










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkInvestigaciones Andina
Print version ISSN 0124-8146
Investig. andina vol.15 no.27 Pereira Sept. 2013
Caracterización del gen de la dopamina (β-hidroxilasa en población mestiza colombiana
Caracterization of dopamine β-hydroxylase gene in the Colombian Mestizo Population
Caracterização do gene da dopamina β-hidroxilasa em população mestiça
Carlos A Isaza Mejía***, Julieta Henao Bonilla**, Sandra Valencia Castillo***, Leonardo Beltrán Angarita***, Ana M Sepúlveda Calvo**
* Programa de Medicina, Fundación Universitaria Autónoma de las Américas.
** Grupo de Investigación en Farmacogenética, Facultad de Ciencias de la Salud, Universidad Tecnológica de Pereira.
*** Grupo de Investigación en Salud y Comunidad, Fundación Universitaria Autónoma de las Américas.
Resumen
Introducción: la dopamina P-hidroxilasa cataliza la conversión de dopamina en norepinefrina y es blanco promisorio de intervenciones farmacológicas. Polimorfismos del gen DfiH son responsables de las diferencias individuales en el tono dopaminérgico y adrenérgico de los sistemas nervioso central y autónomo. Ya que las mutaciones defectuosas de la enzima y sus frecuencias varían entre las etnias, se justifican los estudios conducentes a la caracterización genotípica y fenotípica de la enzima en mestizos colombianos.
Métodos: determinamos las frecuencias de los alelos -2073C>T, -970C>T, 444A>G y 1603C>T del gen DfiH en 143 adultos sanos, rasgos mestizos, ambos sexos y no consanguíneos. La genotipificación se hizo por PCR-real time y minisecuenciación (SnaPshot).
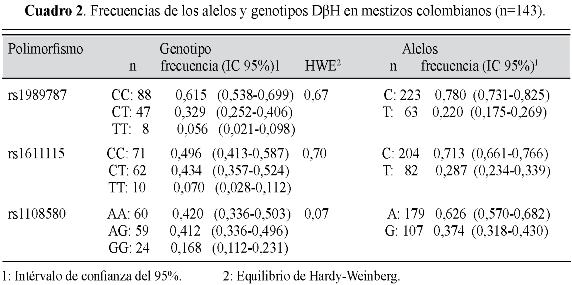
Resultados: las frecuencias de los genotipos polimórficos fueron: -2073C>T (CC 61,5%, CT 32,9%, TT 5,6%), -970C>T (CC 49,6%, CT 43,4, TT 7%) y 444A>G (AA 42%, AG 41,2%, GG 16,8%). Las tres mutaciones están en desequilibrio de ligamiento (D'=1) pero no se sustituyen mutuamente (r2<0,8). 15 personas (10,5%) tuvieron haplotipo de triple homocigoto nativo (CC/CC/AA).
Conclusión: nuestras frecuencias alélicas se asemejan a las reportadas en otros grupos mestizos latinoamericanos.
Palabras clave: Dopamina Beta-Hidroxilasa; Farmacogenética; Genotipo; Polimorfismo Genético.
Abstract
Introduction: dopamine P- hidroxylase catalyzes the conversion of dopamine to norepinephrine and it is a promising target for pharmacological inventions. DPH gene polymorphisms are responsible for individual differences in dopaminergic and adrenergic tone of the central and autonomic nervous systems. Since defective enzyme mutations and their frequencies vary among ethnic groups, it is justify the studies leading to the genotypic and phenotypic characterization of the enzyme in Colombian mestizos.
Methods: we determined the frequencies of the alleles -2073C>T, -970C>T, 444A>G and 1603C>T DfiH gene in 143 healthy adults, mestizo features, both sexes and nonconsanguineous. PCR-real time and minisequencing (SnaPshot) tecnhiques were used for the genotyping.
Results: the frequencies of polymorphic genotypes were: -2073C>T (CC 61,5%, CT 32,9%, TT 5,6%), -970C>T (CC 49,6%, CT 43,4, TT 7%) and 444A>G (AA 42%, AG 41,2%, GG 16,8%).The three alleles are in linkage disequilibrium (D'=1) but they do not replace each other (r2<0,8). 15 people (10,5%) had the homozygous triple native haplotype (CC/CC/AA).
Conclusion: our allelic frequencies are similar to those reported in other Latin American mestizo groups.
Key words: Dopamine P-Hydroxylase; Pharmacogenetics; Genotype; Genetic polymorphism.
Resumo
Introdução: a dopamina P-hidroxilasa catalisa a conversão de dopamina em norepinefrina e é alvo promissor de intervenções farmacológicas. Polimorfismos do gene DfiH são responsavéis pelas diferenças individuais no tom dopaminérgico e adrenérgico dos sistemas nervoso central e autonomo. Como as mutações defeituosas da enzima e suas frequências variam entre as etnias, se justificam os estudos conduzentes à caracterização da e fenotípica da enzima en mestiços colombianos.
Métodos: determinamos as frequencias alelos -2073C>T, -970C>T, 444A>G e 1603C>T do gene DfiH em 143 adultos sãos, traços mestiços, ambos sexos e não consanguíneos. A genotipificação se fez por PCR-real time e minisequência (SnaPshot).
Resultados: as frequências dos genotipos polimórficos foram: -2073C>T (CC 61,5%, CT 32,9%, TT 5,6%), -970C>T (CC 49,6%, CT 43,4, TT 7%) e 444A>G (AA 42%, AG 41,2%, GG 16,8%). As três mutações estão em desiquilibrio de ligamiento (D'=1) mas não se substituem mutuamente (r2<0,8). 15 pessoas (10,5%) tiveram haplotipo de triple homocigoto nativo (CC/CC/AA).
Conclusão: nossas frequências alélicas se assemelham às lembradas em outros grupos mestiças latino-americanos.
Palavras Chave: Dopamina Beta-Hidroxilasa; Farmacogenética; Genotipo; Polimorfismo Genético.
Fecha de recibo: Octubre/2012
Fecha aprobación: Mayo/2013
Introducción
La enzima dopamina P-hidroxilasa (DPH) pertenece a la familia de las monooxigenasas dependientes de ácido ascórbico, existe en forma soluble (73 kDa) o unida a membranas (77 kDa) dependiendo de la ausencia o presencia de un péptido señal. Está presente en las vesículas sinápticas del sistema nervioso central (SNC), de la médula adrenal y de las neuronas simpáticas posganglionares, donde cataliza la hidroxilación oxidativa de DA en NE (1). Al convertir DA en NE, la enzima no solo conecta el sistema dopaminérgico con el adrenérgico, sino que garantiza el adecuado balance de los dos sistemas, lo cual le permite participar en el control de numerosas funciones fisiológicas y jugar un papel esencial en el normal funcionamiento del Sistema Nervioso Central (SNC) (2, 3) y del Sistema Nervioso Autónomo (SNA) (4). Por estas razones la enzima representa un blanco promisorio de intervenciones farmacológicas.
El gen DβH que codifica la enzima, fue clonado y mapeado en el cromosoma 9q24, tiene un tamaño aproximado de 23 Kb y se compone de 12 exones que codifican para una proteína de 603 aminoácidos (NCBI, secuencia de referencia GenBank NC_000009.11). Dicho gen es polimórfico y algunos de sus alelos producen enzima con actividad defectuosa, razón por la cual en todos los grupos étnicos estudiados se encuentran individuos con actividad de la enzima normal, intermedia o baja, indicativo de una herencia codominante (5). Aunque no todas las investigaciones han obtenido los mismos resultados, existen reportes que asocian polimorfismos del gen DβH con desórdenes mentales (esquizofrenia, depresión, déficit de atención e hiperactividad) (6-11), enfermedad de Alzheimer (12), enfermedad de Parkinson (13), migraña (14, 15), conducta agresiva y susceptibilidad a la drogadicción (16-19), alterada respuesta inmune-neuroendocrina al estrés (20-22), hipertensión arterial y aumento del riesgo cardiovascular (23-25) y finalmente, puesto que la deficiencia de la enzima provoca pérdida del balance entre DA y NE, también se ha reportado respuesta alterada a fármacos relacionados con las vías biosintéticas de catecolaminas (2628).
Aunque la prevalencia alélica varía ampliamente entre las etnias, confiriéndole a cada variante distinta importancia en cada población, los alelos mejor estudiados y con mayor evidencia de sus asociaciones con reducción de la actividad enzimática son: 1) rs1989787 (-2073C>T), situado en la región del promotor, altera la transcripción; 2) rs1611115 (-970C>T), ubicado en la región del promotor, trastorna la transcripción y es el polimorfismo que más afecta la actividad de la enzima (reduce ~10 veces la actividad plasmática de la enzima); 3) rs1108580 (444A>G), situado en el exón 2; 4) rs6271 (1603C>t), situado en el exón 11 (29-32).
En este estudio nos propusimos establecer la existencia, la prevalencia y las interacciones de los alelos rs1989787, rs1611115, rs1108580 y rs6271 del gen DβH en mestizos colombianos. Tales polimorfismos han sido identificados como importantes desde el punto de vista fisiológico, patológico y farmacogenético en otros grupos étnicos. Los resultados de la caracterización del gen y su enzima contribuirán a la identificación de los procesos fisiopatológicos involucrados en varias enfermedades psiquiátricas, neurológicas y cardiovasculares, así como en la respuesta a medicamentos utilizados en tales condiciones. Este tipo de investigaciones genera la información necesaria para el empleo de fármacos más acorde con el perfil farmacogenético del mestizo, evitando la prescripción basada en datos obtenidos en grupos étnicos distintos al nuestro.
Materiales y métodos
Individuos. Para la identificación de los polimorfismos del gen DβH se enrolaron 143 voluntarios (para un estimativo de 10% de individuos homocigotos mutados del alelo rs1611115, de acuerdo con la frecuencia en población colombiana, con d=0.05, se aplica la fórmula: n = z2pq/ d2 = (1.96)2 X 0.1*0.9/0.052 =138,3), con edades entre 16 y 44 años, de rasgos fenotípicos mestizos, de ambos sexos, no consanguíneos entre sí y clínicamente sanos. Fueron excluidas las personas que consumían medicamentos y las mujeres embarazadas. Una vez firmado el consentimiento informado, a cada voluntario se le registraron sus datos personales y se le tomó muestra de mucosa bucal para su respectiva genotipificación. Este protocolo fue aprobado por el Comité de Bioética de la Universidad Tecnológica de Pereira, en la categoría de investigación con riesgo mínimo, según el artículo 11 del literal b de la resolución No. 008430 de 1993 del Ministerio de Salud de Colombia.
Genotipificación. Se llevó a cabo en el Laboratorio de Genética Médica de la UTP. Previa extracción del ADN obtenido mediante CHELEX-100 a partir de células bucales, se procedió a la amplificación de cuatro fragmentos del gen DBH en el cual están incluidos los SNPs a ser estudiados. La PCR de los fragmentos se hizo en un volumen total de 8 (µl, que contenían 2 µl de DNA genómico, 4 (µl QIAGEN Multiplex PCR Master Mix y 0,8 de la mezcla de primers 2 (µM (cuadro 1).

Para la amplificación de los fragmentos se utilizó un termociclador automático (DNA Thermal Cycler MJ Research) con los siguientes parámetros: un paso inicial de calentamiento por 11 min a 95°C, seguido por 30 ciclos de amplificación consistentes en desnaturalización a 94oC por 20s, annealing/extension a 59°C por 3 min y una extensión final a 60oC por 20 min. El exceso de primers y nucleótidos se removió adicionando 1 µL de ExoSAP-IT (Amersham Pharmacia) a 2,5 µl del producto amplificado e incubando a 37°C por 45 min, seguidos de 15 min a 85°C. Usando este producto de pre-amplificación como plantilla se efectuaron las reacciones de multiplex para la detección de los SNPs, utilizando el método de minisecuenciación o extensión de sondas (SBE, single base extention).
La reacción SBE se realizó en un volumen total de 6 µl, los cuales contienen 2 µl del producto de pre-amplificación, 1 µl de SNaPShot Reaction Mix (Applied Biosystems), 1,5 µl de la mezcla de primera SBE con concentración 0,2 µM (cuadro 1) y 1,5 µl de agua desionizada. Los tubos se pusieron en un termociclador automático (DNA Thermal Cycler MJ Research) para su amplificación por 30 ciclos, usando las siguientes condiciones: desnaturalización a 96°C por 10 s, annealing a 55oC por 5 s y extensión a 60oC por 30 s.2 µl del producto de la reacción SBE se mezclaron con 9 µl de Hi-Di formamida y 0,5 GeneScan-120 Liz internal size standard (Applied Biosystems) y se analizaron por electroforesis capilar (CE, capillary electrophoresis) usando el ABI Prims 3100-Avant Genetic Analyzers (Applied Biosystems), con capilar de 36 cm de longitud y polímero POP-4 (Applied Biosystems), con el módulo de corrida SNP36-POP-4. Los datos fueron analizados por el color de los picos y el tamaño de los fragmentos, mediante el software Genemapper v3.2 (Applied Biosystems).
Para la amplificación y detección del polimorfismo rs1989787 se empleó la metodología Real Time PCR. Con esta técnica se localizó y amplificó un segmento del ADN para ser detectado simultáneamente con una sonda marcada con fluorescencia específica para el SNP, con el siguiente código de la casa Applied
Biosystem, C_12020327_10 (cuadro 1). Cada reacción de PCR se realizó en un volumen de 25 µl conteniendo 12,5 µl Genotiping Master Mix, 0,625 µl de primer y 0,8 µl del DNA extraído mediante CHELEX-100. En cada PCR se utilizaron controles positivo y negativo. La amplificación y detección se realizó en el equipo SmartCycler, con las siguientes condiciones: un calentamiento inicial de 95°C durante 60 s, seguido por 60 ciclos que consisten en 15 s a 92°C y 60 s a 60°C. De acuerdo con la visualización en el software Smartcycler se identificó la presencia del SNP nativo o mutado.
Manejo estadístico. Las frecuencias alélicas se calcularon a partir del número observado de cada alelo. El equilibrio de Hardy-Weinberg se estableció mediante la prueba Chi cuadrado. Los análisis haplotípicos se hicieron con el programa Haploview, versión 3.32. Los demás análisis estadísticos se realizaron con el software SPSS 19 para Window, se utilizaron intervalos de confianza del 95%.
Resultados
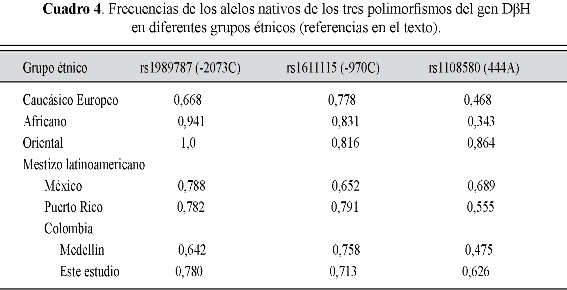
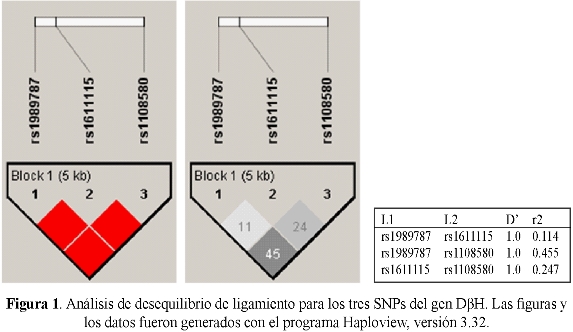
La variante alélica rs6271 fue monomórfica en nuestra muestra y se descarta para los análisis subsiguientes. En el cuadro 2 se presentan las distribuciones alélicas y genotípicas de los 143 mestizos estudiados (41,3% hombres; edad: 20±3,6 años) y los valores de Hardy-Weinberg de los tres genotipos analizados, en tanto que en el cuadro 3 se encuentran agrupados los diferentes haplotipos hallados, con sus respectivas frecuencias. El cuadro 4 muestra la variabilidad interétnica de los tres polimorfismos del gen DβH (31,33: catálogo de variación genética humana del proyecto 1000 Genomas). Los tres SNPs (rs1989787, rs1611115 y rs1108580) están en fuerte desequilibrio de ligamiento (D'=1,0 para todas las combinaciones), pero no son Tag SNP entre ellos (r2<0,8 para todas las combinaciones), es decir, no se representan entre sí (figura 1).

Discusión
La DβH es una enzima con funciones clave, ya que sintetiza norepinefrina a partir de dopamina y de este modo regula los tonos adrenérgico y dopaminérgico en el SNC y en el SNA. Para cada individuo la actividad enzimática de la DPH es relativamente estable a lo largo de la vida y los efectos de factores ambientales sobre la actividad de la enzima son pequeños, comparados con los factores genéticos: la heredabilidad da cuenta de entre el 80 y el 90% de la variación en la actividad de la enzima, tanto en el plasma como en el LCR (12, 29); este control genético sobre la enzima explica que en cualquier población las personas se pueden categorizar como de actividad enzimática normal, intermedia o baja.
Los desequilibrios de ligamiento encontrados entre los tres SNPs estudiados concuerdan con los resultados de otros estudios (9). Resulta interesante el hallazgo que ninguno de ellos es Tag SNP (figura 1) y por tanto los tres son insustituibles y sumarían sus efectos sobre la actividad de la enzima (34), lo que nos permite predecir que a juzgar por las distribuciones haplotípicas halladas en la muestra (cuadro 3), solamente el 10,5% de nuestra población tiene actividad normal de la enzima. Esta hipótesis de efectos alélicos aditivos que expliquen la alta variabilidad interpersonal en la actividad de la DBH y por ende la diversidad de síndromes en los cuales está implicada la enzima, solo puede confirmarse con estudios diseñados para explorar las correlaciones genotipo-fenotipo (34) com/sch. Pero la presencia y la prevalencia de los polimorfismos asociados con cambios en la actividad de la enzima, deben ser primero confirmadas en cada grupo étnico, razón por la cual los estudios como este constituyen el primer paso en la búsqueda de la influencia de los polimorfismos del gen DβH, en respuesta a fármacos y en una serie de desórdenes neuropsiquiátricos, potencialmente relacionados con la actividad de la enzima dopamina beta-hidroxilasa.
El perfil genético DβH de esta muestra, representativa de mestizos del Eje Cafetero, es similar al de otros grupos de mestizos latinoamericanos (cuadro 4). Además, si juntamos los cuatro grupos mestizos reportados en el cuadro, nos da una distribución alélica (rs1989787: 0,748; rs1611115: 0,729; rs1108580: 0,586) que se aproxima más a las de negros y caucásicos, precisamente dos de los tres grupos étnicos con los que el mestizo latinoamericano está emparentado genéticamente.
Agradecimientos
A la Fundación Universitaria Autónoma de las Américas y a la Universidad Tecnológica de Pereira, por haber financiado esta investigación. A los estudiantes de la Fundación Universitaria Autónoma de las Américas, que nos colaboraron como voluntarios.
Referencias
1. Cubells J, Zabetian C. Human genetics of plasma dopamine beta-hydroxylase activity: applications to research in psychiatry and neurology. Psychopharmacology 2004; 174 (4): 463-76. [ Links ]
2. Dudas B, Baker M, Rotoli G, Grignol G, Bohn M, Merchenthaler I. Distribution and morphology of the catecholaminergic neural elements in the human hypothalamus. Neuroscience 2010; 171 (1): 187-95. [ Links ]
3. Kapoor A, Shandilya M, Kundu S. Structural insight of dopamine P-hydroxylase, a drug target for complex traits, and functional significance of exonic single nucleotide polymorphisms. PloS one 2011; 6 (10): e 26509.
4. Usera P, Vincent S, Robertson D. Human phenotypes and animal knockout models of genetic autonomic disorders. Journal of biomedical science 2004; 11 (1): 4-10. [ Links ]
5. Wei J, Ramchand C, Hemmings G. Possible control of dopamine beta-hydroxylase via a codominant mechanism associated with the polymorphic (GT) n repeat at its gene locus in healthy individuals. Human genetics 1996; 99 (1):52-5. [ Links ]
6. Hui L, Zhang X, Huang XF, Fernandez F, Yu Y, et. a. The dopamine b-hydroxylase 19 bp insertion/deletion polymorphism was associated with first-episode but not medicated chronic schizophrenia. Psychiatric Research 2012; 46 (6):733-7. [ Links ]
7. Paclt I, Koudelova J, Pacltova D, Kopeckova M. Dopamine beta hydroxylase (DBH) plasma activity in childhood mental disorders. Neuro endocrinology letters 2009; 30 (5):604-9. [ Links ]
8. Segurado R, Bellgrove M, Manconi F, Gill M, Hawi Z. Epistasis between neurochemical gene polymorphisms and risk for ADHD. European Journal of Human Genetics 2011; 19 (5): 577-82. [ Links ]
9. Cubells J, Sun X, Li W, Bonsall R, McGrath J, Avramopoulos D, et al. Linkage analysis of plasma dopamine P-hydroxylase activity in families of patients with schizophrenia. Human genetics 2011; 130 (5): 635-43. [ Links ]
10. Wang G-X, Ma Y-H, Wang S-F, Ren G-F, Guo H. Association of dopaminergic/GABAergic genes with attention deficit hyperactivity disorder in children. Molecular Medicine Reports 2012; 6 (5):1093-8. [ Links ]
11. Ribasés M, Ramos-Quiroga J, Hervás A, Sánchez-Mora C, Bosch R, Bielsa A, et al. Candidate system analysis in ADHD: Evaluation of nine genes involved in dopaminergic neurotransmission identifies association with DRD1. World Journal of Biological Psychiatry 2012; 13 (4): 281-92. [ Links ]
12. Combarros O, Warden D, Hammond N, Cortina-Borja M, Belbin O, Lehmann M, et al. The dopamine beta-hydroxylase -1021C/T polymorphism is associated with the risk of Alzheimer's disease in the Epistasis Project. BMC medical genetics 2010; 11(1):162. [ Links ]
13. Punia S, Das M, Behari M, Mishra B, Sahani A, Govindappa S, et al. Role of polymorphisms in dopamine synthesis and metabolism genes and association of DBH haplotypes with Parkinson's disease among North Indians. Pharmacogenetics and genomics 2010; 20 (7):435-41. [ Links ]
14. Fernández F, Colson N, Quinlan S, MacMillan J, Lea R, Griffiths L. Association between migraine and a functional polymorphism at the dopamine beta-hydroxylase locus. Neurogenetics 2009; 10(3):199-208. [ Links ]
15. Ghosh J, Pradhan S, Mittal B. Identification of a novel ANKK1 and other dopaminergic (DRD2 and DBH) gene variants in migraine susceptibility. Neuromolecular medicine 2012; 15(1):61-73. [ Links ]
16. Grigorenko E, DeYoung C, Eastman M, Getchell M, Haeffel G, Klinteberg B, et al. Aggressive behavior, related conduct problems, and variation in genes affecting dopamine turnover. Aggressive behavior 2010; 36(3):158-76. [ Links ]
17. Brousse G, Vorspan F, Ksouda K, Bloch V, Peoc'h K, Laplanche J, et al. Could the interindividual variability in cocaine-induced psychotic effects influence the development of cocaine addiction?: Towards a new pharmacogenetic approach to addictions. Medical hypotheses 2010; 75(6):600-4. [ Links ]
18. Xie X, Xu L, Liu H, Chen W, Zhuang D, Zhang J, et al. Positive association between -1021TT genotype of dopamine beta hydroxylase gene and progressive behavior of injection heroin users. Neurosci Lett 2013. [ Links ]
19. Ella E, Sato N, Nishizawa D, Kageyama S, Yamada H, Kurabe N, et al. Association between dopamine beta hydroxylase rs5320 polymorphism and smoking behaviour in elderly Japanese. Journal of human genetics 2012;57(6):385-90. [ Links ]
20. Flak J, Ostrander M, Tasker J, Herman J. Chronic stress-induced neurotransmitter plasticity in the PVN. Journal of Comparative Neurology 2009; 517(2):156-65. [ Links ]
21. Kim Y CE, Doo M, Kim J, Kim C, Kim C, et al. Anti-stress effects of ginseng via down-regulation of tyrosine hydroxylase (TH) and dopamine P-hydroxylase (DBH) gene expression in immobilization-stressed rats and PC12 cells. Nutr Res Pract 2010; 4(4):270-5.
22. Radley J, Williams B, Sawchenko P. Noradrenergic innervation of the dorsal medial prefrontal cortex modulates hypothalamo-pituitary-adrenal responses to acute emotional stress. The Journal of Neuroscience 2008;28(22):5806-16. [ Links ]
23. Ting-Kuang Y, Ting-Chi Y, Chi-Feng W, Bing-Fu S, Hsueh-Jen T, Chien-Hua H, et al. Association of polymorphisms in genes involved in the dopaminergic pathway with blood pressure and uric acid levels in Chinese females. Journal of Neural Transmission 2010; 117 (12):1371-6. [ Links ]
24. Friese R, Schmid-Schõnbein G, O'Connor D. Systematic polymorphism discovery after genome-wide identification of potential susceptibility loci in a hereditary rodent model of human hypertension. Blood pressure 2011; 0 (4):222-31. [ Links ]
25. Yuqing C, Gen W, Fangwen R, Kuixing Z, Lei W, Rodriguez-Flores JL, et al. Human dopamine òeta-hydroxylase (DBH) regulatory polymorphism that influences enzymatic activity, autonomic function, and blood pressure. J Hypertens 2011; 28(1):76-86.
26. Mutschler J, Abbruzzese E, Witt SH, Dirican Gl, Nieratschker V, Frank J, et al. Functional polymorphism of the dopamine P-hydroxylase gene is associated with increased risk of disulfiram-induced adverse effects in alcohol-dependent patients. Journal of clinical psychopharmacology 2012; 32(4):578-80.
27. Kosten T, Wu G, Huang W, Harding M, Hamon S, Lappalainen J, et al. Pharmacogenetic randomized trial for cocaine abuse: disulfiram and dopamine P-hydroxylase. Biological Psychiatry 2013; 73(3):219-24.
28. Helmstaedter C, Mihov Y, Toliat M, Thiele H, Nuernberg P, Schoch S, et al. Genetic variation in dopaminergic activity is associated with the risk for psychiatric side effects of levetiracetam. Epilepsia 2013; 54(1):36-44. [ Links ]
29. Chen Y, Wen G, Rao F, Zhang K, Wang L, Rodriguez-Flores JL, et al. Human dopamine beta-hydroxylase (DBH) regulatory polymorphism that influences enzymatic activity, autonomic function, and blood pressure. Journal of hypertension 2010; 28(1):76-86. [ Links ]
30. Tang Y-l, Epstein M, Anderson G, Zabetian C, Cubells J. Genotypic and haplotypic associations of the DBH gene with plasma dopamine beta-hydroxylase activity in African Americans. European Journal of Human Genetics 2007; 15 (8):878-83. [ Links ]
31. Tang Y, Anderson G, Zabetian C, Kohnke M, Cubells J. Haplotype-controlled analysis of the association of a non-synonymous single nucleotide polymorphism at DBH (+ 1603C --> T) with plasma dopamine beta-hydroxylase activity. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics 2005; 139(1):88-90. [ Links ]
32. Bhaduri N, Mukhopadhyay K. Correlation of plasma dopamine beta-hydroxylase activity with polymorphisms in DBH gene: a study on Eastern Indian population. Cellular and molecular neurobiology 2008; 28 (3):343-50. [ Links ]
33. Ensembl Project. 1000 Genomes Catalog of Human Genetic Variation. Cambridge, United Kingdom2013 [04/2013]; Available from: http://www.ensembl.org/Homo_sapiens/Gene/Summary?g=ENSG00000123454;r=9:136501482-136524466. [ Links ]
34. Zabetian C, Buxbaum S, Elston R, Kõhnke M, Anderson G, Gelernter J, et al. The structure of linkage disequilibrium at the DBH locus strongly influences the magnitude of association between diallelic markers and plasma dopamine beta-hydroxylase activity. The American Journal of Human Genetics 2003; 72(6):1389-400. [ Links ]

