










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.58 no.2 Medellín July/Dec. 2005
NET PHOTOSYNTHESIS AND CO2 COMPENSATION CONCENTRATION IN THREE COFFEE (Coffea sp.) GENOTYPES, BEAN AND MAIZE UNDER THREE TEMPERATURES
FOTOSÍNTESIS NETA Y CONCENTRACIÓN DE COMPENSACIÓN DE CO2 EN TRES GENOTIPOS DE CAFÉ (Coffea sp.), FRÍJOL Y MAÍZ BAJO TRES TEMPERATURAS
Lyda Patricia Mosquera Sánchez1; Néstor Miguel Riaño Herrera2; Yamel López Forero3 y Jaime Arcila Pulgarín4
1 Departamento de Biología, Unidad de Microscopia Electrónica. Universidad del Cauca. Popayán, Cauca, Colombia. <lmosquera@ucauca.edu.co>
2 Fisiología Vegetal, Centro Nacional de Investigaciones de Café. CENICAFÉ, Chinchiná, Caldas, Colombia. <nestorm.riano@cafedecolombia.com>
3 Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira, Colombia. <yamel@telesat.com.co>
4 Fitotecnia, Centro Nacional de Investigaciones de Café. CENICAFÉ, Chinchiná, Caldas, Colombia. <cenicafe@cafedecolombia.com>
Recibido: Junio 1 de 2005; aceptado: Octubre 10 de 2005.
ABSTRACT
The coffee (Coffea arabica L.) genotypes Colombia, Caturra, and Híbrido de Timor, and bean (Phaseolus vulgaris L.) and maize (Zea mays L.) plants were exposed to three temperatures (15°C, 25°C and 35°C), and net photosynthetic rates (PN) and CO2 compensation concentrations (GCO2) were measured. PN in coffee leaves was similar for the three genotypes at 15 °C [5,0 – 5,3 µmol(CO2) m-2s-1] and 35 °C [4,9 – 5,5 µmol(CO2) m-2s-1], but lower at 25 °C [5,4 – 11,7 µmol(CO2) m-2s-1]. GCO2 increased with temperature in coffee and bean, while in maize no effect was observed. PN and GCO2 values documented in coffee genotypes were typical for C3 plants.
Key words: Coffee, Coffea arabica L., CO2 compensation concentration, leaf temperature, net photosynthesis, bean, maize.
RESUMEN
Se expusieron plantas de café Coffea arabica L. de los genotipos Colombia, Caturra e Híbrido de Timor, fríjol (Phaseolus vulgaris L.), maíz (Zea mays L.), a tres temperaturas (15 °C, 25 °C y 35 °C); se midió la fotosíntesis neta (PN) y se obtuvo la concentración de compensación de CO2 (GCO2). PN en las hojas de café fueron similares para los tres genotipos a 15 °C [5,0 – 5,3 µmol(CO2) m-2s-1] y 35 °C [4.9 – 5,5 µmol(CO2) m-2s-1] pero más bajas a 25 °C [5,4 – 11,7 µmol(CO2) m-2s-1]. El GCO2 en café y fríjol se incrementó con la temperatura, mientras en maíz no se presentó ningún efecto. Los valores observados PN y de GCO2, en los genotipos de café fueron los típicos de plantas C3.
Palabras clave: Café, Coffea arabica L., concentración de compensación de CO2, temperatura de la hoja, fotosíntesis neta, fríjol, maíz.
Abbreviations: ATP – adenosine triphosphate; NADH – nicotinamide adenine dinucleotide reduced form; PAR – photosynthetic active radiation; Pi – inorganic phosphate; PN – net photosynthesis; RuDP – Ribulose 1,5-diphosphate; VPD – vapour pressure deficit; GCO2 – CO2 compensation concentration.
 MATERIALS AND METHODS
MATERIALS AND METHODS
RESULTS AND DISCUSSION
REFERENCES
Continuous increase in concentration of CO2 and other atmospheric gases are contributing to the rise of global air temperature, causing variations in plant growth (Kimball et al.,1993; Taylor et al., 1994).
The effect of temperature on photosynthesis depends on plant species and environmental conditions under which the plant is growing (Kimball et al., 1993; Larcher, 1995). In general, optimum temperatures for photosynthesis in C3 plants with high GCO2 are between 20 °C and 25 °C, while for C4 plants optimum temperature is between 30 °C and 40 °C (Fitter and Hay, 1987; Kimball et al., 1993). GCO2 has been used to estimate photosynthetic efficiency: In C4 plants this value is near to zero, which makes them photosynthetically more efficient than C3 plants, which have GCO2 values above [30 µmol(CO2)mol-1(air)] (Kennedy 1976, Ogren, 1984, Kimball et al., 1993; Taylor et al., 1994).
In coffee, the study of factors that influence photosynthesis started with the works of Nutman (1937), who established that PN is higher under low light intensities and leaf temperatures because stomatal opening is not affected. Nunes et al. (1968) and Kumar and Tieszen (1980), confirmed Nutmans observations and found that optimum temperature for photosynthetic activity is between 20 °C and 25 °C.
Photosynthesis in C3 plants is limited by photorespiration and most of this activity is closely related to temperature (Zelitch, 1971; Long, 1991 and Pastenes, 1996). Pioneer works of Decker and Tio (1959) showed that photosynthesis in coffee leaves were cancelled almost immediately by respiration in the presence of light (photorespiration) and only a small fraction was left as net gain. Heath and Orchard (1957), Jones and Mansfield (1970), and Sondhal (1976) studied GCO2 as an estimate for photosynthesis efficiency involving photorespiration, and found GCO2 values close to [65 µmol(CO2)mol-1(air)] at 25°C.
The objective of this research was to study the behavior of PN and GCO2, under different temperatures, in three coffee (Coffea arabica L.) genotypes, bean (Phaseolus vulgaris L.), and maize (Zea mays L.), under conditions of the Colombian central coffee-growing zone.
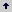
This research was performed at the Centro Nacional de Investigaciones de Café CENICAFE, Chinchiná, Colombia, 5°01 N, 75°36 W, altitude of 1425 m, mean annual temperature of 20,6 °C, 2530 mm cumulative rainfall and 1830 hours cumulative sunshine.
8 months Coffea arabica L. cv. Caturra, Colombia , and interspecific natural hybrid Híbrido de Timor - plants; bean (Phaseolus vulgaris L. cv. Ica Cafetero PVA 916) and maize (Zea mays L. cv. ICA V305) plants were 45 days old. All were planted in individual pots with soil and decomposed coffee fruit pericarp (3:1), and growing under full exposition to sunlight. Net photosynthesis rate (PN) was measured with a portable photosynthesis system (model 6200, LI-COR, Lincoln, NE, U.S.A. ), infrared gas analyzer (IRGA), data logger, and a 4 L Plexiglas leaf chamber. PN was measured on two mature leaves of the higher part of five plants per genotype, between 08:00 hours and 12:00 hours, with photosynthetic active radiation (PAR) above [600 µmol(photon) m-2 s-1], measured with a quantum sensor (model LI-190S-1, LI-COR, Lincoln, NE, U.S.A.). Air and leaf temperatures inside the chamber (15 °C, 25 °C, and 35 °C) were maintained with a JULABO-J10 recirculation water system. Measurement started with air containing [450 µmol(CO2) mol-1(air)] and PN was recorded for every [50 µmol(CO2) mol-1(air)] decrease inside the leaf chamber until a constant CO2 concentration (GCO2) was reached.
The highest PN observed for the three genotypes [11,7 µmol(CO2)m-2 s-1] was reached at 25°C and [350 µmol(CO2) mol-1(air)] (Figure 1a), with values higher than those reported by Nutman (1937) [0,44 - 2,84 µmol(CO2) m-2s-1] at 25°C; Nunes et al., (1968) [3,51 - 4,41 µmol(CO2) m-2 s1] at 24°C; Sondhal (1976) [3,87 µmol(CO2) m-2s1] at 20°C; Yamaguchi and Friend (1979) [3,5 µmol(CO2) m-2s1] at 25°C and Friend (1984) [4,4 µmol(CO2) m-2s1]. Temperature influenced PN directly, when measured at [350 µmol(CO2) mol-1(air)] (Table 1), values obtained at 15 °C and 35 °C were similar, but lower than those at 25 °C. At 15 °C, PN would be limited by RuDP regeneration, because electronic transport velocity diminishes and consequently ATP and NADH formation are limited, and Pi regeneration capacity during starch and sucrose synthesis is lowered, as well as carbohydrate transport from chloroplasts (Azcon-Bieto, 1983; Makino, Nakano and Mae, 1994). Another limitation is Rubisco activity because its optimum activation temperature in coffee is between 25 °C and 28 °C (Riaño and López, 1998), similar to what has been reported for other perennial species (Woodrow and Berry, 1988). At 35 °C, decrease in PN would be related to stomatal closure, leaf sensitivity to increase in water vapor pressure deficit (VPD) as reported by Hernández, Cock and E-Sharkawy (1989), and the increase of photorespiration due to increase of oxygenase activity on RuBPCO, caused by higher diffusion of O2 to the active site, which causes an additional CO2 loss (Zelitch, 1971; Ogren, 1984; Lawlor, 1987; Poorter, 1993; Pearson and Brooks, 1995; Pastenes and Horton, 1996a,b), thus lowering photosynthetic efficiency. Additionally, temperatures above the optimum reduce RuBPCO content, as reported by Vu et al., (1997) in soybean and rice.
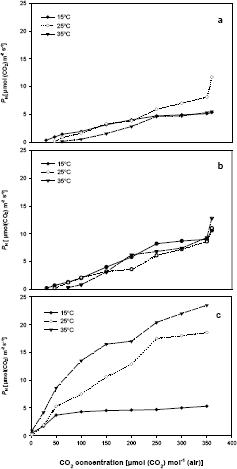
Figure 1. Net photosynthesis (PN) under 15 °C (·), 25 °C (¡), 35 °C (Ñ) and different CO2 concentrations in (a) coffee (Coffea arabica L.) cv. Caturra, (b) bean (Phaseolus vulgaris L.) (c) maize (Zea mays L).
Table 1. Net photosynthesis (PN) of Coffea sp genotypes, bean and maize under three temperatures and [350 µmol(CO2) mol-1(air)]. Means ± SD of ten measurements.

In bean plants (Figure 1b), PN increased slightly with temperature variation despite being considered a C3 plant. The genotype used, ICA Cafetero PVA 916, adapted to the Colombian central coffee-growing zone conditions, [1000 - 2000 m of altitude], accounts for this behavior, even though GCO2 was within the range for C3 plants (Table 2).
Table 2. CO2 compensation point (GCO2) in Coffea arabica L. cvs. Caturra and Colombia, Híbrido de Timor, bean and maize, for 15 °C, 25 °C, and 35 °C. Means ± SD of ten measurements.
Maize (C4 photosynthetic metabolism), showed an increase in PN with temperature increase, reaching highest values at 35 °C (Figure 1c). High temperatures induced higher activity of phosphoenol pyruvate carboxylase (PEPC) in this species (Tolbert,1980; Ogren, 1984).
Two groups, according to photosynthetic activity are formed (Figure 2) for 25 °C and several CO2 concentrations. The first group is conformed by coffee and bean with PN between [5,4 and 13 µmol(CO2) m-2s-1], and the second group is represented by maize with higher rates [18,6 µmol(CO2) m-2 s-1].
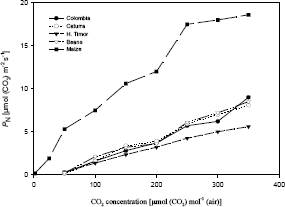
Figure 2. Net photosynthesis (PN) of coffee (Coffea arabica L.) cv. Caturra (¡), Colombia (l), Híbrido de Timor (Ñ), bean (Phaseolus vulgaris L.) (Ñ) and maize (Zea mays L.) () at 25 °C and several CO2 concentrations [µmol (CO2) mol-1(air)].
For the species studied, the lower the CO2 concentration at the three temperatures (15 °C, 25 °C, and 35 °C), the lower the PN, until CO2 compensation point is reached, where photosynthesis rate is equal to CO2 release by photorespiration and mitochondrial respiration (Espie and Colman, 1987). In coffee and bean, GCO2 increased with temperature, while in maize no changes were observed (Table 2). This increase in GCO2 with higher temperatures in coffee and bean indicates the occurrence of photorespiration, not detected in maize (Zelitch, 1971; Canvin,1979; Tolbert, 1980; Ogren, 1984).
Highest GCO2 value at 25 °C in coffee was observed with Híbrido de Timor [80,1 ± 1,9 µmol(CO2) mol-1(air)], suggesting the occurrence of a higher photorespiration rate in this genotype, followed by Caturra [45,6 ± 1,3 µmol(CO2) mol-1(air)] and Colombia [40,6 ± 1,0 µmol(CO2) mol-1(air)]. For the last two genotypes, GCO2 values are lower than those found by Jones and Mansfield (1970) [85 µmol(CO2) mol-1(air)] and Sondhal (1976) [65 µmol(CO2) mol-1 (air)], at 23 °C and 25 °C, respectively. GCO2 at 25 °C in bean plants [36,05 ± 0,5 µmol(CO2) mol-1(air)], and maize [0,8 ± 0,3 µmol(CO2) mol-1(air)] are similar to the values presented by Zelitch (1971).
In Coffea arabica L. cv. Colombia and bean, 60-65 minutes were necessary to reach GCO2, while in maize only 25 min were needed (Figure 3). Comparison of times necessary to reach GCO2 in C3 and C4 plants is an evidence of the level of competition caused by photorespiration in C3 plants, which is poorly reported in literature. Under the same temperature and initial CO2 concentration in a closed system, more time is required by C3 plants due to significant return of the CO2 fixed to the environment by photorespiration (Zelitch, 1971; Tolbert, 1980; Ogren, 1984). In maize, a C4 plant, where CO2 lost by photorespiration is recycled; time necessary to reach GCO2 is shorter, allowing higher net CO2 input rates than those in C3 plants (Canvin, 1979).
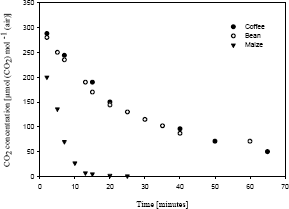
Figure 3. Time required for coffee Coffea arabica L. cv. Caturra (·), bean Phaseolus vulgaris L. (¡) and maize Zea mays L. (Ñ) to reach photosynthesis and (dark respiration + photorespiration) balance [CO2 compensation point (GCO2)] at 25 °C.
Results indicate that in coffee plants, similar to what happens with other C3 plants, GCO2 is dependent on temperature and that there is a wide range of variation in photosynthetic activity directly related to photorespiratory competition. The study of different genotypes of Coffea arabica L., species and Coffea genus would allow identification of photosynthetically efficient genotypes to be used in breeding programs in order to increase productivity. Photosynthetic behavior of the plants studied was that of a typical C3 plant, and optimum leaf temperature for photosynthesis under conditions of the Colombian central coffee-growing zone is around 25 °C.
AZCON – BIETO, J. Inhibition of photosynthesis by carbohydrates in wheat leaves. In: Plant Physiology. Vol. 73 (1983); p. 681-686. [ Links ]
CANVIN, D.T. Photorespiration: comparison between C3 and C4 plants. In: GIBBS, M., and LATZKO, E., eds. Encyclopedia of Plant Physiology. Berlin: Springer Verlag, 1979. p. 368-398. [ Links ]
DECKER, J. P. and TIO, M. A. Photosynthetic surges in Coffee seedlings. In: Journal of Agriculture of the University ofPuerto Rico. Vol. 43 (1959); p. 50-55. [ Links ]
ESPIE, G. S., COLMAN, B. The effect of pH, CO2 and temperature on the CO2 compensation point of isolated Asparagus mesophyll cells. In: Plant Physiology. Vol. 83 (1987); p. 113-117. [ Links ]
FITTER, A. H. and HAY, R. K. M. Environmental physiology of plants. London: Academic Press, 1987. 367 p. [ Links ]
FRIEND, D. J. C. Shade adaptation of photosynthesis in Coffea arabica L. In: Photosynthesis Research. Vol. 5 (1984); p. 325-334. [ Links ]
HEATH, O. V. S. and ORCHARD, B. Midday closure of stomata. Temperature effects on the minimum intercellular space carbon dioxide concentration G. In: Nature. Vol. 180 (1957); : p. 180-182. [ Links ]
HERNANDEZ, A. Del P.; COCK, J. H. and EL-SHARKAWY, M. A. The responses of leaves gas exchange and stomatal conductance to air humidity in shade grown coffee, tea and cacao plants as compared with sunflower. In: Revista Brasileira de Fisiología Vegetal. Vol. 1 (1989); p. 155-161. [ Links ]
JONES, M. B., Mansfield, T. A. A circadian rhythm in the level of the carbon dioxide compensation point in Briophyllum and Coffea. In: Journal of Experimental Botany. Vol. 21 (1970); p. 159-163. [ Links ]
KENNEDY, R. A. Photorespiration in C3 and C4 plant tissue cultures. In: Plant Physiology. Vol. 58 (1976); p. 573 – 575. [ Links ]
KIMBALL, A. et al. Effects of increasing atmospheric CO2 on vegetation. In: ROZEMA, J. et al., eds. CO2 and biosphere. The Netherlands: Kluwer, Dordrecht, 1993. p. 65-75. [ Links ]
KUMAR, D. and TIESZEN, L. D. Photosynthesis in Coffea arabica L. Part 1. Effects of light and temperature. In: Experimental Agriculture. Vol. 16 (1980); p. 13-19. [ Links ]
LARCHER, W. Photosynthesis as a tool for indicating temperature stress events. In: SCHULZE, E. D. and CALDWELL, M. M., eds. Ecophysiology of photosynyhesis. New York: Springer Verlag, 1995. p. 261-274. [ Links ]
LAWLOR, D. W. Photosynthesis: metabolism control and physiology. New York: Longman Scientific and Technical, 1987, p. 183-201. [ Links ]
LONG, S. P. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations: Has its importance been underestimated? In: Plant Cell and Environment. Vol. 14 (1991); p. 739-736. [ Links ]
MAKINO, A.; NAKANO, H. and MAE, T. Effects of growth temperature on the response of ribulose- 1,5 - bisphosphate carboxylase, electron transport components and sucrose synthesis enzymes to leaf nitrogen in rice and their relationships to photosynthesis. In: Plant Physiology. Vol. 105 (1994); p. 1231-1238. [ Links ]
NUNES, M. A.; BIERHUIZEN, J. F. and PLOEGMAN, C. Effect of light, temperature and CO2 concentration on photosynthesis of Coffea arabica L. In: Acta Botanica Neerlandica. Vol. 17 (1968); p. 93-102. [ Links ]
NUTMAN, F. J. Studies of the physiology of Coffea arabica: photosynthesis of coffee leaves under natural conditions. In: Annals of Botany, London. Vol. 1, No. 3 (1937); p. 353-367. [ Links ]
OGREN, W. L. Photorespiration: pathways, regulation and modification. In: Annual Review of Plant Physiology. Vol. 35 (1984); p. 415-442. [ Links ]
PASTENES, C. and HORTON, P. Effect of high temperature on photosynthesis in beans. Part 1: oxygen evolution and chlorophyll fluorescence. In: Plant Physiology. Vol.112 (1996); p. 1245-1251. [ Links ]
________ and ________. Effect of high temperature on photosynthesis in beans. Part 2: CO2 assimilation and metabolite contents. In: Plant Physiology. Vol. 112 (1996); p. 1253-1260. [ Links ]
PEARSON, M. and BROOKS, G. The influence of the elevated CO2 on growth and age -related changes in leaf gas exchange. In: Journal of Experimental Botany. Vol. 46 (1995); p. 1651-1659. [ Links ]
POORTER, H. Interspecific variation in the growth response of plants to an elevated ambient CO2 concentration. In: ROZEMA, J. et al., eds. CO2 and biosphere. The Netherlands: Kluwer, Dordrecht, 1993. p. 677-697. [ Links ]
RIAÑO, N. M. y LÓPEZ, Y. Cuantificación de RuBPCO en hojas y frutos de café. En: OSPINA, H. F., ed. Informe anual de labores. Chinchiná, Colombia : Cenicafé, 1998. p. 95- 97. [ Links ]
SONDHAL, M. R. Measurements of 14C incorporation by illuminated intact leaves of coffee plants from gas mixtures containing 14CO2. In: Journal of Experimental Botany. Vol. 27 (1976); p. 1187-1195. [ Links ]
TAYLOR, G. et al. Elevated CO2 and plant growth: celular mechanisms and responses of whole plants. In: Journal of Experimental Botany. Vol. 45 (1994); p. 1761-1774. [ Links ]
TOLBERT, N. E. Glicolate metabolism by higher plants and algae. In: GIBBS, M. and LATZKO, E., ed. Encyclopedie of Plant Physiology. Berlin: Springer Verlag, 1980. p. 338-351. [ Links ]
VU, J. C. V. et al. Effects of elevated CO2 and temperature on photosynthesis and rubisco in rice and soybean. In: Plant Cell and Environment. Vol. 20 (1997); p. 68-76. [ Links ]
WOODROW, I. E. and BERRY, J. A. Enzymatic regulation of photosynthetic CO2 fixation in C3 plants. In: Annual Review of. Plant Physiology. Vol. 39 (1988); p. 533-594. [ Links ]
YAMAGUCHI, D. and FRIEND, D. J. C. Effect of leaf and irradiance on photosynthesis of Coffea arabica L. In: Photosynthetica. Vol. 13 (1979); p. 271-278. [ Links ]
ZELITCH, I. Photosynthesis, photorespiration and plant productivity. New York: Academic Press, 1971. 347 p. [ Links ]

