










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.58 no.2 Medellín July/Dec. 2005
EVALUACIÓN DE DOS TRATAMIENTOS FOTOPERIÓDICOS EN CRISANTEMO (Dendranthema grandiflorum (Ramat.) Kitam.), BAJO CONDICIONES DEL INTERTRÓPICO ANDINO ALTO
EVALUATION OF TWO PHOTOPERIOD TREATMENTS IN CHRYSANTHEMUM (Dendranthema grandiflorum ((Ramat.) Kitam.), UNDER CONDITIONS OF HIGH ANDEAN INTERTROPIC
Fáber de Jesús Chica Toro1 y Guillermo Antonio Correa Londoño2
1 Ingeniero Agrónomo. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia.<fchica@unalmed.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <gcorrea@unalmed.edu.co>
Recibido: Septiembre 24 de 2004, aceptado: Julio 22 de 2005.
RESUMEN
En la floricultura colombiana, cerca del 14 % de los costos de producción del crisantemo o pompón (Dendranthema grandiflorum (Ramat.) Kitam.), está representado por la luz complementaria requerida para controlar su floración. Con el fin de encontrar un protocolo que hiciera uso de un menor suministro de luz, en este trabajo se compararon los efectos de dos esquemas de complemento lumínico nocturno sobre algunas características precosecha y poscosecha en dos variedades comerciales tipo spider de crisantemo: Super White y Super Yellow. El esquema testigo de suplemento lumínico nocturno consistió en aportar la adición durante 28 días para la variedad Super Yellow, y 26 para la variedad Super White. La propuesta alternativa de adición lumínica nocturna consistió en interrumpir el estímulo durante una noche completa cada dos días. Tal disminución del estímulo lumínico, en aproximadamente un 30 %, no afectó el crecimiento ni las características poscosecha de las variedades evaluadas.
Palabras claves: Fotoperíodo, Dendranthema grandiflorum, crisantemo, pompón.
ABSTRACT
In Colombian floriculture, nearly 14% of the production costs of chrysanthemum (Dendranthema grandiflorum (Ramat.) Kitam.) consists of the additional lighting required to control its flowering. In order to find a protocol that consumes a lower amount of electricity, this study compared the effects of two supplementary noctural lighting schemes on several preharvest and postharvest characteristics in two commercial varieties of chrysanthemum: Super White and Super Yellow. The control nocturnal lighting scheme consisted of providing the supplemental lighting during 28 days for the Super Yellow variety and during 26 days for the Super White variety. The alternative nocturnal lighting scheme consisted of interrupting the stimulus during one complete night every two days. This approximately 30% reduction in light stimulation did not affect either growth or postharvest characteristics of the varieties evaluated.
Key words: Photoperiod, Dendranthema grandiflorum, chrysanthemum.
 MATERIALES Y MÉTODOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
RECOMENDACIONES
BIBLIOGRAFÍA
Un significativo componente de los costos de producción del sector florícola está representado por la luz complementaria requerida para controlar la floración en algunas especies, entre ellas el crisantemo o pompón (Dendranthema grandiflorum (Ramat.) Kitam.). Las adiciones lumínicas nocturnas usadas actualmente en el trópico para ampliar el fotoperíodo se derivan de las aplicadas en la zona templada; sin que se haya desarrollado un modelo de manejo de la luz a partir de las condiciones particulares del trópico. De optimizarse su uso, el sector florícola colombiano se vería notablemente favorecido, al disminuirse los costos de producción por concepto del suministro de energía lumínica nocturna.
Langhans (1964) define el fotoperíodo como el número de horas de luz por día. Para Kofranek (1992), el día dura mientras exista una intensidad lumínica solar de 15 bujías-pie. Luego, puede definirse el fotoperíodo como el número de horas por día en que la intensidad lumínica es mayor o igual que 15 bujías-pie. En términos fisiológicos, 15 bujías-pie equivalen a 0,6456 wm-2 (Cockshull, 1987; citado por Thomas y Vince-Prue, 1997).[A1] [A2]
Entre las especies sensibles al fotoperíodo respecto al fenómeno de inducción floral, pueden distinguirse plantas de día corto, en las que la floración se induce como consecuencia de la exposición a días cortos; plantas de día largo en las que la floración se induce por exposición a días largos; y plantas neutras, en las que el proceso de inducción floral no es afectado por la duración del día (Wareing y Phillips, 1986).
Fides (1990) clasifica al crisantemo como una planta de día corto, que inicia la floración cuando el fotoperíodo es inferior a 14,5 horas; éste es su fotoperíodo crítico inductivo (FCI). Esto significa que para mantener su condición vegetativa requiere un fotoperíodo mayor de 14,5 horas.
En las condiciones del Oriente Cercano Antioqueño, a unos 6° de latitud Norte, la longitud del día oscila entre 11,65 h (diciembre 21: solsticio de invierno) y 12,35 h (junio 21: solsticio de verano) (Bustamante, 1989). Así, para producir el efecto de día largo (DL), que se requiere para mantener la condición vegetativa de la planta, sería necesario prolongar artificialmente la luz del día entre 2,5 y 3 horas, según el faltante para superar el FCI.
En algunos trabajos que sobre esta especie realizó Cathey (1969), citado por Vince-Prue y Cockshull (1981), los tratamientos de suspensión nocturna (iluminación con lámparas de tungsteno) aparentemente resultaron efectivos en retardar la floración. Acorde con Kofranek (1992), la luz complementaria debe ser suministrada en la mitad del período oscuro, sin que éste exceda las cinco horas continuas.
Se observa que la luz puede darse intermitentemente a través de varios periodos durante la noche, sin que aparentemente se presente una pérdida de su efecto inhibidor de la floración. En las empresas floricultoras del Oriente Antioqueño (Antioquia, Colombia), el protocolo de complemento para Dendranthema grandiflorum (Ramat.) Kitam no está unificado. Se encuentran diferentes duraciones, tanto de los periodos afóticos, como del tiempo de suministro lumínico artificial. Se tienen, por ejemplo, adiciones lumínicas desde una hasta cinco horas continuas de luz. No obstante, ninguno de los protocolos utilizados por las empresas dedicadas a esta actividad ha sido generado con base en investigaciones formales, por lo menos con publicaciones al respecto. De todos los protocolos de adición lumínico nocturno implementados por los floricultores de la zona, el más común consiste en suministrar 5 minutos de luz artificial por cada 25 minutos de oscuridad, hasta sumar 80 minutos de adición lumínica.
El uso de genotipos con una respuesta fotoperiódica pronunciada, en combinación con un adecuado control fotoperiódico constituye una poderosa herramienta para regular la producción de la cosecha y sus características, siendo posible, por ejemplo, concentrar la recolección en un periodo corto y de gran interés para el cultivador. Con el suministro de luz adicional también se promueve el desarrollo vegetativo de la planta, con lo cual el tallo logra alcanzar la longitud necesaria para cumplir con los estándares internacionales de comercialización. Así, Karaguzel et al. (2004), al evaluar diferentes duraciones del fotoperíodo en Lupinus varius, encontraron que los fotoperíodos que suministraron mayor cantidad de luz incrementaron la longitud del tallo en comparación con el tratamiento de luz natural.
El crisantemo es una de las especies con alta respuesta al fotoperíodo, lo que permite prolongar su estado vegetativo, mediante el suministro de días largos, obteniéndose, por tanto, flores con un tallo largo y frondoso (Vince-Prue y Cockshull, 1981).
En algunos trabajos pioneros con crisantemo en el Reino Unido, realizados por Vince– Prue (1953), citado por Vince-Prue y Cockshull (1981), se logró retardar la floración, mediante el suministro de días largos. Hoy en día las flores del crisantemo se producen en cada semana del año dándole a cada lote de siembra el tratamiento fotoperiódico apropiado. El mismo autor informa sobre un incremento de hasta un 300 % (para el Reino Unido) en la productividad, a causa del manejo fotoperiódico de la especie.
Los cortes energéticos a los que se vio sometida Colombia en el año 1992 redundaron en la disminución de la adición lumínica nocturna del crisantemo, lo cual, en términos generales, no ocasionó mayores trastornos fisiológicos en esta especie, ni pérdidas económicas al cultivo por concepto de su manejo.
Tal circunstancia revivió el debate acerca de las verdaderas necesidades del complemento lumínico nocturno para el crisantemo en el trópico. Se iniciaron desde entonces algunos ensayos tendientes al ahorro energético, todos ellos sin publicar; en tal sentido, este trabajo es pionero en el área, al menos para Colombia. Vale la pena destacar el trabajo realizado por Arango (1995) en La Ceja, Antioquia, Colombia (6º de latitud Norte), en el que se evaluó un programa consistente en suministrar dos noches de adición lumínica nocturna, seguidas de una noche sin el complemento. En tal ensayo no se detectaron diferencias con el programa convencional ininterrumpido.
El objetivo del presente trabajo fue evaluar y proponer, para el Oriente Cercano Antioqueño (trópico andino alto), un protocolo para el manejo del estímulo lumínico nocturno en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), en las variedades comerciales tipo spider: Super White y Super Yellow, haciendo uso de una menor cantidad del recurso luz. Para ello se comparó el efecto del actual esquema de prolongación del estímulo lumínico nocturno con uno de menor duración, sobre las características poscosecha de las variedades. Además se comparó el crecimiento y desarrollo entre el actual esquema de complemento lumínico nocturno y uno de menor duración.
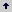
Localización.
El trabajo se ejecutó en el cultivo de flores C. I. Jardines del Chaparral S.A., ubicado en el municipio de La Ceja (Oriente Cercano Antioqueño), con una temperatura de 18° C, humedad relativa del 78 %, precipitación promedio anual de 2.000 mm y 2.150 msnm, que, según Espinal (1992), pertenece a la zona de vida bosque húmedo montano bajo (bh–MB). Las pruebas de vida en florero se realizaron en el campus de la Universidad Nacional de Colombia, Sede Medellín.
Metodología.
Se usó una metodología que permitiera comparar el efecto de dos esquemas de adición lumínica nocturna (niveles de luz), sobre el crecimiento y desarrollo, y sobre las características poscosecha de Dendranthema grandiflorum (Ramat.) Kitam., variedades comerciales tipo spider: Super White y Super Yellow.
Cada nivel de luz se asignó a un grupo de cuatro camas contiguas, sembrando dos de ellas con cada una de las variedades. Los niveles de luz se aislaron mediante cortinas plásticas oscuras. El conjunto de ocho camas que reciben los dos niveles de luz constituye una repetición (Figura 1). El experimento se replicó 12 veces, sembrando semanalmente un bloque completo de ocho camas durante 12 semanas. La unidad experimental para los niveles de luz constó de cuatro camas; mientras que la unidad experimental para las variedades estuvo constituida por dos camas. Las dimensiones de la cama son de 1,2 m de ancho, por aproximadamente 30 m de largo. En cada una se sembraron alrededor de 2.400 esquejes.
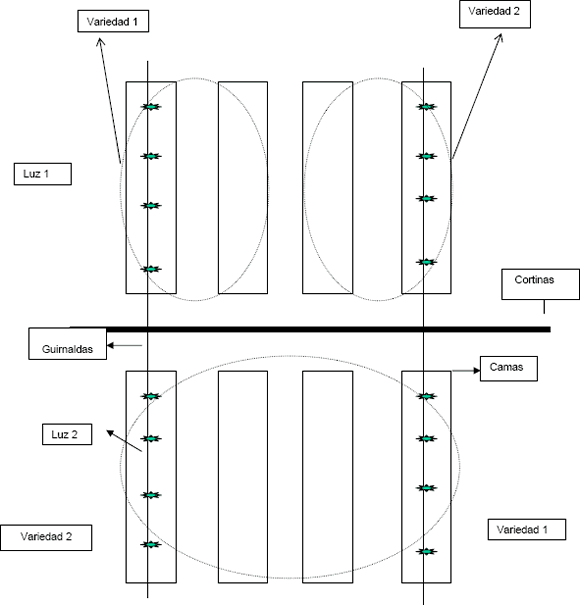
Figura 1. Esquema de aleatorización de los tratamientos fotoperiódicos en crisantemo. La parte superior (luz 1) y la parte inferior (luz 2) constituyen las parcelas principales, dentro de las cuales están las subparcelas (variedad 1 y 2), cada una de ellas constituida por dos camas.
Se consideraron dos niveles del factor luz: el primer nivel (L1) fue el testigo, consistente en días largos ininterrumpidos, que oscilaron entre 13,36 y 12,98 horas; el segundo (L2) consistió en días largos interrumpidos. Así mismo, se evaluaron dos niveles del factor variedad: Super White (SW) y Super Yellow (SY). Luego, los tratamientos estuvieron conformados por las cuatro combinaciones de los niveles de los factores luz y variedad, así: L1-SW, L1-SY, L2-SW, L2-SY.
El suministro del estímulo lumínico nocturno tradicional (L1) se aplicó con base en el siguiente programa: aporte de cinco minutos de luz por 25 minutos de oscuridad, a partir de las 21:15 horas y hasta las 04:50 horas (80 minutos de complemento lumínico). Este programa se aplicó durante 28 días para la variedad SY y 26 para la variedad SW, conservando el programa lumínico aplicado por el cultivo, lo cual garantiza obtener la floración en el mismo momento, dadas las diferencias genéticas entre las dos variedades. El segundo nivel del factor luz (L2) consistió en interrumpir el estímulo cada dos días. Los ciclos fueron de dos días con estímulo lumínico nocturno, siguiendo el esquema anteriormente descrito (días largos í) y uno sin el estímulo (día corto ‚) (Figura 2). Se usaron 11 bombillas de 200 vatios por cada tres camas.

Figura 2. Esquema de aplicación del suministro lumínico nocturno en crisantemo. Bajo el tratamiento propuesto (L2) se elimina cada dos días el complemento lumínico nocturno (‚). Bajo el tratamiento testigo (L1) se suministra complemento lumínico (í) ininterrumpidamente.
Variables respuesta.
Las variables cuya evolución se analizó a través del tiempo (diámetro y longitud del tallo, área foliar y masa seca) se midieron sobre tres plantas por cama, esto es, seis plantas por unidad experimental para variedad, 12 por unidad experimental para luz y 24 por repetición, para un total de 288 plantas por semana. Las variables que se midieron una única vez al final de cada corte (diámetro del capítulo, longitud y peso final del tallo y vida en florero) se evaluaron sobre 30 tallos por cama, esto es, 60 tallos por unidad experimental para variedad, 120 tallos por unidad experimental para luz y 240 tallos por repetición, para un total de 2.880 plantas.
Precosecha.
La información de campo se tomó cada siete días, desde el transplante del material vegetal a las camas, hasta la cosecha. El muestreo se basó en un esquema al azar restringido. La restricción consistió en no incluir aquellas plantas adyacentes a un sitio muestreado previamente, pues éstas podrían presentar un desarrollo diferente con relación a aquéllas cuyas vecinas no hubiesen sido retiradas. Mediante esta restricción se evita sobreestimar el efecto natural de borde. En esta etapa se evaluaron las siguientes variables:
No destructivas:
- Diámetro del tallo. Se evaluó semanalmente en tres plantas diferentes de cada cama, desde la tercera hasta la duodécima semana. La medición se realizó siempre a una altura de 10 cm.
- Diámetro del capítulo. Se midió a 30 flores de cada cama el día del full de corte, definido éste como el día en el que se ha cosechado el 50 % de las flores de cada cama.
- Longitud del tallo. Se evaluó semanalmente en tres plantas diferentes de cada cama, desde la tercera hasta la duodécima semana. Se midió la longitud desde el nivel del suelo hasta la inserción de la última hoja completamente abierta.
- Longitud final del tallo. Se midió a 30 tallos de cada cama al momento del full de corte, desde el nivel del suelo hasta la inserción del botón floral.
- Días a floración. Se midió para cada cama el tiempo transcurrido desde el transplante hasta el momento del full de corte.
Destructivas:
- Área foliar. Se evaluó semanalmente en tres plantas por cama, utilizando el método de la masa seca, reportado por Cartagena, Martínez y Hoyos (2001). Su lectura se realizó sólo hasta la novena semana, pues la labor cultural de desbotonado que ocurre durante la décima semana genera daños mecánicos en las hojas, alterando la dinámica de esta variable.
- Masa seca. Se registraron separadamente las masas secas de tallos y hojas, durante las nueve primeras semanas a tres plantas por cama. Se utilizó una estufa de secado a 80 ºC durante 72 horas, tiempo estimado para alcanzar masa constante.
Poscosecha.
Las variables evaluadas fueron:
- Masa final del tallo. Se evaluó con base en 30 tallos completos (tallo, hojas y flor) escogidos al azar en cada cama.
- Vida en florero. Se definió como el tiempo transcurrido para que la mitad de los tallos de la unidad experimental presentaran necrosis ascendente mayor o igual al 80 % de los dos tercios superiores del tallo (Figura 3). La unidad experimental constó de 60 tallos. Se utilizaron floreros separados para cada tallo con el fin de garantizar la independencia de las muestras. A cada tallo se le realizó un seguimiento durante 20 días o hasta que hubiese alcanzado una necrosis total. Las condiciones de evaluación fueron aproximadamente 75 % de humedad relativa y una temperatura promedio de 24 oC, y un fotoperíodo promedio de 12 y 11,58 horas para los primeros dos meses y para los segundos dos meses de la medición, respectivamente.
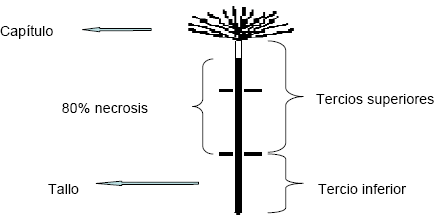
Figura 3. Esquema del estado de necrosis del tallo utilizado para la medición de la vida en florero de crisantemo sometido a tratamientos fotoperiódicos.
Método estadístico.
Usando regresiones no lineales, se modeló el crecimiento y desarrollo, medido a través de las variables: diámetro del tallo, masa seca, área foliar y altura del tallo, con la edad de la planta, en semanas, como variable predictora.
Se evaluaron los modelos propuestos por Kiviste et al. (2002): Exponencial, Terazaki, Korf (Bailey-Clutter o Schumacher), Gamesi, Korsun, Gram, Gompertz, Gompert-Wenk, Sloboda, Verhults-Logística, Wingert, Peral-Reed, Simek, Moiseev III, Weber, Monomolecular-Weber, Mitscherlich I (Richards-Chapman), Mitscherlich II, Mitscherlich III, Bertalanffy, Weibull I, Weibull II, Todorovic III, Van der Vliet, Kövessy, Thomasius I, Thomasius II y Bass.
La comparación del ajuste de los modelos se basó en los valores de cuatro estadísticos: Sesgo, Error Medio Cuadrático, Coeficiente de Determinación de Predicción y Estadístico PRESS.
Para evaluar el efecto de los factores luz y variedad cada uno de ellos con dos niveles así como sus posibles interacciones, se usó un diseño de parcelas divididas, en el que el efecto de transplante se eliminó por bloqueo. Los diferentes niveles del factor luz se asignaron a las unidades experimentales principales con base en un esquema de aleatorización por bloques completos al azar con 12 repeticiones, donde el bloque consistió en un conjunto de ocho camas en las que se realizó el transplante en la misma época. En cada subunidad -dos camas- se realizó una asignación aleatoria de cada uno de los niveles del factor variedad. El análisis de varianza se realizó mediante el programa SAS.
Para dar cumplimiento al objetivo relacionado con la comparación de los protocolos de adición lumínica nocturna evaluados en esta investigación, los datos obtenidos fueron sometidos a análisis de varianza, para evaluar el efecto de los factores luz y variedad.
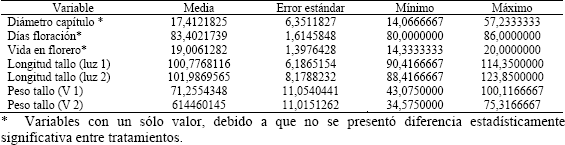
El efecto de la interacción entre los factores luz y variedad no resultó significativo para vida en florero (P = 0,2759), además el tratamiento lumínico no afectó la vida en florero (P = 0,6380), ni hubo diferencia estadísticamente significativa de la vida en florero entre las dos variedades (P = 0,8633). El valor promedio de esta variable fue de 19 días. De los resultados anteriores se deduce que la disminución en el suministro de la adición lumínica nocturna no afectó, en ninguna de las dos variedades, el tiempo de duración de las flores en el florero.
El efecto de la interacción entre los factores luz y variedad tampoco resultó significativo para los días a floración (P = 0,7603). El tratamiento lumínico no afectó significativamente los días a floración (P = 0,4082) y no hubo diferencia estadísticamente significativa de los días a floración entre las dos variedades (P = 0,2303). El valor promedio de esta variable fue de 83,4 días. Así, de los resultados anteriores se deduce que la disminución en el suministro de la adición lumínica nocturna no afectó el promedio los días a floración (tomados como el día en el cual se cosecha el 50 % de la cama), en ninguna de las dos variedades.
El efecto de la interacción entre los factores luz y variedad no resultó significativo para el diámetro del capítulo (P = 0,4286), así como el tratamiento lumínico no afectó significativamente el diámetro del capítulo (P = 0,3027) tampoco hubo diferencia estadísticamente significativa del diámetro del capítulo entre las dos variedades (P = 0,2763). El valor promedio de esta variable fue de 17,41 cm. De los resultados anteriores se deduce que la disminución en el suministro de la adición lumínica nocturna no afectó el diámetro promedio del capítulo en ninguna de las dos variedades.
El efecto de la interacción entre los factores luz y variedad no resultó significativo para la longitud del tallo (P = 0,8266); sin embargo, el tratamiento lumínico afectó significativa-mente la longitud promedio del tallo (P = 0,0280), disminuyéndola en 2,36 cm. El efecto de la variedad sobre la longitud del tallo no fue significativo (P = 0,3997).
En resumen, el tratamiento lumínico no afectó los días que dura la flor en florero, ni el diámetro del capítulo, que son los parámetros de mayor interés para el comprador. Solamente se afecta la longitud de los tallos evaluada al momento de la cosecha.
El efecto de la interacción entre los factores luz y variedad no resultó significativo para la masa fresca del tallo (P = 0,7225). El tratamiento lumínico no afectó significativamente la masa fresca del tallo (P = 0,1289). No obstante, se presentó diferencia estadísticamente significativa para la masa fresca del tallo entre las dos variedades (P = 0,0054): la variedad SY tuvo una masa fresca promedio 9,8 g por debajo de la de la variedad SW. La no significancia de la interacción indica que la masa fresca del tallo para cada una de las variedades evaluadas es independiente del tratamiento de luz a que fueron sometidas, siendo la variedad el único factor con efecto significativo sobre esta variable. En la Tabla 1 se presentan los valores de media, error estándar, mínimo y máximo para cada una de las variables analizadas.
Tabla 1. Valores para las variables analizadas en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), tipo spider, variedades Super White y Super Yellow, bajo dos tratamientos de luz.
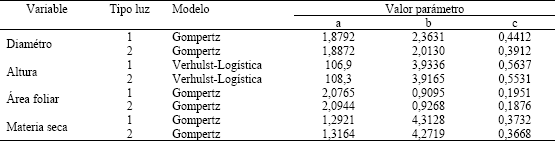
Al evaluar, mediante regresiones no lineales, el crecimiento y desarrollo floral, expresados a través de la evolución en el tiempo de las variables diámetro del tallo, área foliar, masa seca y altura, se encontraron los mejores ajustes al usar los modelos Gompertz y Verhulst-Logística. En la Tabla 2 se presentan los modelos y los valores de sus parámetros que mejor explicaron el comportamiento para cada una de las variables evaluadas; las expresiones generales se muestran a continuación:
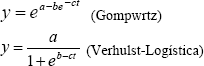
donde:
y: Variable cuyo comportamiento se modeló
t: Tiempo en semanas
a, b y c: Parámetros del modelo
Tabla 2. Modelos utilizados para estimar el comportamiento del crecimiento y desarrollo en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), tipo spider, variedades Super White y Super Yellow, bajo dos tratamientos de luz
En las Figuras 4, 5, 6, y 7 se presenta el comportamiento de las variables a través del tiempo, para los dos tratamientos.
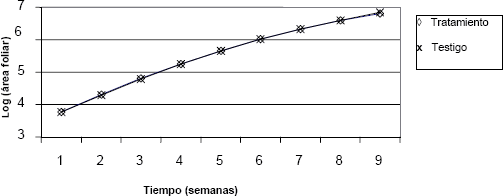
Figura 4. Comportamiento del logarítmo del área foliar a través del tiempo en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), bajo dos tratamientos de luz.
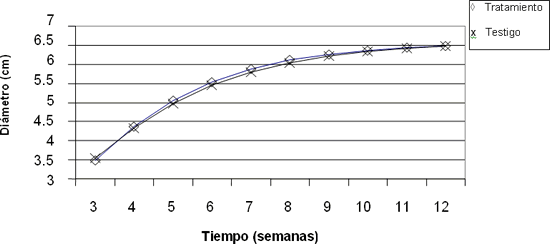
Figura 5. Comportamiento del logarítmo de la masa seca a través del tiempo en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), bajo dos tratamientos de luz.
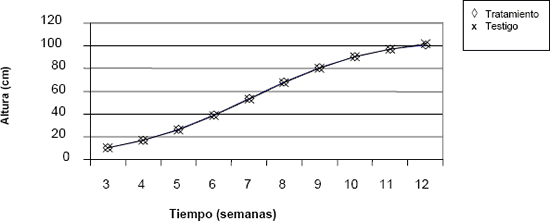
Figura 6. Comportamiento de la variable altura de la planta a través del tiempo en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), bajo dos tratamientos de luz.
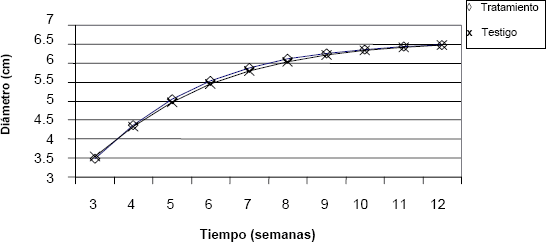
Figura 7. Comportamiento de la variable diámetro del capítulo floral a través del tiempo en crisantemo (Dendranthema grandiflorum (Ramat.) Kitam.), bajo dos tratamientos de luz.
En las figuras anteriores se observa que no hay diferencia en el comportamiento de las variables: diámetro, área foliar, masa seca y altura, entre los grupos sometidos a los diferentes tratamientos lumínicos. Es posible afirmar, a partir de dichos resultados, que la disminución en el suministro lumínico nocturno no afectó el crecimiento de ninguna de estas dos variedades.
Los resultados de esta investigación muestran que la disminución del suministro lumínico nocturno, con base en el esquema propuesto, no afectó el crecimiento y desarrollo de la planta ni los días necesarios para alcanzar la floración. En consecuencia, la vida de los tallos en florero tampoco se vio afectada.
El modelo de ampliación del fotoperíodo utilizado por el cultivo, consistente en interrumpir la noche con cortos periodos de luz (5 minutos de luz por 25 de oscuridad), es consecuente con el planteamiento hecho por Martínez-Zapater y Salinas (1993), quienes afirman que para que la noche tenga efecto inductivo hacia la floración, la oscuridad debe ser continua. La interrupción del periodo oscuro con unos pocos minutos de luz elimina el efecto inductivo de una noche larga en plantas de día corto.
Se ha demostrado que el efecto inductivo sobre la floración está dado por la longitud de la noche; no por la del día (Wareing y Phillips, 1981). Kofranek (1992) indica que la adición lumínica nocturna para plantas de día corto, entre las cuales está Dendranthema grandiflorum (Ramat.) Kitam., debe suministrarse de tal forma que el periodo oscuro no supere las cinco horas continuas; de lo contrario, se generarían las condiciones necesarias para la inducción floral.
Lo anterior sugiere que durante el periodo de oscuridad sucede algún evento promotor de los procesos que desencadenan la floración. Sin embargo, los cambios morfológicos, como la floración, no sólo están influenciados por factores externos como luz, temperatura y humedad, sino que responden a complejas interacciones genéticas asociadas con la duración del fotoperíodo. Según Hayama y Coupland (2003), la floración está mediada por la regulación trascripcional del reloj circadiano y por una postranscripción condicionada por longitudes de día específicas. La interacción entre ambas circunstancias activa proteínas para el control de la floración. Por ejemplo, recientes estudios realizados por estos autores, en Arabidopsis thaliana L., han candidatizado a los genes Circadian Clock Associated 1 (CCA 1), Late Elonged Hipocotyl (LHY) y Timing of Cb ExPresión 1 (TOC 1), como los responsables del modelo del comportamiento de la floración, a través del oscilador circadiano. En arroz, planta de día corto como el crisantemo, se ha aislado el loci QTL asociado con su sensibilidad fotoperiódica. En este loci se encontraron genes relacionados con proteínas similares ligadas a los genes encontrados en especies de día largo como Arabidopsis thaliana.
Basándose en este comportamiento, Hayama y Coupland (2003) plantean que el mayor control sobre el tiempo de florecimiento lo ejerce el factor genético. Argumentan en el mismo sentido que el gen CO, inducido, según Piñeiro y Coupland (1998), en mayor cantidad en días largos es el promotor de la expresión del gen FT, involucrado en la promoción del florecimiento en plantas con este tipo de requerimiento fotoperiódico.
Aunque es evidente el control intrínseco (genético) sobre la floración, también es claro que ésta responde a varios factores ambientales como la temperatura y la luz. Existen numerosos trabajos en los que se analiza el efecto de la fuente y la calidad de la luz, entre los cuales puede mencionarse el realizado por Heuvelink et al. (2002), en el cual se evaluó el efecto del uso de lámparas de alta presión de sodio (HPS) sobre el crecimiento en Dendranthema indicum (L.) Desmoul. La luz suplementaría suministrada en invierno (HPS; 48 µmol m-2 s-1 PAR), incrementada en un 24 % logró aumentar en un 45 % la producción de materia seca por m2. El efecto de luz suplementaria sobre la masa fresca y seca, y sobre el número de flores por planta a diferentes densidades (32, 48 y 64 plantas m2), fue alta a bajas densidades. Estos autores resumen que se presenta una relación lineal entre la acumulación y producción de masa seca y la acumulación o intercepción de radiación fotosintéticamente activa (PAR). Langton (1987) investigó la interrupción de la variación y la combinación de los fotoperíodos sobre la calidad final en crisantemos. Plantea que cuando el florecimiento de esta especie es consecuencia de un tratamiento de día corto, una interrupción con día largo puede introducir un mejoramiento de la calidad obtenida durante el invierno. Según este trabajo se concluyó que la interrupción con un día largo al sexto día, después de la inducción floral, mostró la mejor respuesta.
Otros trabajos reportan experiencias sobre el efecto del momento de la interrupción del periodo oscuro, tales como los realizados por Blacquère (2002), quien evaluó el tiempo requerido (ciclo luz-oscuridad) para retardar la floración en crisantemo. Utilizando como fuente de luz HPS, encontró que después de ocho horas de oscuridad, son suficientes ocho minutos de irradiación para lograr el efecto de día largo en esta especie vegetal. Karlsson et al. (1987) evaluaron la influencia de la temperatura sobre la eficiencia de luz en crisantemo. Encontraron que la ganancia de peso durante el desarrollo de la planta es principalmente una función del flujo de fotones fotosintéticos (FPR), y que la temperatura tuvo una pequeña acción sobre el incremento alcanzado por la planta. Concluyen que una alta calidad de la planta se obtiene cuando éstas crecen bajo altos niveles de PFR (mayores que 10 mol día-1) y una temperatura, tanto diurna como nocturna, entre 14 °C y 20 °C. No obstante, son escasos los trabajos reportados que tratan lo relativo a la cantidad del complemento lumínico nocturno. En esta investigación se evaluó el efecto de modificar la cantidad de luz sobre el comportamiento de la floración en una especie de día corto.
La respuesta fisiológica de la planta a la luz está mediada por una estructura capaz de captar y medir las diferentes longitudes de onda, generando una señal en respuesta. Esta estructura, llamada fitocromo, se encuentra bajo dos formas fotorreversibles denominadas Pr (Phytochrome Red) y Pfr (Phytochrome Far Red) (Bergareche y Moysset, 1993).
Cuando la forma Pfr absorbe luz se convierte a la forma Pr; y cuando la forma Pr absorbe luz se convierte a la forma Pfr. La forma Pr tiene el máximo pico de absorbancia para longitudes de onda roja (666 nm); mientras que el máximo pico de absorbancia para la forma Pfr se da para las longitudes de onda roja lejana (730 nm). Sin importar la longitud de onda predominante, las dos formas coexisten en equilibrio dinámico, debido a que el espectro de absorbancia de ambas formas se traslapa significativamente en la región roja del espectro, además de que la forma Pr absorbe una pequeña cantidad de luz en la región roja lejana del espectro (Taiz y Zeiger, 1998).
Dado que la luz del día contiene todas las longitudes de onda, durante el día se sintetizan ambas formas del fitocromo. Salisbury y Ross (2000) reportan que al momento del crepúsculo, la proporción es de aproximadamente 60:40, a favor del Pfr. Al entrar la noche, la longitud de onda roja lejana predomina sobre la roja, ocasionando un incremento de la forma Pr.
Mediante estudios realizados por Taiz y Zeiger (1998) se ha demostrado que cuando la proporción Pfr:Pr está a favor de la forma Pfr existe una mayor respuesta fisiológica. Esta respuesta fisiológica, según Wereing y Phillips (1981), es la inhibición de la floración. De ahí que para prolongar la fase vegetativa de la planta y retardar su floración se requieran altos niveles de fitocromo de la forma Pfr. Como esta forma se convierte a Pr durante la oscuridad cuando predomina la longitud del rojo lejano, se hace necesario interrumpir el periodo oscuro, lo cual se logra suministrando luz artificial durante la noche. El tratamiento fotoperiódico aplicado en esta investigación sí parece haber afectado la relación Pfr:Pr para retardar la floración.
Los resultados obtenidos en este estudio, bajo ambos tratamientos de estímulo lumínico nocturno, concuerdan con el planteamiento hecho por varios autores (Bernier, 1988; Fides, 1990; Taiz y Zeiger 1998; Salisbury y Ross, 2000), quienes indican que al interrumpir el periodo oscuro se mantienen altos los niveles del fitocromo Pfr y, por tanto, se logra alargar el periodo vegetativo de la planta y evitar la floración. Así, por ejemplo, Bunnig (1937) citado por Salisbury y Ross (2000) desarrolló investigaciones con plantas de día corto y encontró que suministrando luz roja durante cuatro horas cada 64 horas de oscuridad se lograba inhibir la floración.
En este trabajo, aunque no se cuantificaron las proporciones de las diferentes formas del fitocromo, es de suponer que, durante el tiempo en que se suministró la adición lumínica nocturna, se mantuvo suficiente cantidad de fitocromo bajo la forma Pfr como para retardar la floración hasta los 85 días después del transplante (ddt).
En investigaciones con Xanthium, Salisbury y Ross (2000) demuestran que si bien existe un fotoperíodo por debajo del cual se induce la floración (fotoperíodo crítico inductivo FCI), se requiere un fotoperíodo aun menor para que dicha floración se desarrolle normalmente (fotoperíodo crítico de desarrollo FCD), es decir, a velocidades adecuadas. Cuando no se cuenta con el FCD apropiado, los diferentes estadios florales se retardan. Según Fides (1990), el FCI para especies de día corto, como las del género Dendranthema, es de 14,5 horas, mientras que su FCD es de 13,5 horas.
Los anteriores reportes no coinciden con lo observado en el intertrópico y, particularmente en el cultivo donde se desarrolló este estudio, pues al tomar como base el trabajo de Bustamante (1989) se encuentra que la duración del día en la zona osciló entre 12,03 horas (septiembre) y 11,65 horas (diciembre), durante el tiempo en que se realizó el trabajo. El fotoperíodo máximo, que se obtiene al sumar el complemento lumínico nocturno suministrado por el cultivo (80 minutos cada noche; 1,33 horas) con los días más largos del periodo evaluado, es de 13,36 horas; y el mínimo es de 12,98 horas, siendo ambos valores de fotoperíodo claramente inferiores al FCI mencionado por Fides (1990) para plantas de día corto como las del género Dendranthema.
Se plantea, por tanto, con base en los resultados obtenidos en esta investigación, que para Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow, el FCI puede ser menor de las 14,5 horas propuestas para este género, y que el FCD puede ser menor de 13,5 horas. En consecuencia, la duración del periodo oscuro necesario para la inducción de la floración, denominado por Vince-Prue (1983) como longitud crítica de la noche o noche crítica inductiva, puede ser superior a 9,5 horas.
Los fotoperíodos alcanzados con la adición lumínica nocturna suministrada bajo el tratamiento testigo días largos ininterrumpidos oscilaron entre 12,98 y 13,36 horas. Así los periodos nocturnos o fase afótica estuvieron entre 11,02 y 10,64 horas, respectivamente. Luego, se tiene que la cantidad de horas luz suministrada durante esta investigación para mantener en estadio vegetativo a Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow, estuvo por debajo de las 14,5 horas sugeridas por varios autores para lograr dicho efecto vegetativo (Vince-Prue, 1983; Wareing y Phillips, 1986; Bernier, 1988; Fides, 1990). Consecuentemente, la fase afótica alcanzada con el protocolo testigo fue mayor a las 9,5 horas propuestas por los mismos autores para inducir la floración.
Tales resultados coinciden con lo encontrado por otros autores acerca de que las plantas clasificadas como de día corto tienen requerimientos de luz (fotoperíodo) y de oscuridad (fase afótica) de diferente duración. Por ejemplo, Salisbury y Ross (2000) listan varias especies vegetales clasificadas como plantas de día corto con requerimientos de noche crítica inductiva de diferente duración: Lemna perpusilla Torrey, 12 horas; Pharbitis nil Choisy, 9-10 horas; y Xanthium strumarium L. 8,3 horas. De otro lado, Veit et al. (2004), en estudios con Chenopodium rubrum L., especie de día corto, encontraron que su fotoperíodo crítico inductivo es de seis horas.
Por otra parte, bajo el tratamiento de adición lumínica nocturna propuesto en esta investigación, durante los 26 y 28 días que dura dicho complemento para cada una de las variedades, se suministraron dos días de luz complementaria por uno sin el complemento. Es decir que con este protocolo, cada tres días, durante el periodo de adición, se mantenía una noche de completa oscuridad o noche crítica inductiva, o sea un periodo afótico superior a 9,5 horas, convirtiéndose ello en un factor que podría disparar la floración; sin embargo, no sucedió así. No hubo diferencia significativa para los días a floración entre los dos protocolos utilizados: con interrupción y sin interrupción.
Si el periodo afótico aplicado cada tercer día hubiese sido inductivo, las plantas bajo el tratamiento propuesto habrían alcanzado su pico de floración antes de los 85 ddt. Se observó, sin embargo, que dicho periodo de oscuridad no tuvo efecto en adelantar la floración en ninguna de las dos variedades evaluadas. Lo anterior sugiere que Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow no responde a una sola noche crítica inductiva, y que necesita de más de un ciclo inductivo (día corto-noche larga) para pasar del estadio vegetativo al estadio reproductivo o de floración.
Este hecho coincide con el presentado por Salisbury y Ross (2000), quienes se refieren a una variedad de Dendranthema grandiflorum (Ramat.) Kitam. que requiere varios ciclos inductivos para iniciar la inducción floral. De igual forma, Kefu et al. (2002) evaluaron varias longitudes del día (8, 10, 12, 14, 15 y 16 horas) y varios ciclos fotoinductivos sobre Suaeda salsa (L.) Pall. Concluyeron que la longitud del día crítica para esta especie es de 14 horas y que se requieren mínimo ocho ciclos fotoinductivos para inducir la floración.
Para Vince-Prue (1983), el mecanismo de percepción de la luz por parte de las plantas obedece a procesos que alternan a través de fases de alta y baja actividad, con una periodicidad regular de cerca de 24 horas. Estos cambios rítmicos se conocen como circadianos. Bönning (1936), citado por Vince-Prue (1983), sostiene que el sistema circadiano de fotoperiodicidad de las plantas involucra una oscilación regular de fases con diferentes sensibilidades a la luz. Al periodo de 12 horas de requerimientos de luz lo denomina fase fotofílica, a la fase de 12 horas de requerimiento de oscuridad la denomina escotofase. Bernier (1988) sostiene que la medición de estos periodos lumínicos brinda información a la planta para desencadenar y desarrollar procesos fisiológicos, tales como la floración. Estos mecanismos pueden ser alterados modificando la duración de las fases. Así se logra producir efectos de día corto en plantas de respuesta a días largos y viceversa, es decir, se logra inhibir la floración. Sin embargo, debe existir concordancia entre el estímulo externo y las condiciones internas para que se desencadene determinado proceso fisiológico. En este caso, en particular, las condiciones externas de una escotofase larga (fotofase corta) deben coincidir con niveles bajos del fitocromo de la forma Pfr, para que se induzca la floración.
Cuando las plantas sometidas a los tratamientos de adición lumínica nocturna retornaron a las condiciones naturales de días cortos y noches largas, se inició el proceso de inducción floral, que concluyó en la floración 55,4 días después para Super Yellow y 57,4 para Super White (full de corte). Estos resultados coinciden con los planteamientos realizados por varios autores (Vince-Prue, 1983; Wareing y Phillips, 1986; Fides, 1990), quienes afirman que las interrupciones de las fases oscuras de las plantas de día corto con destellos de luz, logran influir para retardar la floración sin alterar el ritmo circadiano de la planta. Es decir, el ritmo puede alterarse por determinado tiempo, sin que ello logre modificar la memoria fotoperiódica de la planta.
Para Martínez-Zapater (1983), en plantas herbáceas, como el crisantemo, el programa de desarrollo reproductivo se presenta como continuación del programa de desarrollo vegetativo. El mismo autor indica que la transición entre ambos programas (transición floral) es el resultado de la interacción entre factores relacionados con el estado de madurez de la planta y factores medioambientales. Para las condiciones de la región en donde se realizó esta investigación, el periodo de la adición para la variedad Super White dura 26 días y para la variedad Super Yellow su duración es de 28 días. Así, se evidencia que existe variabilidad en el requerimiento lumínico entre plantas pertenecientes a una misma especie vegetal para pasar del estado vegetativo al estado de floración. Ello indica que la respuesta de la planta a factores externos está gobernada genéticamente y que dicha respuesta puede cambiar entre variedades, aun siendo clasificadas bajo la misma categoría de requerimientos lumínicos.
Los resultados obtenidos en este trabajo sugieren que la transición floral en Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow ocurre, como ya se ha mencionado, después de suministrar la totalidad de la adición lumínica nocturna, esto es, a partir de los 26 y 28 ddt, respectivamente. Luego, lo que ocurre con dicho complemento es un aumento en la actividad de crecimiento vegetativo, el cual está reflejado en la mayor altura en los tallos (2,36 cm en promedio) alcanzada cuando el suministro de la adición lumínica nocturna fue ininterrumpido. Esto hace suponer una mayor actividad vegetativa de las plantas sometidas al tratamiento testigo, frente a las plantas bajo el tratamiento de reducción en el suministro. Esta mayor altura de los tallos, alcanzada por el material que recibió mayor cantidad de luz, puede estar indicando que esos días de más son los que causan dicho aumento en la altura, pues fue el único factor que varió durante todo el experimento. No obstante, la menor altura alcanzada por los tallos, bajo el tratamiento propuesto de menor complemento lumínico nocturno, sigue estando por encima de la longitud comercial (70 cm), por lo que tal disminución no representa ninguna desventaja, sino, por el contrario, un ahorro energético (recursos lumínicos artificiales, químicos, fisiológicos, etc.), ya que actualmente se están produciendo en promedio 2,36 cm de más, que luego serán desechados. Surge, entonces, el interrogante sobre si la mayor acumulación de biomasa presentada por el testigo se debió a una mayor tasa fotosintética.
Los tratamientos planteados en esta investigación no permiten determinar con certeza el fotoperíodo óptimo para Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow. Sin embargo, se logra demostrar que disminuyendo la cantidad de luz suministrada rutinariamente por el cultivo no se modifican las características poscosecha de la variedad. En lo relativo al crecimiento y desarrollo de las variedades evaluadas, dicha disminución de la adición lumínica nocturna solamente afectó la altura del tallo; no obstante, el promedio de los tallos obtenidos bajo el protocolo propuesto continúa dentro de los estándares requeridos para su comercialización. En tal sentido la disminución en la utilización de energía lumínica para obtener plantas comerciales de Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow, bajo las condiciones en que se realizó esta investigación, redundaría en un ahorro económico para los floricultores de la región.
El ahorro por hectárea por año se estima teniendo en cuenta que durante el año se realizan cuatro cortes, que en una hectárea se ubican 180 camas y que la iluminación se suministra con base en guirnaldas de 11 bombillos de 200 vatios por cada tres camas. Además, el valor del Kilovatio/hora, para la región, tiene un costo diferencial así: $147 entre las 01:00 y las 03:00 horas; y $162 el resto de la franja nocturna. Luego, bajo el esquema testigo de complemento lumínico nocturno, el costo por concepto de consumo energético por hectárea/año es de $2.896.608 para SW y de $3.119.424 para SY. Bajo el esquema de complemento lumínico nocturno propuesto, el costo por concepto de consumo energético por hectárea/año es de $2.005.344 para SW y de $2.116.752. Esto representa un ahorro de $891.264 (30,77 %) para SW y de $1.002.672 (32,14 %) para SY.
La vida promedio en florero de los tallos sometidos a los diferentes tratamientos de luz no difirió significativamente, en ninguna de las variedades Super White y Super Yellow de Dendranthema grandiflorum (Ramat.) Kitam., tipo Spider. Tampoco se detectaron diferencias al comparar las medias de esta variable entre las dos variedades. Esto significa que el tratamiento propuesto, consistente en disminuir el subsidio lumínico nocturno, no afectó la durabilidad poscosecha de la flor. Para la población representada en este estudio, es posible disminuir el subsidio lumínico nocturno en un 30,769 % para Dendranthema grandiflorum (Ramat.) Kitam., tipo Spider variedad Super White, y en un 32,143 % para Dendranthema grandiflorum (Ramat.) Kitam., tipo spider variedad Super Yellow, sin afectar significativamente la vida media en florero.
Dado que los días a floración no mostraron diferencia significativa entre variedades, aun suministrando un complemento lumínico nocturno con dos días de diferencia entre éstas, se ratifica que, bajo las condiciones en que se realizó esta investigación, las variedades Super White y Super Yellow de Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, presentan diferentes requerimientos de complemento lumínico nocturno.
La eliminación del suplemento lumínico nocturno durante una noche, en medio de dos noches con suplemento lumínico, no tiene efecto inductivo para la floración en Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow, bajo las condiciones en que se realizó esta investigación. Ambas variedades requieren más de una noche sin suplemento lumínico nocturno para que se desencadene el proceso inductivo de la floración.
El valor de 14,5 horas, propuesto por varios autores para plantas de día corto como el fotoperíodo crítico inductivo (FCI) o cantidad de horas luz necesarias para inducir la floración, parece no cumplirse en las dos variedades evaluadas, bajo las condiciones en que se desarrolló este trabajo. Puede afirmarse, con base en los resultados de este estudio, que el FCI está alrededor de las 13 horas.
El valor de 13,5 horas, propuesto por varios autores para plantas de día corto como la cantidad de horas luz necesarias para que la floración se desarrolle normalmente o fotoperíodo crítico de desarrollo (FCD), parece no cumplirse para las dos variedades evaluadas, bajo las condiciones en que se ejecutó este estudio. Según los resultados obtenidos con la aplicación de los dos protocolos utilizados, el FCD puede ser menor de 13,5 horas.
La interrupción del periodo afótico de la planta con ciclos cortos de luz (alrededor de 5 minutos) parece no alterar el ritmo circadiano de la especie evaluada. Se presume, a partir de la inhibición del proceso de floración, que la cantidad sintetizada de fitocromo bajo la forma Pfr se logró mantener en niveles adecuados como para inhibir la floración.
Al someter las plantas de Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow, a las diferentes condiciones de adición lumínica nocturna, no se presentaron diferencias en el desarrollo vegetativo.
De las variables evaluadas, el tratamiento propuesto de disminución del complemento lumínico nocturno sólo afectó la altura del tallo, haciéndola disminuir en promedio 2,36 cm en relación con el tratamiento testigo. Esta longitud (99,99 cm), sin embargo, sigue estando por encima de la longitud comercial (70 cm), por lo que tal disminución no representa ninguna desventaja, sino, por el contrario, un ahorro energético (recursos lumínicos artificiales, químicos, fisiológicos, etc.), pues actualmente se están produciendo en promedio 2,36 cm de más, que luego son desechados.
La duración en florero y el diámetro del capítulo, dos de los parámetros de mayor interés para comercializar la flor, no resultaron afectados por el tratamiento propuesto de disminución del complemento lumínico nocturno.
Con los resultados obtenidos en esta investigación no es posible determinar con certeza cuáles son los requerimientos fotoperiódicos óptimos para las variedades evaluadas. Sin embargo, puede indicarse que el FCI está alrededor de 13 horas, valor inferior a las 14,5 horas propuestas para este tipo de plantas. En consecuencia, el periodo afótico puede estar alrededor de 11 horas, el cual es superior a las 9,5 horas propuestas para especies de día corto.
El actual esquema de adición lumínica nocturna podría disminuirse en aproximadamente un 30 %, sin que se vean afectados el crecimiento y desarrollo ni las características poscosecha de Dendranthema grandiflorum (Ramat.) Kitam., tipo spider, variedades Super White y Super Yellow.
Este trabajo se considera de carácter exploratorio, de ahí que se recomiende continuar con ensayos en donde se evalúen otros protocolos que incluyan:
- Aumento de los periodos afóticos a que se someten las plantas durante la duración de los días de complemento lumínico nocturno; es decir, aumentar el número de noches críticas inductivas, por ejemplo a dos.
- Aumentar los intervalos de oscuridad entre los destellos de luz; por ejemplo, que éstos sean de 60 minutos.
- Disminuir la duración del destello lumínico suministrado, de manera que se reduzcan los 80 minutos de complemento lumínico nocturno que actualmente se suministran. Podrían ensayarse destellos de luz de tres o cuatro minutos entre las fases de oscuridad.
Acompañar posteriores estudios de evaluación fotoperiódica con un testigo absoluto; es decir, un tratamiento bajo condiciones naturales de luzoscuridad. Ello permitiría determinar con exactitud el número de días necesarios para la floración del material evaluado sin ningún complemento lumínico nocturno, por consiguiente, los días que se retarda la floración, producto del complemento lumínico nocturno. Dado que el testigo absoluto representa una pérdida, tal tratamiento podría tener menos repeticiones que los demás.
Acompañar estos estudios (de requerimientos fotoperiódicos) con un seguimiento histológico que evalúe los diferentes tiempos en que se suceden los distintos estadios de la diferenciación floral. Ello permitiría establecer regiones exploratorias con el fin de determinar los tiempos requeridos para cada una de las fases involucradas en el proceso de floración.
Se recomienda que antes de adoptar rutinariamente el protocolo de disminución de complemento lumínico nocturno evaluado durante esta investigación, se realicen nuevos ensayos para verificar los resultados presentados en este trabajo.
BERGARECHE, C. y MOYSSET, L. El fitocromo. En: AZCÓN-BIETO, J. y TALÓN, M., eds. Fisiología y bioquímica vegetal. Nueva York: Interamericana McGraw Hill, 1993. p. 393-419. [ Links ]
BERNIER, G. The control of floral evocation and morphogenesis. En: Annual Review of Plant Physiology and Plant Molecular Biology. Vol. 39 (1988); p. 75-219 . [ Links ]
BLACQUIÈRE, T. How much light is needed for the prevention of flowering of cut chrysanthemums when using intensity HPS lighting as a night break. En: Acta Horticulturae. Vol. 580 (2002); p. 69-75. [ Links ]
BUSTAMANTE BETANCUR, Fabio. Riego por aspersión. En: CURSO SOBRE ACTUALIZACIÓN EN SISTEMAS DE RIEGO (1989: Medellín). Conferencias del Curso sobre actualización en sistemas de riego. Medellín: Universidad Nacional de Colombia, 1989. p. 172-239. [ Links ]
CARTAGENA, J. R.; MARTÍNEZ, E. y HOYOS, R. Manual de prácticas de laboratorio de Fisiología Vegetal, 12 ensayos. Medellín: Universidad Nacional de Colombia, 2001. 100 p. [ Links ]
ESPINAL, T. L. S. Apuntes ecológicos. Medellín: Universidad Nacional de Colombia, 1992. 234 p. [ Links ]
FIDES, H. BV. Fides mum manual: for all year round chrysanthemums. The Netherlands : De Leir, 1990. 101 p. [ Links ]
HAYAMA, R. and COUPLAND, G. Shedding light on the circadian clock and the photoperiodic control of flowering. En: Plant Biology. Vol.6, No.1 ( 2003); p. 13-19. [ Links ]
HEUVELINK, E. et al. Light on cut chrysanthemum : measurement and simulation of crop growth and yield. En: Acta Horticulturae. Vol. 580 (2002); p. 197-202. [ Links ]
KARAGUZEL, O. et al. Responses of native Lupinus varius (L.) to culture condition : effects of photoperiod and sowing time on growth and flowering characteristics. En: Scientia Horticulturae. Vol. 103, no.3 (Jan. 2005) Disponible en Internet: www.sciencedirect.com/science (Consultado en octubre 26 de 2004). [ Links ]
KARLSSON, M. G.; HEINS, R. D. and HOLCOMB, E. J. Influence of temperature, photosynthetic flux and plant age on light utilization efficiency in chrysanthemum. En: Acta Horticulturae. Vol. 197 (1987); p. 21-30. [ Links ]
KEFU, Z. et al. Critical day-lenght and photoinductive cycles for the induction of flowering in halophyte Suaeda salsa. En: Plant Science. Vol. 162, No. 1 (2002); p. 27-31. [ Links ]
KIVISTE, A.et al.Funciones de crecimiento de aplicación en el ámbito forestal. Madrid:Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, 2002. 190 p. (Monografías INIA: Forestal, no. 4). [ Links ]
Madrid: Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, 2002. 190 p. (Monografías INIA: Forestal, no. 4).
KOFRANEK, A. A. Cut chrysanthemums. En: LARSON R. A., ed. Introduction to floriculture: 2ed. San Diego, California: Academic Press, 1992. p. 5-42. [ Links ]
LANGHANS, R. W. Chrysanthemums: a manual of culture, diseases, insects and economics of chrysanthemum. The New York State Extension Service Chrysanthemum School. 1964. 185 p. [ Links ]
LANGTON, F. A. Apical dissection and light-integral monitoring as methods to determine when long-day interruptions should be given in chrysanthemum growing. En: Acta Horticulturae. Vol. 197 (1987); p. 31-42. [ Links ]
MARTÍNEZ-ZAPATER, J. M. y SALINAS, J. La transición floral. En: AZCON-BIETO, J. y TALON, M., eds. Fisiología y bioquímica vegetal. Madrid: Interamericana-Mc Graw Hill, 1993. p. 435-449 [ Links ]
PIÑEIRO, M. and COUPLAND, G. The control of flowering time and floral identity in Arabidopsis. En: Plant Physiology. Vol. 117 (1998); p. 1-8. [ Links ]
SALISBURY, F. y ROSS, C. Fisiología de las plantas. México D. F.: Iberoamericana, 2000. 759 p. [ Links ]
TAIZ, L. and ZEIGER, E. Plant physiology. 2ed. Sunderland: Sinauer Associates, 1998. 791 p. [ Links ]
THOMAS, B. and VINCE-PRUE, D. Photoperiodism in plants. San Diego, California: Academic Press, 1997. 428 p. [ Links ]
VEIT, J.; WAGNER, E. and ALBRECHTOVÁ, J. Isolation of a floricaula/leafy putative orthologue from Chenopodium rubrum and its espression during photoperiodic flower induction. En: Plant Physiology and Biochemistry. Vol. 42, No. 7-8 ( 2004); p. 573-578. [ Links ]
VINCE-PRUE, D. and COCKSHULL, K. E. Photoperiodism and crop production. En: JHONSON, C. B., ed. Physiological process limiting plant productivity. London; Boston: Butterworths, 1981. p. 175-197. [ Links ]
VINCE-PRUE, D. Photomorphogenesis and flowering. En: MOHR H., ed. Encyclopedia of Plant Physiology New Series. Vol. 16B (15). 1983. p. 457-499. [ Links ]
WAREING, P. F. and PHILLIPS, I. D. J. Growth and differentiation in plants. 3ed. New York: Pergamon Press, 1981. 343 p. [ Links ]

