










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.58 no.2 Medellín July/Dec. 2005
BANCO DE SEMILLAS EN UN BOSQUE DE ROBLE DE LA CORDILLERA CENTRAL COLOMBIANA
SOIL SEED BANK IN AN OAK FOREST FROM THE COLOMBIAN CENTRAL CORDILLERA
Luz Bibiana Moscoso Marín1 y Maria Claudia Diez Gómez2
1 Ingeniera Forestal. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <lbmosco@unalmed.edu.co>
2 Profesora Asociada. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <mcdiez@unalmed.edu.co>
Recibido: Noviembre 22 de 2004; aceptado: Mayo 25 de 2005.
RESUMEN
El banco de semillas del suelo es importante para el funcionamiento del ecosistema, pues se constituye en un reservorio de especies listas a germinar cuando se presenta una perturbación o cuando las condiciones ambientales cambian para iniciar el proceso de sucesión. El objetivo principal de esta investigación fue evaluar la variación espacial del banco de semillas y su relación con la vegetación actual en un fragmento de bosque altoandino dominado por roble de tierra fría (Quercus humbodtii Bonpl.) a 2.290 m.s.n.m. en la cuenca de la Quebrada Piedras Blancas, en el departamento de Antioquia, Colombia. Allí se estableció una parcela de 6.400 m², donde se eligieron 57 puntos de muestreo. En cada uno de ellos se colectó una muestra de hojarasca y una de suelo. Además se marcaron cinco subparcelas de 160 m2 para hacer el muestreo de la vegetación. La determinación del banco de semillas se hizo por el método de germinación directa, evaluando la emergencia de plántulas. En total, se censaron 204 individuos pertenecientes a 25 familias y 41 géneros. Las especies con mayor representación fueron Alfaroa cf. colombiana, Myrcia popayanensis Rich. y Quercus humboldtii Bonpl. En el ensayo de germinación se obtuvieron 428 individuos en las muestras de suelo y 113 en las de hojarasca, pertenecientes a 20 morfoespecies. La composición de especies del banco de semillas de este fragmento de robledal, no reflejó la composición actual de la vegetación del bosque. El banco de semillas esta constituido principalmente por especies herbáceas y en menor proporción por arbustos. La hojarasca contiene poca cantidad de semillas (198 semillas m-2) y en los primeros 10 cm del suelo se acumula una gran cantidad de semillas viables (4.330 semillas m-2). Se presenta una variación espacial significativa en la densidad de semillas del suelo a través del bosque a escala de unos pocos metros.
Palabras claves: Banco de semillas, bosques altoandinos, robledal, densidad, distribución espacial, diversidad de especies, riqueza, germinación, regeneración.
ABSTRACT
Soil seed banks are important for ecosystem functioning because they stores species ready to germinate when a perturbation occurs or when environmental conditions change to start the successional process. The main objective of this research was to evaluate the spatial variation of seed banks and its relation with the current vegetation in a highland Andean forest dominated by oak (Quercus humbodtii Bonpl.) at 2.290 m.a.s.l. in the watershed of the Piedras Blancas stream, in the department of Antioquia, Colombia. We sampled 57 points in a 6.400 m² plot established in that forest. In each point we collected one sample each of litter and soil. We also established five subplots of 160 m2 each for sampling vegetation. Seed banks were assessed by the method of direct germination. We obtained 204 trees belonging to 25 families and 41 genera. The most representative tree species were Alfaroa cf. colombiana, Myrcia popayanensis Rich. and Quercus humboldtii Bonpl. We obtained 428 individuals from soil samples and 113 individuals from litter samples in the germination trial, belonging to 20 morpho species. Species composition of the soil seed bank did not reflect the composition of the forest vegetation, and we found mainly herbaceous and shrub vegetation. Litter had 198 seeds m-2 and soil had 4.330 seeds m-2. There was large spatial variation in the amount of seeds in this forest fragment.
Key words: Seed bank, Andean highland forests, oak forest, density, spatial distribution, species diversity, richness, germination, regeneration.
 MÉTODOS
MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
AGRADECIMIENTOS
BIBLIOGRAFÍA
La composición y dinámica del banco de semillas en el suelo del bosque, son fundamentales para predecir el curso de la sucesión secundaria cuando se presentan perturbaciones (Garwood, 1989), por lo cual es un tema de gran relevancia en ecosistemas sometidos a procesos de conversión intensos. Este es el caso de los bosques alto andinos colombianos con una gran biodiversidad (Gentry, 1995, 2001; Henderson; Churchill y Luteyn, 1991) pero con una larga historia de conversión a monocultivos de gran escala, pequeñas parcelas para agricultura de subsistencia, pastos para la producción lechera (Cavelier y Tobler, 1998) y más recientemente para el establecimiento de cultivos ilegales (Cavelier y Etter, 1995). En algunas de estas áreas se hace evidente la necesidad de realizar acciones de restauración de la vegetación natural, partiendo de los mecanismos más apropiados en cada caso, entre los cuales se encuentra la activación del banco de semillas del suelo.
El banco de semillas es uno de los principales mecanismos de regeneración en ecosistemas forestales y en el caso de las especies pioneras puede ser más importante que la regeneración a partir de la lluvia de semillas (Cubiña y Aide, 2001). El banco de semillas está conformado por las semillas que permanecen latentes en el suelo, por lo cual es un reservorio de especies, principalmente pioneras, que están listas a germinar en el momento en que se presente una perturbación, se forme un claro o las condiciones ambientales cambien para iniciar de este modo un proceso de sucesión secundaria (Garwood, 1989).
La densidad y composición del banco de semillas puede variar a escala de unos pocos metros cuadrados (Bigwood y Inouye, 1988; Bertiller, 1998; Lortie y Turkington, 2002; Butler y Chazdon, 1998), y en bosques tropicales esta variación parece estar asociada con factores como la topografía (Singhhakumara; Uduporuwa y Ashton, 2000) y la distribución espacial de las especies actuales o del pasado, y sus los patrones de dispersión de semillas (Saulei y Swaine, 1988; Dalling, Swaine y Garwood, 1998). Estas características del banco de semillas también cambian en función de la profundidad en el suelo, y se pueden modificar estacionalmente a lo largo del año (Enright, 1985; Dalling, Swaine y Garwood, 1997).
El objetivo de esta investigación fue evaluar la variación espacial y en profundidad, del banco de semillas de un fragmento de bosque de roble de tierra fría (Quercus humboldtii Bompl.) en la región de Piedras Blancas, y establecer la relación existente entre el banco de semillas y la vegetación actual del robledal.
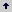
Área de estudio. El estudio se desarrolló en un fragmento de robledal de la cuenca de la Quebrada Piedras Blancas, localizada en la cordillera central colombiana (6°1815 Norte y 75° 30 20 Oeste), entre los 2.200 y 2.600 m.s.n.m. La temperatura promedia anual es de 15,5 °C, y la precipitación promedia anual es de 1.871 mm, distribuidos en dos períodos de lluvia al año (abril–mayo y septiembre–noviembre) (Duque 1993). Las principales formaciones de vegetación según el sistema de Holdridge, son bosque húmedo montano bajo (bh–MB), y bosque muy húmedo montano bajo (bmh–MB). El relieve es ondulado en casi toda el área, con suelos derivados de cenizas volcánicas (Andisoles); predominan los suelos ácidos, desaturados con alto contenido de materia orgánica , bajos en bases y con buenas propiedades físicas (Jaramillo 1989).
El uso del suelo predominante en la cuenca es la protección con vegetación natural compuesta por robledales y rastrojos, así como plantaciones forestales de las especies Cupressus lusitanica Mill, Pinus patula Schltdl y Cham. y Pinus elliotii Engelm. combinada con pequeñas áreas dedicadas a la agricultura y al pastoreo.
Trabajo de campo. Se estableció una parcela de 6.400 m² (80 m x 80 m) en un fragmento de bosque de roble localizado sobre relieve ondulado, con valores promedios de pendiente de 23 ° y a una altitud de 2.290 m.s.n.m. La profundidad promedia de la capa de hojarasca (descompuesta y sin descomponer) era de 18 cm, con valores extremos de 11 y 32 cm.
En esta parcela se establecieron cinco transectos de 160 m2 (4 m x 40 m), en los cuales se censaron y midieron todos los individuos arbóreos con DAP ³ 10 cm. Se incluyó además el reporte de las especies que por su porte o hábito de crecimiento podrían ser fuente de semillas, como hierbas, lianas y bejucos. Las muestras botánicas se empacaron para su posterior identificación en el herbario Gabriel Gutiérrez Villegas, (MEDEL), de la Universidad Nacional de Colombia, Sede Medellín.
En el perímetro de la parcela de 6.400 m² se establecieron puntos de muestreo a una distancia de 20 m, para un total de 16 puntos de intersección externos. En el interior de la parcela se elaboró una cuadrícula de 10 m, para obtener así 25 puntos de intersección adicionales. Finalmente, alrededor del punto central de esta parcela se marcaron otros 16 puntos con una distancia de 2,5 m (Figura 2). Se generaron en total 57 puntos de muestreo. En cada punto de intersección se colectó una muestra de la capa superficial del suelo (hojarasca) con un marco metálico de 10 x 10 cm, y se tomó también una muestra de suelo de 10 cm de profundidad con un barreno de cilindro de 4,7 cm de diámetro. De esta manera se obtuvieron 57 muestras de hojarasca y 57 muestras de suelo para un total de 114 muestras. Cada una de ellas se guardó en una bolsa plástica, se rotuló y se almacenó a baja temperatura hasta el momento de la siembra.
Evaluación de la emergencia de plántulas e identificación del material. La determinación de la densidad y composición del banco de semillas se hizo por el método de germinación directa, evaluando la emergencia de las plántulas. Estos ensayos se llevaron a cabo en el covertizo de la Estación Experimental de Piedras Blancas (temperatura entre 16 y 27 °C e humedad relativa entre 90 y 93 %).
Cada muestra de suelo se esparció en cubetas plásticas de germinación de 22 x 33 cm con tapa, sobre un sustrato de cuarzo fino inerte, que tenía una profundidad aproximada de 3 cm. Este sustrato se utilizó para darle soporte y retener la humedad de la muestra. La profundidad de la capa de suelo extendido sobre el cuarzo fue de aproximadamente 0,5 cm, lo cual permitió que toda la muestra quedara expuesta a iguales condiciones de luz, humedad y temperatura, de acuerdo con lo recomendado por Dalling, Swaine y Garwood (1994).
Las muestras de hojarasca se limpiaron cuidadosamente con una brocha pequeña para separar el componente más grande (hojas, ramas, flores, etc.) y extraer las posibles semillas presentes en ellas. Las muestras se mantuvieron con riego periódico y se realizaron observaciones una vez por semana, durante cinco meses a partir del montaje del ensayo, para determinar la emergencia total y la emergencia por especie. Las plántulas que emergieron marcaron con códigos, se contaron e identificaron utilizando claves. No se realizó un chequeo del suelo al final del experimento, para determinar si aún quedaban semillas sin germinar.
Para la determinación de las semillas y las plántulas se acudió a la comparación con las especies del sotobosque y a la ayuda de manuales de morfología de estructuras vegetales como embriones, cotiledones y de regeneración en general. Se registró el número de semillas por morfo especie y por muestra y en la identificación se trató en lo posible de llegar hasta familia, género y especie, tanto para las muestras de vegetación, como para las plántulas y las semillas.
Análisis de los resultados
Diversidad de especies. Para evaluar la diversidad de la vegetación actual y la diversidad del banco de semillas se utilizaron los siguientes índices de alfa diversidad: Riqueza de especies (S), índice de Margalef (DMg), índice de Menhinick (DMn), índice de Shannon (H´) (Magurran 1988). Para comparar la diversidad entre el banco de semillas y la vegetación, así como entre la hojarasca y el suelo, se utilizaron los siguientes índices de beta diversidad: Índice de Jaccard (Cj) y Coeficiente de Sorenson (Cs) (Magurran 1988).
Análisis de la distribución espacial. Para este análisis se partió del conjunto de puntos de muestreo en los que se midió la variable densidad de semillas, con base en los resultados de emergencia de plántulas. La representación de los puntos se hizo por medio del programa Arc-View GIS 3.2. Mediante el uso de elementos lineales, se unieron los puntos donde la variable tomaba rangos de valores cercanos. Se construyeron así líneas basadas en la interpolación lineal de valores medidos en una serie de puntos de control dentro del bosque.
Composición florística. En total se censaron 204 individuos pertenecientes a 32 géneros determinados y siete indeterminados; entre ellos se encuentran dos géneros de lianas con 11 individuos del género Cavendishia y un individuo de la familia Melastomataceae. El número total de familias encontradas en el muestreo fue de 25, de las cuales tres son indeterminadas. La familia con mayor representatividad, tanto en número de géneros como de especies, resultó ser Lauraceae, seguida por Myrtaceae y Melastomataceae. El 60 % de las familias encontradas, es decir 15 familias, están representadas por un solo género y la misma cantidad de especies. De la misma manera, el 16 % de las familias están representadas por dos géneros y dos especies. Solamente las familias Lauraceae y Melastomataceae, están representadas con 5 y 4 géneros respectivamente; esta última también con un género de lianas.
Entre las especies más representativas se encuentran Myrcia popayanensis, Clethra sp., Ilex danielis Killip and Cuatrec., Clusia sp. y Hedyosmum bonplandianum Mart., además de algunas especies de crecimiento secundario como Vismia cf. ferruginea Kunth, Befaria glauca Bonpl., Cavendishia pubescens Britton y Miconia sp., entre otras. Se destaca además, la presencia e importancia del Quercus humboldtii, como la especie dominante de la formación. Los resultados del índice de valor de importancia (IVI), para las especies vegetales presentes en la zona de estudio, muestran que la especie más abundante, más uniformemente distribuida y que presenta mayores tamaños es el roble Quercus humboldtii, con un valor de importancia del 52,96 %, seguida por Alfaroa cf. colombiana y Myrcia popayanensis, con valores de importancia de 30,41 % y 25,51 %, respectivamente (Figura 1).
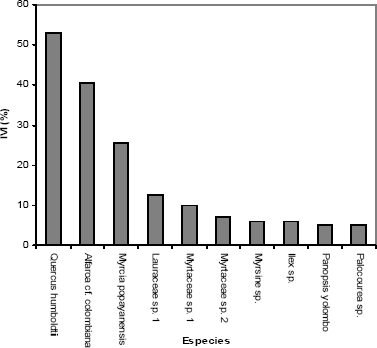
Figura 1. Índice de Valor de Importancia para las especies vegetales presentes en la cuenca de la quebrada Piedras Blancas, Antioquia-Colombia.
Emergencia de plántulas
Emergencia semanal. Los ensayos de germinación y emergencia de plántulas, se establecieron con 114 muestras del banco de semillas, de las cuales 57 eran de la capa de hojarasca y 57 de los primeros 10 cm de suelo. El seguimiento se realizó una vez por semana durante cinco meses, y se registró el número de plántulas que emergían por muestra (Tabla 1). El registro de la emergencia semanal para los sustratos suelo y hojarasca, muestra patrones similares a lo largo del tiempo para ambos componentes, aunque se presentan valores más altos en las muestras de suelo, excepto en las semanas 2 y 20, donde hay mayor emergencia en las muestras de hojarasca. Durante el tiempo del ensayo emergieron en total 541 plántulas; 428 de ellas (79 %), pertenecen a las muestras de suelo y 113 (21 %) a las muestras de hojarasca (Tabla 1). La densidad de semillas en la hojarasca fue de 198 semillas/m2 y la del suelo (en los primeros 10 cm) es de 4.330 semillas/m2.
Tabla 1. Número de plántulas que emergieron semanalmente en el banco de semillas del suelo y de la hojarasca, localizados en la cuenca de la quebrada Piedras Blancas, Antioquia-Colombia.
Determinación de plántulas y semillas: En la identificación de las plántulas se diferenciaron 22 morfo especies (Tabla 2). Sin embargo, una proporción significativa de individuos germinados, no pudieron ser identificados, ya que tenían tamaños muy pequeños y características morfológicas poco observables que se mantuvieron durante todo el tiempo de duración del ensayo. Se presentó una alta cantidad de semillas de roble (Quercus humboldtii), lo cual se vio favorecido por la época del muestreo, pues esta especie se encontraba en plena fructificación. Su presencia se limitó a la capa de hojarasca y por observaciones hechas en el campo se puede afirmar que no se almacenan por mucho tiempo en el banco de semillas, sino que germinan poco tiempo después de llegar al suelo.
Tabla 2. Determinación de plántulas por morfoespecie en el banco de semillas del suelo y la hojarasca establecidos en la cuenca de la quebrada Piedras Blancas, Antioquia-Colombia.

Diversidad de especies
Diversidad de la vegetación actual y diversidad del banco de semillas. Los índices de Margaleff y Menhinick muestran que hay una mayor riqueza de especies en la vegetación del robledal, con valores de 8,27 y 3,15, respectivamente, donde se alcanzaron valores de 3,01 y 0,86 para estos mismos índices. El índice de Shannon-Wiener tuvo un valor de 3,16 para la vegetación, lo cual indica una composición florística muy diversa de la comunidad, mientras que la diversidad del banco de semillas fue intermedia, con un valor de 2,16.
Comparación entre el banco de semillas y la vegetación. La determinación de la composición florística del banco de semillas tuvo dificultades por tratarse en su mayoría de especies de porte herbáceo, lo cual obligó a caracterizarlas como morfo especies; sin embargo, a partir de esta información se encontró que las especies compartidas son muy pocas. En la composición de la vegetación sobresale el componente arbóreo, seguido por arbustos y lianas. En el banco de semillas, la única especie de árbol encontrada fue el roble de tierra fría (Quercus humboldtii). La vegetación y el banco de semillas tuvieron un número muy bajo de especies comunes; además, la vegetación de porte arbóreo y arbustivo del bosque estuvo muy poco representada en el banco de semillas y esto se refleja en los índices. El índice de Jaccard mostró que la semejanza entre la vegetación y el banco de semillas alcanzó un valor de 1,5 % y un 3 % de similitud entre ambos componentes a partir del coeficiente de Sorenson.
Comparación entre el suelo y la hojarasca Son pocas las especies que comparten estos dos sustratos (Tabla 2) pues solo ocho morfo especies son comunes. De acuerdo con esto, los índices de Jaccard y de Sorenson alcanzaron valores muy bajos. Para el índice de Jaccard se obtuvo un 4,5 % de similitud entre el suelo y la hojarasca y con el coeficiente de Sorenson se alcanzó un 12,5 % de semejanza entre los mismos componentes.
Análisis de la distribución espacial. Los puntos con mayor densidad de líneas (F5, A6, C2, E1, E2) corresponden a aquellos que obtuvieron mayores resultados en la prueba de germinación (Figura 2). Las menores densidades de germinación se dieron en el centro de la parcela, con valores que oscilaron entre 0 y 10 semillas por punto muestreado, lo cual corresponde en el campo a un claro pequeño.
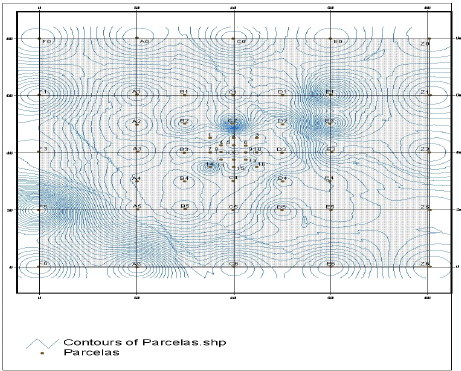
Figura 2. Diagrama de contornos de la distribución espacial del banco de semillas del suelo y la hojarasca ubicado en la cuenca de la quebrada Piedras Blancas, Antioquia-Colombia, de acuerdo con los resultados de emergencia de plántulas.
Las familias Lauraceae, Myrtaceae y Melastomataceae resultaron ser las más representativas en este estudio. Estos datos coinciden con los resultados de composición florística promedia de otros bosques altoandinos, en los cuales las Lauraceas son el elemento predominante, así como también se reporta la importancia de las familias Melastomataceae y Rubiaceae, (Gentry, 1995; 2001; Henderson, Churchill y Luteynm, 1991; Giraldo-Cañas, 1995). Los índices de riqueza y abundancia de especies indican que la vegetación es más diversa que el banco de semillas del suelo. La composición de la vegetación es más estable, mientras que al reservorio del suelo están entrando y saliendo continuamente individuos por factores como fructificación, germinación, muerte por depredación y cambios ambientales desfavorables (Garwood, 1989).
El mayor valor de emergencia de plántulas en las muestras de suelo (en los primeros 10 cm) es de 4.330 semillas m-2, comparado con las de hojarasca (198 semillas m-2). Esto se puede deber al efecto de la gravedad que acumula las semillas en los primeros centímetros del suelo, y es allí donde se presentan las mejores condiciones para que éstas puedan permanecer latentes y viables, al estar protegidas por la capa de hojarasca que las cubre. Las semillas con una ubicación más superficial (como las presentes en la hojarasca) son más susceptibles al ataque por depredadores y a los daños causados por factores ambientales (Skoglund, 1992; Dupuy y Chazdon, 1998).
Los valores de densidad promedio de semillas en el suelo encontrados en este estudio, son más altos que los reportados para bosques de montaña en el neotrópico, tal vez por la mayor profundidad de suelo que se muestreó. En bosques montano bajos de México en los primeros 5 cm de profundidad se encontraron 2.341 semillas m-2 (Williams-Lineira, 1993) y en bosques nublados 3.632 semillas m-2 (Alvarez-Aquino; Williams-Linera y Newton, 2005).
Gran parte de las semillas que se encontraban en el banco de semillas del robledal y que emergieron como plántulas pertenecen a especies de porte herbáceo, lo cual ya se había evidenciado en otros tipos de bosque tropical, como en bosques de rivera (Grombone-Guaratini, 1993) y bosques de niebla (Alvarez-Aquino; Williams-Linera y Newton, 2005). Estas especies herbáceas cumplen un papel importante en la regeneración del bosque (Skoglund, 1992). Además, algunas de ellas persisten en el sotobosque a través de varias etapas en el proceso de regeneración (Gómez-Pompa y Vázquez-Yanes, 1985). Esta mayor proporción de semillas de especies herbáceas, puede reflejar un ingreso excesivo de éstas provenientes de las áreas abiertas que rodean el fragmento.
La baja representatividad de especies arbóreas y arbustivas heliófitas en el banco de semillas estudiado, no parece deberse a condiciones ambientales adversas durante el tiempo de emergencia de las plántulas, puesto que el microclima del covertizo estaba entre los rangos adecuados para la germinación de este grupo de especies. Igualmente se considera que el tiempo de germinación fue suficiente para la emergencia de las plántulas heliófitas.
Las semillas de los árboles del bosque maduro, como el roble de tierra fría, forman parte del banco de semillas del suelo, solamente durante períodos muy breves de tiempo, ya que tienden a germinar o a ser depredadas muy rápidamente (Gómez-Pompa y Vásquez-Yanes, 1985). Su baja representatividad en la muestra, se puede atribuir a que la época en la cual se realizó el estudio, no coincidió con las épocas de fructificación de estas especies. Para tener una mejor representatividad de la composición de estas semillas que están temporalmente en el banco, sería necesario realizar muestreos en diferentes épocas del año. Es decir, se debería realizar un estudio de la dinámica poblacional del banco de semillas, ya que varios autores han informado sobre variaciones en la abundancia y composición del banco de semillas a lo largo del año (Butler y Chazdon, 1998; Grombone-Guaratini y Rodrígues, 2002). También es posible que en las muestras de suelo, hayan quedado semillas remanentes sin germinar y permanecieran en estado latente hasta el final del ensayo.
Por otro lado, los resultados obtenidos con el mapa de isolíneas de distribución espacial, muestran que existe una variación espacial significativa en la densidad de semillas del suelo a través del bosque (Figura 2). En general, la abundancia de semillas varía más que la riqueza de especies, entre diferentes puntos del bosque. Las mayores concentraciones en la densidad de semillas dentro del bosque correspondieron a zonas de hondonadas, pues estas condiciones topográficas del terreno favorecen el movimiento de semillas hacia esas zonas, con la ayuda de otros factores de arrastre como el viento y la escorrentía. La zona con menos densidad de semillas correspondió al centro de la parcela, en donde se ponía notar un claro pequeño en el dosel del bosque. Tal vez se trataba de un claro más grande que ya se estaba cerrando y cuyas condiciones microclimaticas habían activado la germinación de las semillas del banco hace un tiempo, y por esto su abundancia fue menor la abundancia de semillas en las muestras de este sector.
La composición de especies del banco de semillas de este fragmento de robledal, no reflejó la composición actual de la vegetación del bosque. El banco de semillas esta constituido principalmente por especies herbáceas y en menor proporción por arbustos, pero para tener una mejor representatividad de su composición es indispensable adelantar muestreos en diferentes épocas del año. También es posible que en las muestras de suelo, hayan quedado semillas remanentes sin germinar y permanecieran en estado latente hasta el final del ensayo.
La familia con mayor representatividad en la vegetación fue Lauraceae, seguida por Mirtaceae y Melastomataceae, lo cual concuerda con lo reportado para otros bosques altoandinos. Las especies más abundantes, más uniformemente distribuidas y de mayores tamaños fueron Quercus humboldtii, Alfaroa cf. colombiana y Myrcia popayanensis.
La hojarasca contiene poca cantidad de semillas (198 semillas m-2) y en los primeros 10 cm del suelo se acumula una gran cantidad de semillas viables (4.330 semillas m-2). Además existe una variación espacial significativa en la densidad de semillas del suelo a través del bosque a escala de unos pocos metros.
Este trabajo hace parte del proyecto de investigación Caracterización del banco de semillas en bosques de roble (Piedras Blancas, Ant.), cofinanciado por la Dirección de Investigaciones de la Universidad Nacional, Sede Medellín (DIME). El Departamento de Ciencias Forestales de la Universidad Nacional de Colombia, Sede Medellín suministró apoyo logístico con el personal y las instalaciones de la Estación Forestal de Piedras Blancas, el Laboratorio de Ecología y Conservación Ambiental y el laboratorio de Manejo Forestal. El Ingeniero Forestal Juan Carlos Penagos Zuluaga apoyó la identificación del material botánico en el herbario MEDEL de la Universidad Nacional de Colombia, Sede Medellín.
ALVAREZ-AQUINO, C.; WILLIAMS-LINERA, G. and NEWTON, A. C. Disturbance effects on the seed bank of mexican cloud forest fragments. En: Biotropica. Vol. 37, No. 3 (2005); p. 337- 348. [ Links ]
BERTILLER, M. B. Spatial pattern of the germinable soil seed bank in northern Patagonia. En: Seed Science. Vol. 8 (1998); p. 39-45. [ Links ]
BIGWOOD, D. W. and INOUYE, D. W. Spatial pattern analysis of seed bank: an improved method and optimized sampling. En: Ecology. Vol. 69: (1988); p. 497-507. [ Links ]
BUTLER, B. J. and CHAZDON, R. L. Species richness, spatial variation, and abundant of soil seed bank of a secondary tropical rain forest. En: Biotropica. Vol. 30, No.2 (1998); p. 214-222. [ Links ]
CAVELIER, J. and ETTER, A. Deforestation of montane forest in Colombia as a result of illegal plantations of opium (Papaver somniferum). En: CHURCHILL, S. P. et al., ed. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium. The New York Botanical Garden, 1995. p. 541-550. [ Links ]
________ and TOBLER, A. The effect of abandoned Pinus patula and Cupressus lusitanica on soils and regeneration of a tropical montane forest in Colombia . En: Biodiversity and Conservation. Vol. 7 (1998) ; p. 335-347. [ Links ]
CUBIÑA, A. and AIDE, T. M. The effect of distance from forest edge on seed rain and soil seed bank in a tropical pasture. En: Biotropica. Vol. 33, No.2 (2001); p. 321-326. [ Links ]
DALLING, J.; SWAINE, M. and GARWOOD, N. Effect of soil depth on seedling emergence in tropical soil seed – bank investigations. En: Functional Ecology. Vol. 9 (1994); p. 119-121. [ Links ]
________; ________ and ________. Soil seed bank community dynamics in seasonally moist lowland tropical forest, Panama. En: Journal of Tropical Ecology. Vol. 13 (1997); p. 659- 680. [ Links ]
________; ________ and ________. Dispersal patterns and seed bank dynamics of pioneer species in moist tropical forest. En: Ecology. Vol. 79 (1998); p. 564 – 578. [ Links ]
DUPUY, J. and CHAZDON, R. Long – term effects of forest regrowth and selective logging on the seed bank of tropical forest in NE Costa Rica. En: Biotropica. Vol. 30, No. 2 (1998); p. 223-237. [ Links ]
DUQUE, A. P. Estimación de la evapotranspiración por los modelos de Penman y Penman-Montheith en la cuenca de Piedras Blancas, Ant. Medellín, 1993. 215 p. Trabajo de Grado (Ingeniería Forestal). Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
ENRIGHT, N. Evidence of a soil seed bank Ander rain forest in New Guinea. En: Australian Journal of Ecology. Vol. 10 (1985); p. 67-71. [ Links ]
GARWOOD, N. Tropical soil seed banks: a review: En: ALLESIO, M.; PARKER, T. and SIMPSON, R., eds. Ecology of soil seed banks. United States of America : Academic Press, 1989. p. 149-204. [ Links ]
GENTRY, A. Patterns of diversity and floristic composition in neotropical montane forest: En: CHURCHILL, S. P. et al., ed. Proceedings of the Neotropical Montane Forest Biodiversity and Conservation Symposium. The New York Botanical Garden, 1995. p. 103-126. [ Links ]
___________. Patrones de diversidad y composición florística en los bosques de las montañas neotropicales: p 85-123. En: KAPELLE, M. and BROWN, A., eds. Bosques nublados del neotrópico. Costa Rica: Instituto Nacional de Biodiversidad, 2001. [ Links ]
GERHARDT, K. and HYTTERBORN, H. Natural dynamics and regeneration methods in tropical dry forests – an introduction. En: Journal of Vegetation Science. Vol. 3 (1992); p. 361-364. [ Links ]
GIRALDO-CAÑAS, D. Estructura y composición de un bosque secundario fragmentado en la cordillera central, Colombia: En: CHURCHILL, S. P. et al., ed. Proceedings of The Neotropical Montane Forest Biodiversity and Conservation Symposium. The New York Botanical Garden, 1995. p 159-167. [ Links ]
GOMEZ-POMPA, A. y VAZQUEZ-YANEZ, C. Estudios sobre la regeneración de selvas en regiones cálido - húmedas de México. En: GÓMEZ-POMPA, A. y DEL AMO RESTREPO, S., eds. Investigaciones sobre la regeneración de selvas altas en Veracruz. México D.F.: Alambra Mexicana De C.V., 1985. v. 2, p 1-26. [ Links ]
GROMBONE-GUARATINI, M. T. Banco de sementes de uma floresta riparia no Río Mogi Guacú, S P., 1993. 131 p. Master thesis. Universidad de Campinas, Campinas, SP. Institute of Biology [ Links ]
________ and RODRIGUES, R. R. Seed bank and seed rain in a seasonal semi-deciduos forest in south-eastern Brazil . En: Journal of Tropical Ecology. Vol.18 (2002); p. 759-774. [ Links ]
HENDERSON, A.; CHURCHILL, S. and LUTEYN, J. Neotropical plant diversity. En: Nature. Vol. 351 (1991); p. 21-22. [ Links ]
JARAMILLO, D. Reconocimiento de suelos con fines recreacionales y agropecuarios en la cuenca de la quebrada Piedras Blancas. Estudio realizado para la Dirección de Planeación de las Empresas Públicas de Medellín. Medellín- EPM, 1989. 67 p + mapas. [ Links ]
LORTIE, C. J. and TURKINGTON, R. The small-scale apatiotemporal pattern of a seed bank in the Negev desert, Israel . En: Ecoscience. Vol. 9, No. 3 (2002); p. 407-413. [ Links ]
MAGURRAN, A. Ecological diversity and its measurement. Princeton University, 1988. 179 p. [ Links ]
SAULEI, S. and SWAINE, M. Rain forest seed dynamics during succession at Gogol Papua, New Guinea . En: Journal of Ecology. Vol. 76 (1988); p. 1133-1152. [ Links ]
SINGHAKUMARA, B. M. P.; UDUPORUWA, R. S. J. P. and ASHTON, P. M. S. Soil seed banks in relation to light and topographic position of a hill dipterocarp forest in Sri Lanka . En: Biotropica. Vol. 32, No. 1 (2000); p. 190- 196. [ Links ]
SKOGLUND, J. The role of seed banks in vegetation dynamics and restoration of dry tropical ecosystems. En: Journal of Vegetation Science. Vol. 3 (1992); p. 357-360. [ Links ]
WILLIAMS-LINEIRA, G. Soil seed banks in four lower montane forests of México. En: Journal of Tropical Ecology. Vol. 9 (1993); p. 321-337. [ Links ]

