sin crías. En el primer caso, la respuesta depende del intruso: cuando los intrusos son pequeñas aves de especies residentes o migratorias, los adultos las persiguen corriendo tras ellas hasta alejarlas de los polluelos, así no representen ningún peligro. Aunque no se analizó la inversión de tiempo en la defensa de los padres, era evidente que el macho es más activo en la defensa que la hembra, como también lo observaron Kis et al. (2000). En cambio, cuando los intrusos son personas o perros, la respuesta es muy agresiva y persistente, y atacan hasta alejarlas de las crías. En el segundo caso, cuando la amenaza se cierne sobre los adultos sin crías, estos atacan al intruso y después se alejan. En ninguna ocasión se observó el ataque de un depredador silvestre, los cuales son escasos en el área.
En los tres sitios de estudio se observó un territorio con una sola pareja (macho y hembra) y en ningún caso grupos constituidos por un par reproductor primario y uno o más individuos extra que defienden activamente los nidos, como observaron Walters (1982), Saracura et al. (2008) y Alves dos Santos (2009). Sin embargo, en una ocasión en el PAA se observó la llegada de dos individuos a reunirse con la pareja y dos crías residentes, ante los llamados de alarma. Los individuos recién llegados se mantuvieron gritando poco tiempo y luego abandonaron el lugar.
El tamaño de los territorios registrados en este trabajo es variable y relativamente grande en comparación a los registrados por Gallegos (1984) que en promedio ocuparon 5.4 ha. Posiblemente se deba a que la población de Vanellus chilensis del lago San Pablo es baja, no forman colonias y en los alrededores inmediatos al lago disponen de grandes superficies húmedas. Además, el área de crianza de las dos parejas observados en San Pablo estuvo dentro de sus territorios y no fuera de él, como observó Walters (1982).
En ningún momento se observó la parada de ala herida ni la parada militar; en la primera, el ave extiende el ala y la arrastra ligeramente; la segunda, en cambio, es una formación de un grupo de machos erguidos marcando el paso (Zuberbuhler, 1973; Hudson, 1974; Walter & Walter, 1980; Gallegos, 1984). En las dos parejas con cría observadas en San Pablo, la crianza de los polluelos estuvo exclusivamente a cargo de los padres y no se observó la presencia de individuos adicionales como en otros lugares (Santos & Macedo, 2011).
Vanellus chilensis ya forma parte de la avifauna de los alrededores del lago San Pablo; su establecimiento ha sido favorecido por actividades humanas que modifican el paisaje permanentemente, hecho que se evidencia en el aumento de su población en los últimos años. Sin embargo, los pantanos de postura y crianza son usados como áreas de pastoreo, son desecados o se han convertido en basureros, lo que constituye una amenaza para la especie y otras especies residentes o migratorias. Sin embargo, la agricultura y la ocupación de las áreas por los humanos causan mortalidad y reducción poblacional de V. chilensis (Milléo, 2002).
Conclusiones
Vanellus chilensis es una especie que expande su área de distribución constantemente. En los últimos años se ha establecido en alrededores del lago San Pablo y forma parte de la avifauna residente. Su comportamiento es muy parecido a lo observado en otros lugares de su distribución; sin embargo, algunos comportamientos como la parada de ala herida y la militar no se observaron en ningún momento; asimismo el cuidado de la cría no requiere de otros individuos ayudantes y no hay agrupación de territorios ni asociación temporal. Posiblemente esto se deba a que la población en San Pablo es baja y aún disponen de grandes extensiones para establecer sus territorios. Los sitios que habita la especie se encuentran amenazados por las actividades humanas, por lo que es urgente la conservación y manejo adecuado de los humedales a fin de resguardar y mantener la dinámica del ecosistema. El seguimiento de esta especie ayudará a comprender su dinámica en hábitats que comparte con otras especies.

