










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.59 no.1 Medellín Jan./June 2006
DEPRESIÓN EN VIGOR POR ENDOGAMIA Y HETEROSIS PARA EL RENDIMIENTO Y SUS COMPONENTES EN ZAPALLO Cucurbita moschata Duch. Ex Poir
INBREEDING DEPRESSION AND HETEROSIS FOR YIELD AND ITS COMPONENTS IN PUMPKIN Cucurbita moschata Duch. Ex Poir
Miguel Mariano Espitia Camacho1; Franco Alirio Vallejo Cabrera2 y Diosdado Baena García3
1 Profesor Asociado. Universidad de Córdoba. Facultad de Ciencias Agrícolas. Código Postal 354, Montería, Colombia. <mespitia@unicordoba.edu.co >
2 Profesor Titular. Universidad Nacional de Colombia, Sede Palmira. Facultad de Ciencias Agropecuarias. A.A. 0237. Palmira, Colombia. <favallejoc@palmira.unal.edu.co>,
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Palmira. Facultad de Ciencias Agropecuarias. A.A. 0237. Palmira, Colombia. <dbaenag@palmira.unal.edu.co>
Recibido: Septiembre 6 de 2005; aceptado: Febrero 26 de 2006.
RESUMEN
El estudio tuvo como objetivos estimar la depresión por endogamia (DE) y cuatro tipos de heterosis: promedio general (HP), promedio de progenitores (HM), heterobeltiosis (HB) y útil (HU), en dos dialélos [uno entre cinco variedades (V) y el otro entre cinco líneas endogamicas S1 (LS1), derivadas de éstas], para el rendimiento por planta (PFP), número de frutos por planta (NFP) y peso promedio de fruto (PPF). Los 30 genotipos se evaluaron en el municipio de Candelaria (Valle del Cauca, Colombia), en un diseño de bloques completos al azar con cinco repeticiones. Se presentaron diferencias significativas (p < 0,0l) para los tres caracteres, entre los cruzamientos (C) en los dos dialelos; mientras que la significancia estadística para progenitores (P) y P versus C, fue diferente en cada díalelo. En promedio las LS1 presentaron DE del 15,4 % (PFP), -4,6 % (NFP) y 20,3 % (PPF). La HP sólo fue significativa (p < 0,0I) para PFP (17,8 %) y NFP (-1,3 %) en el dialelo varietal; mientras que en el dialelo de LSI, lo fueron PFP (65,6 %) y PPF (35,4 %) con un valor p del 1 %. La HM, HB y HU tuvieron mayor expresión significativa (transheterosis) en el dialelo de LS1 (4,1 % a 65 %), que en el dialelo varietal (-12 % a 17,5 %), para los tres caracteres. Los híbridos LSO x LA34, LBV x LA126, LBV x LA34, LSO x LA126 y LBV x A88, presentaron los mayores niveles de HB (>40 %) y HU (>60 %) en PFP.
Palabras claves: Cruzamientos dialélicos, hibridación, heterosis, ahuyama.
ABSTRACT
With the goal of estimating inbreeding depression (DE) and the four types of heterosis: general average (HP), mean (HM), heterobeltiosis (HB) and standard (HU), in two diallelic [one among five cross pollinated varieties (VAR) and the other among five self-pollinated lines S1(LS1), derived from these], for yield/plant (PFP), number of fruits/plant (NFP) and weight/fruit (PPF). The 30 genotypes were evaluated using a complete randomized design with five replicates in Candelaria (Valle del Cauca, Colombia ). Significant differences (p < 0,01) occurred between crosses (C) for the three characters in both dialleles; whereas the statistical significance for parents (P) and P vs C, was different in each diallele. On average the LS1 presented inbreeding depression of 15,4 % (PFP), -4,6 % (NFP) and 20,3 % (PPF). The HP was significant ( p< 0,01) only for PFP (17,8 %) and NFP (-1, 3 %) in diallele VAR; whereas in the diallele of LS1 HP was significant for PFP (65,6 %) and PPF (35,4 %) to 1 % of probability. The HM, HB and HU had greater significant expression (trans-heterosis) in the diallele of LS1(4,1 % to 65 %) than in the diallele VAR (-12 % to 17,5 %) for the three characters. The hybrids LSO x LA34, LBV x LA126, LBV x LA34, LSO x LA126 and LBV x A88, showed the greatest levels of HB (>40 %) and HU (>60 %) in PFP.
Key words: Dialelic crossing, hybridization, heterosis, pumkin.
 MATERIALES Y MÉTODOS
MATERIALES Y MÉTODOS
RESULTADOS Y DISCUSIÓN
CONCLUSIONES
BIBLIOGRAFIA
El término heterosis fue propuesto por G.H. Shull para describir el vigor híbrido que se presenta en generaciones heterocigotas, derivadas del cruzamiento entre individuos genéticamente diferentes (Shull 1909).
La heterosis ha sido ampliamente utilizada en programas de mejoramiento de muchos cultivos para la identificación de poblaciones genéticamente divergentes como base para el desarrollo de líneas endogámicas para ser usadas en cruzamientos F1 (Hallauer y Miranda 1981).
No existe una teoría concluyente sobre la heterosis como agente causal del vigor híbrido (Birchler, Auger y Riddle 2003). Para explicar el fenómeno de heterosis se han formulado varias teorías y posiciones al respecto, a saber: 1) teoría de dominancia; 2) teoría de sobredominancia; 3) teoría epistática; 4) teoría de la acción conjunta de la dominancia y sobredominancia (Allard 1975) A las anteriores teorías se ha sumado también la metilación del ADN (Tsaftaris et al. 1997).
La heterosis puede ser expresada de diferentes formas, dependiendo del criterio usado para comparar el comportamiento de un híbrido: a) heterosis media (con base al promedio de los progenitores), b) heterosis útil (con base al promedio de un testigo estándar comercial) y c) heterobeltiosis (con base al promedio del mejor progenitor). Desde el punto de vista práctico, la heterosis útil es la más importante, porque permite desarrollar híbridos deseables superiores a los genotipos comerciales existentes en los sistemas de producción actual. Alam et al. 2004 señalan que es mejor expresar la heterosis no solo en comparación con los valores parentales sino también con un buen genotipo testigo, dado que la heterosis en los cruzamientos de variedades poco productivas no tiene interés comercial.
Miranda 1999, afirma que la expresión de la heterosis es menor en cruzamientos entre poblaciones de polinización abierta de base genética amplia. Varias publicaciones han señalado que el grado de endogamia, la dominancia unidireccional y la diversidad genética están estrechamente relacionados con la manifestación del vigor híbrido (Beck, Vasal y Crossa 1990, Crossa, Vasal y Beck 1990, Vasal et al. 1992 y Rezende y Souza Jr. 2000).
C. moschata, es una especie monoica de polinización cruzada natural. Posee altos niveles de heterocigosidad y variabilidad genética. Se han encontrado diversos niveles de manifestaciones de cis y transheterosis, en la mayoría de los caracteres agronómicos y de calidad del fruto (Doijode, Sulladmath y Kulkarini 1983, Doijode y Sullamath 1984, Berenji 1986, Shirohi, Kumar y Choudhury 1986, Galka 1987, Peixoto et al. 1988, Sirohi 1993, Amaya y Garza-Ortega 1996, Vallejo y Mosquera 1998, Vallejo y Gil 1998, Mohanty y Mohanty 1998, Mohanty, Mohanty y Mishra 1999, Mohanty y Mishra 1999, Mohanty y Prusti 2002 y Sirohi, Reddy y Behera 2002), con valores de heterosis (promedio, relativa, heterobeltiosis y útil) que han oscilado entre -53 % y 182 %.
Las investigaciones sobre los efectos depresivos por endogamia en cucúrbitas son muy antiguas y sus resultados son inconsistentes y diversos. Algunos autores registran disminución del vigor y expresión en varios caracteres agronómicos (Whitaker y Davis 1962, Peixoto et al. 1988, Giraldo y Vallejo 1989); mientras que otros no han encontrado depresión (Vallejo y Gil 1998).
En Colombia la importancia de C. moschata, es considerada como especie promisoria, teniendo en cuenta el área de siembra (3000 ha), producción (33.000 t), versatilidad en consumo directo, alimento saludable, materia prima para la agroindustria, artesanías, decoración, calidad nutricional, cultivo rústico, potencial de exportación, posible centro de origen, cultivos propios y ligados a pequeños agricultores (Espitia 2004). Actualmente se busca conocer el grado de heterosis, para las características agronómicas más importantes de las diferentes accesiones que conforman el banco de germoplasma de la Universidad Nacional de Colombia, Sede Palmira, con el fin de producir híbridos comerciales de zapallo de mayor rendimiento y calidad.
El estudio tuvo como objetivos específicos estimar la depresión por endogamia (DE) y cuatro tipos de heterosis: promedio general (HP), promedio de progenitores (HM), heterobeltiosis (HB) y útil (HU), en dos dialelos [uno entre cinco variedades (V) y el otro entre cinco líneas endogámicas S1 (LS1)], derivadas de éstas para el rendimiento por planta (PFP), número de frutos por planta (NFP) y peso promedio de fruto (PPF).
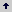
El trabajo se realizó durante el año 2003, en el Centro Experimental de la Universidad Nacional de Colombia, Sede Palmira (CEUNP), ubicado en el municipio de Candelaria (Valle del Cauca, Colombia). Según la clasificación de Holdridge, la zona pertenece a la formación Bosque Seco Tropical (BS-T) (Rodríguez 1999).
Se evaluaron dos dialélicos de zapallo (C. moschata) de cinco progenitores seleccionados, cada uno. El primero de ellos conformado por cinco variedades de libre polinización de diferente procedencia geográfica, a saber: Bolo Verde (BV), Soler (SO), Accesión 88 (A88), Accesión 34 (A34) y Accesión 126 (A126) y el otro por cinco líneas endogámicas S1 (LBV; LSO; LA88; LA34 y LA126), obtenidas por autofecundación de las anteriores variedades. En cada dialélico se evaluaron 15 genotipos (5 progenitores + 10 cruzamientos directos).
Se estimaron los caracteres rendimiento por planta (PFP), frutos por planta (NFP) y peso promedio de fruto (PPF). La evaluación agronómica de los 30 genotipos de los dos dialélicos se realizó en un sólo ensayo, mediante el uso del diseño de bloques completos al azar con cinco repeticiones. La distancia de siembra utilizada fue de 3 x 3 metros en cuadro. La unidad experimental efectiva consistió de un surco de seis plantas (54 m2). Los análisis estadísticos se realizaron con base en los promedios por unidad experimental.
La depresión endogámica (DE), heterosis promedio general (HP), heterosis con base al promedio de los progenitores (HM), heterobeltiosis (HB) y heterosis útil (HU), se estimaron a través de las siguientes formulas: DE = [(S0 - S1)/S0] x 100; donde S0 y S1, correspondieron al promedio para el carácter de interés en la variedad y su respectiva línea S1. HP = [(C/P) x 100] - 100; donde C = promedio de todos los cruzamientos con un progenitor común y P = promedio de ese progenitor. HM = [(F1/Xp) x 100] - 100; donde F1 = promedio de cada uno de los híbridos F1 y Xp = promedio de los dos progenitores. HB = [(F1/PMC) x 100] - 100; donde F1 = promedio de cada uno de los híbridos y PMC = promedio del progenitor de cada híbrido con mayor promedio HU = [(F1/TC) x 100] - 100; donde F1 = promedio de cada uno de los híbridos y TC = promedio del progenitor Unapal Bolo Verde (BV). La significancia estadística para los diferentes tipos de heterosis para cada cruzamiento, se probó mediante el uso de una prueba de t de Student.
En la Tabla 1, se muestra el ANDEVA para producción por planta (PFP) número de frutos por planta (NFP) y peso promedio de fruto (PPF). Las diferencias significativas (p < 0,01) entre genotipos sugieren la variabilidad genética en al menos uno de los 30 genotipos para los tres caracteres considerados. Al descomponer la fuente de variación genotipos en sus componentes, se observó que para PFP se presentaron diferencias significativas (p < 0,01) en todas las fuentes de variación consideradas, con excepción de los progenitores en los dos dialelos: variedades (V) y líneas S1 (L). El NFP, mostró diferencias estadísticas (p < 0,05) para todos las fuentes de variación, excepto para: líneas S1(L), los contrastes L vs HL y (V vs HV) versus (L vs HL). La variable PPF, excepto para el contraste V vs HV, presentó diferencias estadísticas significativas (p < 0,05) para el resto de las fuentes de variación.
Tabla 1. Cuadrados medios (CM) del análisis de varianza integral, promedios para el rendimiento por planta (PFP), número de frutos por planta (NFP) y peso promedio por fruto (PPF), considerando los dos dialelos. Candelaria (Valle del Cauca, Colombia). 2004.
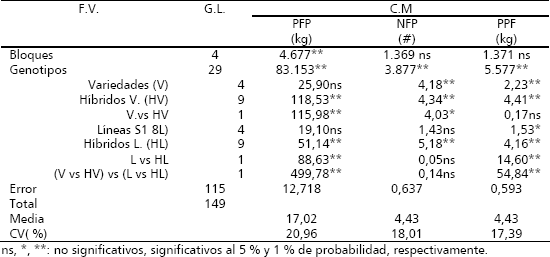
En la Tabla 2 se muestran por separado los promedios obtenidos para los progenitores del dialelo varietal (VAR) y de líneas S1 (LS1). Se puede detectar que en promedio las líneas SI presentaron menores valores, en comparación con las variedades a nivel de PFP (12,3 versus 14,6 kg) y PPF (3,4 versus 4,2 kg). Con algunas excepciones, los genotipos mejorados variedad Bolo Verde (6V) y Soler (SO) presentaron los mayores promedios para los tres caracteres considerados, señalando el efecto positivo del mejoramiento genético en tales cultivares, ya que estos son cultivares comerciales en Colombia y Puerto Rico, respectivamente.
Tabla 2. Promedios y depresión endogámica (DE) para el rendimiento por planta (PFP), número de frutos por planta (NFP) y peso promedio de fruto (PPF) de cinco varidades (VAR) y cinco líneas S1 (LS1) de zapallo (C. moschata). Candelaria (Valle del Cauca, Colombia). 2004.
Los menores promedios de las líneas S1, señalan el efecto depresivo que se produce como consecuencia de una mayor cantidad de loci homocigotos recesivos indeseables, a raíz de una generación de autofecundación en las variedades que las originaron. En promedio la depresión por endogamia (DE) tuvo su mayor expresión en PPF con 20,3 %, lo cual significó una reducción de 0,8 kg por fruto en las líneas, seguida por PFP con un valor de 15,4 %, el cual implicó una disminución del rendimiento de 2,3 kg por planta (Tabla 2). Contrario a las dos características anteriores, el NFP no se vio afectado significativamente por la depresión por endogamia, ya que su estimación fue de -4,6 %; ello representó una pequeña ganancia de 0,2 frutos por planta en las líneas S1. A nivel individual se puede notar que existen niveles de depresión diferentes entre las líneas S1 en cada carácter. Esto señala que cada una de las variedades progenitoras tenía una carga genética diferencial para cada variable, siendo mayor en A88 (LA88: DE = 25,2 %), A34 (LA34: DE = 23,6 %) y BV (LBV: DE = 23,6 %) para PFP; en A34 (LA34: DE = 24 %) para NFP y en, SO (LSO: DE = 36,7 %), A126 (LA126: DE = 3,3 %) y A88 (LA88: DE = 20 %) para PPF.
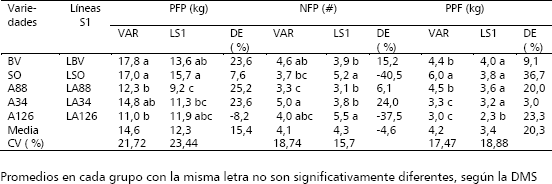
La tendencia general que se observa en tos resultados están de acuerdo con los obtenidos por otros investigadores (Witaker y Davis 1962, Vallejo y Gil 1998) y son contrarios a los reportados por Peixoto et al. 1988 y Giraldo y Vallejo 1989, quienes han encontrado que en las cucúrbitas la depresión por endogamia no es muy notable. Esta situación confirma la tendencia de los resultados contrastantes que se han obtenidos en los estudios de endogamia en cucúrbitas a nivel mundial. En relación a los dos grupos de híbridos (Tabla 3), se observó en forma general que las progenies F1 provenientes de las líneas SI (LS1) superaron en promedio a las obtenidas de las variedades (VAR), en 18,6 % para PFP y 25 % para NFP. Aún cuando los híbridos entre LS1 presentaron tendencia a producir frutos de menor peso (4,6 kg), en comparación con los híbridos derivados de VAR (4,9 kg), esta aparente desventaja, los puede hacer atractivos para el mercado de consumo en fresco, el cual prefiere frutos de tamaño pequeño a mediano (2 a 4 kg).
Tabla 3. Promedios y heterosis promedio (HP) para el rendimiento por planta (PFP), número de frutos por planta (NFP) y peso promedio de fruto (PPF) de diez híbridos (F1s entre cinco variedades (VAR) y diez entre cinco líneas S1 (VAR) de zapallo (C. moschata). Candelaria (Valle del Cauca, Colombia). 2004.
Entre los diez (10) híbridos de mayor comportamiento en los dos dialelos, sobresalen LSO x LA34 (26 kg); LBV x LA126 (23,7 kg); LBV x LA34 (22,7 kg); LSO x LA126 (21,9 kg); LBV x LSO (21,8 kg); LBV x LA88 (20,3 kg); BV x A34 (20,2 kg); BV x 50 (20,1 kg); SO x A34 (19,9 kg) y LSO x LA88 (19,4 kg). De este grupo de mayor producción siete (70 %) son híbridos entre LS1 y los tres restantes (30 %) son híbridos entre VAR, lo cual confirma el mejor comportamiento de los primeros sobre los segundos. Las variedades progenitoras que originaron los tres híbridos de mayor PFP, también dieron origen a líneas S1 que al cruzarse originaron tres de los híbridos de mayor PFP. Estos resultados permiten prever la existencia de niveles de heterosis y patrones heteróticos importantes para esta característica
De los nueve híbridos anteriores, sobresalen: LBV x LSO, LBV x LA34, LSO x LA126 y BV x A34, por presentar mayor NFP y PPF intermedio (4,4 a 4,5 kg/fruto), los cuales aún cuando no son los ideales (2 - 4 kg), si son aceptables para comercializarse para el mercado de fruto fresco. También se destacan los genotipos: BV x ABS, BV x SO, LBV x LA88, SO x A34 y LSO x LA88, por presentar además de buena producción y buen número de frutos/planta, los mayores promedios para peso/fruto, con valores entre 5,2 y 6,0 kg/fruto, convirtiéndose en posibles alternativas para el mercado agroindustrial, en el cual el tamaño y el peso/fruto, no es una característica limitante de importancia para su comercialización.
Los niveles de heterosis promedio (HP) estimados con base en la diferencia entre la media de todos los híbridos y sus progenitores en cada dialelo (Tabla 3), permiten observar mayor expresión de HP en el dialélico de LS1, en razón, a que sus estimaciones representaron 3,7, 16,3 y 2,3 veces, los obtenidos en el dialélico VAR, para PFP, NFP y PPF, respectivamente. Los mayores niveles de HP se presentaron en el carácter PFP, con incrementos del 17,8 % y 65,6 %, en los dialelos VAR y de LS1, respectivamente. Esto evidencia la presencia de importantes patrones heteróticos y altos niveles de heterosis a nivel de híbridos individuales en los dos dialelos.
Los resultados obtenidos en la HP para PFP y PPF, corroboran lo reportado en la literatura por Hallauer y Miranda 1981, Falconer y Makay 1996, Vallejo y Estrada 2002, quienes han señalado que en las progenies de líneas endogámicas, se expresa una mayor heterosis en relación con las progenies de poblaciones de base genética amplia (variedades de polinización abierta), siempre y cuando se presenten niveles importantes de divergencia genética entre los progenitores y existan niveles de dominancia unidireccional en la mayor cantidad de loci que controlan el carácter de interés en los progenitores, lo cual puede ser lo que ha ocurrido para los caracteres: PFP y PPF, pero no para NFP, en donde se presentó una situación un poco opuesta. Ello puede deberse a la ausencia de divergencia genética entre los progenitores y/o la existencia de niveles de dominancia no unidireccionales en los loci que controlan este carácter, los cuales llevan a que los locus con efectos de dominancia positiva se anulen con los de efectos negativos, generando una expresión nula o no detectable a nivel de las medias relacionadas con la heterosis promedio (Vencovsky y Barriga 1992, Pandini, Vello y Celis 2002 y Machado y Miranda 2003).
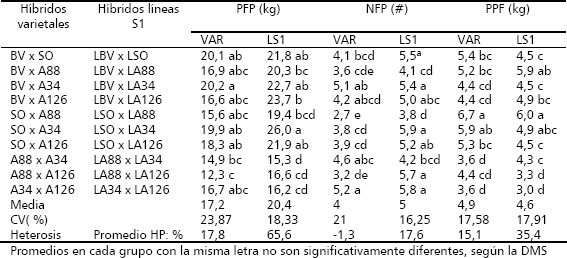
La expresión de los tres tipos de heterosis para los tres caracteres en estudio se presenta en el Tabla 4. La PFP fue de mayor magnitud y significancia estadística en los híbridos originados de líneas que en los provenientes de variedades. En el dialelo de líneas, la HM varió entre 39,8 % y 93 %, presentando todos los híbridos, valores estadísticamente significativos. En la heterobeltiosis (HB), el 70 % de los híbridos presentaron valores significativos, variando entre 39,3 % y 73,6 %. Para la heterosis útil (HU) el 70 % de los híbridos mostraron valores estadísticamente significativos, variando entre 42,7 % y 90,8 %. Esto contrasta con lo sucedido en el dialelo varietal en donde sólo un híbrido (A88 x A126) presentó HU significativa, pero negativa (-30,8 %). La presente situación corrobora lo detectado en el análisis de varianza (Tabla 1), prueba de medias (Tabla 2) y en la estimación de la heterosis promedio (Tabla 3).
Tabla 4. Heterosis con base al promedio de los padres (HM), al mejor padre (HB) y heterosis útil (HU), para la producción de frutos/planta (PFP), número de frutos/planta (NFP) y peso/fruto (PPF), en los cruzamientos F1 de los dialelos varietal (VAR) y de líneas S1, en zapallo (C. moschata). Candelaria (Valle del Cauca, Colombia). 2004.
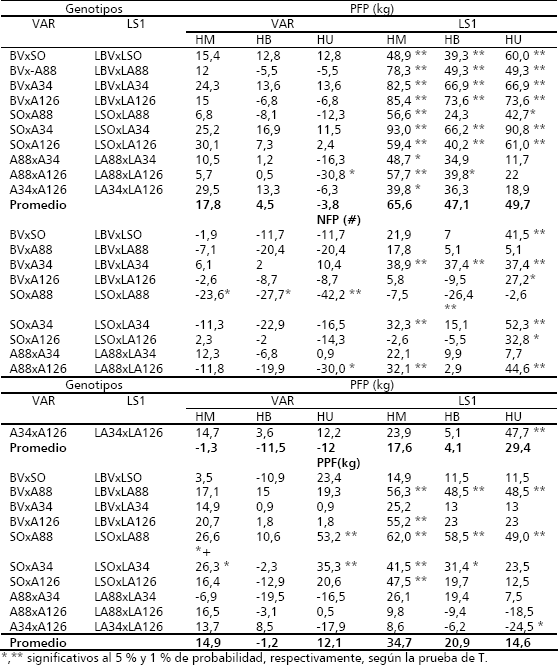
En general las mayores estimaciones de heterosis y niveles de significancia se presentan con base en el promedio de los progenitores (HM), con valores promedios de 17,8 % para el dialelo de VAR y 65,6 % para el de LS1.
Los híbridos entre líneas LSO x LA34 (90,8 %), LBV x LA126 (73,6 %), LBV x LA34 (66,9 %), LSO x LA126 (61 %) y LBV x LSO (60 %), sobresalieron por presentar los mayores niveles de expresión de HU. Casi todos estos híbridos también presentaron niveles altos de heterosis en relación con HM y HB. En general los híbridos que han mostrado mayores niveles de heterosis provienen del cruzamiento de líneas originadas de poblaciones mejoradas con líneas derivadas de poblaciones no mejoradas, lo cual puede sugerir una excelente complementación a nivel de diversidad genética entre los dos grupos de genotipos y un buen patrón heterótico entre ellos.
Los niveles de heterosis obtenidos para el carácter NFP, presentados en el mismo Tabla 4, confirman la superioridad heterótica de los híbridos de líneas en relación con los híbridos varietales. Para los híbridos varietales estas estimaciones fueron en su mayoría negativos (HM = -1,3; HB = -11,5 y HU = -12 %), las cuales contrastan con las obtenidas en promedio por los híbridos de líneas (HM = 17,6; HB = 4,1 y HU = 29,4 %) y a nivel individual en donde los híbridos con heterosis estadísticamente diferente de cero, presentaron valores positivos mayores de 32,1 %. Dentro de los híbridos con los más altos niveles de HU, sobresalieron: LSO x LA34 (52,3 %), LA34 x LA126 (47,7 %), LA88 x LA126 (44,6 %), LBV x LSO (41,5 %), LBV x LA34 (37,4 %) y LSO x LA126 (32,8 %). Cuatro de estos híbridos también se destacaron por sus altas expresiones de HU para PFP, son ellos: LSO x LA34, LBV x LA34, LSO x LA126 y LBV x LSO.
El PPF presentó tendencia en expresión de los tres tipos de heterosis, muy similar al observado en el NFP, en el sentido que la heterosis, tuvo su mayor expresión en los híbridos de LS1, en comparación con los de origen VAR, dado que los promedios en los primeros fueron de mayor magnitud, en HM (34,7 % vs 14,9 %), HB (20,9 % vs -1,2 %) y HU (14,6 % vs 12,1 %), con respecto a los segundos, En este carácter, la HM también presentó las mayores estimaciones de heterosis en los dos dialelos, en relación a HB y HU (Tabla 4).
Los híbridos que sobresalen por exhibir los mayores niveles de heterosis útil (HU), son: SO x A88, LSO x LA88, LBV x LA88, y SO x A34, con valores de 53,2, 49, 48,5 y 35,3 %, respectivamente. Estos híbridos también presentaron valores significativos de HM y HB. Es de resaltar, que ninguno de estos híbridos, presentó heterosis estadísticamente significativa para PFP y NFP, lo cual los hace deficientes en tales caracteres; no obstante, esta situación puede sugerir la existencia de un patrón heterótico específico para explotar la heterosis, cuando el objetivo sea incrementar el PFP, en un programa de mejoramiento para producir genotipos para el mercado agroindustrial.
Al comparar los niveles de heterosis en los tres caracteres de este grupo, se puede concluir con base en los resultados obtenidos, que los mayores niveles de expresión de heterosis, se presentaron en el carácter PFP, sobre todo en los híbridos provenientes de líneas S1, con valores promedios de 65,6 %, 47,1 % y 49,7 %, para HM, HB y HU, respectivamente. Una tendencia similar se observa en la expresión de la heterosis a nivel individual de los híbridos, ya que entre el 70 y 100 % de ellos, presentaron valores positivos y significativos, los cuales variaron entre 39,3 % y 93 %, en los tres tipos de heterosis estimados. Estimaciones de heterosis similares a nivel individual, han sido reportadas por Berenji 1986, Galka 1987, Peixoto et al. 1988, Sirohi 1993, Vallejo y Mosquera 1998, Vallejo y Gil 1998, Moharity y Mishra 1999, Mohanty et al.1999, Mohanty y Prusti 2002, Sirohi et al. 2002, trabajando con líneas endogámicas F4, F5 ó F6; lo cual señala que la explotación de la heterosis a nivel de la PFP y sus componentes puede ser una alternativa real y viable en el mejoramiento genético que busca incrementar la competitividad de la producción en C. moschata.
- En promedio las líneas S1 presentaron su mayor depresión endogámica, en PPF (20,3 %) y en PFP (15,4 %); resultando genéticamente diferentes a las variedades, a nivel de promedios.
- La HP sólo fue significativa (p < 0,01) para PFP (17,8 %) y NFP (-1,3 %) en el dialelo VAR; mientras que en el dialelo de LS1, lo fueron PFP (65,6 %) y PPF (35,4 %) al 1 % de probabilidad, señalando una mayor expresión en el dialelo de LS1.
- La HM, HB y HU tuvieron su mayor expresión significativa (transheterosis) en el dialelo de LS1 (4,1 % a 65 %), que en el dialelo varietal (-12 % a 17,5 %), para los tres caracteres.
- Los híbridos LSO x LA34, LBV x LA126, LBV x LA34, LSO x LA126 y LBV x A88, sobresalieron por presentar los mayores niveles de HB (>40 %) y HU (>60 %) en PFP.
- Se sugiere evaluar nuevamente los híbridos anteriormente mencionados, en varias localidades y semestres para confirmar su comportamiento y estimar su adaptabilidad y estabilidad.
Alam, M. F., Khan, M. R., Nuruzzaman, M., Parvez, S., Swaraz, A. M., Alam, I. and Ahsan, N. 2004. Genetic basis of heterosis and inbreeding depression in rice (Oryza sativa L.). En: Journal Zhejiang University Science. Vol. 5, no. 11; p, 406-411. [ Links ]
Allard, R. 1975. Principios de la mejora genética de las plantas. 2 ed. Barcelona: Omega. 498 p. [ Links ]
Amaya, A. T. and Garza-Ortega, S. 1996. High yields of summer squash lines and hybrids combinations. En: Cucurbit Genetic Cooperative. Report No.19; p. 78-80. [ Links ]
Beck, D. L., Vasal, S. K. and Crossa, J. L. 1990. Heterosis and combining ability of CIMMYTs tropical early and intermediate matury maize germoplasm. En: Maydica. Vol, 35; p. 279-285. [ Links ]
Berenji, J. 1986. Hybrid vigor of naked seeded oil pumpkin, Cucurbita pepo L. En: Uljarstvo. Vol. 23, no. 1; p. 79-85. [ Links ]
Birchler, J. A., Auger, D. L. and Riddle, N. C. 2003. In search of the molecular basis of heterosis. En: The Plant Cell. Vol. 15, no. 5; p. 2236-2239. [ Links ]
Crossa, J., Vasal, S. K. and Beck, D. L. 1990. Combinig ability in diallel crosses of CIMMYTs tropical late yellow maize germoplasm. En: Maydica. Vol. 35; p. 273-278. [ Links ]
Doijode, S. D. and Sulladmath, U. V. 1984. Preliminary studies on heterosis in Pumpkin (C. moschata Poir). En: Mysore Journal of Agricultural Sciences. Vol. 18, no. 1; p. 30-34. [ Links ]
________, ________ and Kulkarini, R. S. 1983. Heterosis for certain seed characters in pumpkin (C. moschata Poir). En: Indian Journal of Heredity. Vol. 15; p. 8-13. [ Links ]
Espitia, M. 2004. Estimación y análisis de parámetros genéticos en cruzamientos dialélicos de zapallo (Cucurbita moschata Duch. Exp Poir). Palmira. 206 h. Tesis de Doctorado. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
Falconer, D. S. and Mackay, T. 1996. Introduction to quantitative genetics. 4 ed. New York: Prentice Hall, 464 p. [ Links ]
Galka, A. T. 1987. Inheritance of some economically useful traits in first generation hybrids of Cucurbita. En: Ovoshchevodstvo-i-Bakhche-vodst-vo. Vol. 32; p. 45-46. [ Links ]
Giraldo, J. y Vallejo, F. A.1989. Modificación del hábito de crecimiento rastrero del zapallo (Cucurbita spp), mediante la transferencia del gen Bu (hábito arbustivo). En: Acta Agronómica. Vol. 39, no. 1-2; p. 19-24. [ Links ]
Hallauer, A. R. and Miranda, J. B. 1981. Quantitative genetics in maize breeding. Iowa State University Press, Ames, Iowa. 468 p. [ Links ]
Machado, R. and Miranda, J. B. 2003. Heterosis expression in crosses beween maize populations: ear yield. En: Scientia Agricola. Vol. 60, no. 3; p. 519-524. [ Links ]
Miranda Filho, J. B. 1999. Inbreeding and heterosis. p. 69-80. En: International Symposium, CIMMYT (1997: México) The genetics and exploitation of heterosis in crops. Proceedings. Madison: Crop Science Society of America . [ Links ]
Mohanty, B. K. and Prusti, A. M. 2002. Heterosis and combining ability for polygenic characters in pumpkin. En: Indian Agriculturist. Vol. 46, no. 3/4; p. 27-36. [ Links ]
Mohanty, B. K and Mishra, R. S. 1999. Heterosis for yield and yield components in pumpkin (Cucurbita moschata Duch. ex. Poir.). En: Indian Journal of Genetics and Plant Breeding. Vol. 59, No. 1; p. 505-510. [ Links ]
________ and Mohanty, S. K. 1998. Genetics of certain flowering characters in Cucurbíta moschata Duch. Ex Poir. En: Orissa Journal of Horticulture. Vol. 26; p. 1-3. [ Links ]
Mohanty, B. K., Mohanty, S. K. and Mishra, R. S. 1999. Genetics of yield and yield components in pumpkin (Cucurbita moschata Duch. Ex Poir). En: Indian Journal of Agriculture Sciences. Vol. 69; p. 781-783. [ Links ]
Pandini, F., Vello, A. N. and Celis, D. A. 2002. Heterosis in soybeans for seed yield components and associated traits. En: Brazilian Archives of Biology and Technology. Vol. 45; p. 401-412. [ Links ]
Peixoto, N., Casali, W. D., Cruz, D. O., Regazzi, J. A. and Seraphin, C. J. 1988. Efeito heterótico em híbridos braquiticos de abóbora Cucurbita moschata Dúchense do grupo baianinha. En: Horticultura Brasileira. Vol. 6, no. 2; p. 9-13. [ Links ]
Rezende, G. S. P. and Souza Junior, C. L. 2000. A reciprocal recurrent selection procedure outlined to integrate hybrid breeding program in maize. En: Journal of Genetics & Breeding. Vol. 54; p. 57-66. [ Links ]
Rodríguez, O. A. 1999. Efecto de cuatro métodos de labranza sobre las propiedades físicas de un suelo vertisol ústico y sobre la producción de Sorghum bicolor en el Valle del Cauca. Palmira. 147 h. Trabajo de grado Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
Sirohi, P. S. 1993. Genetic diversity in cucurbits pumpkin (Cucurbita moschata Duch. ex Poir). En: Indian Horticulture. Vol. 38, no. 4; p.35-37. [ Links ]
Sirohi, P. S., Reddy Y. S. and Behera, T. K. 2002. Heterosis for yield and its attributing traits in pumpkin (Cucurbita moschata Duch. ex Poir.). En: Journal of Vegetable Science. Vol. 29; p. 178-179. [ Links ]
________, Kumar, T. S. and Choud- hury, B. 1986. Studies on combining ability in pumpkin (Cucurbita moschata Ducheen ex Poir). En: Indian Horticulture. Vol. 25; p.1-17. [ Links ]
Shull, G. H. 1909. A pure line method of corn breeding. En: American Breeders' Association Report. Vol. 5; p. 51-59. [ Links ]
Tsaftaris, A. S., Kafka, M., Polidoros, A. and Tani, E. 1997. Epigenetic changes in maize DNA and heterosis. p. 112-113. En: Abstracts of the International Symposium on The Genetic and Exploitation of Heterosis in Crops. (1997: México City, México). Abstracts. México, D.F.: Centro Internacional de Mejoramiento de Maíz y Trigo, CIMMYT. [ Links ]
Vallejo, F. A. y Mosquera S., E. 1998. Transferencia del gen Bu a poblaciones de zapallo. Cucurbita sp. con crecimiento postrado. En: Acta Agronómica. Vol. 48, no. 3/4; p. 7-18. [ Links ]
Vallejo, F. A. y Estrada, E. 2002. Mejoramiento genético de plantas. Palmira: Feriva. 402 p. [ Links ]
Vallejo, F. A. y GIL, V. O. 1998. Efecto de la endogamia sobre algunos caracteres agronómicos del zapallo Cucurbíta moschata Poir. En: Acta Agronómica. Vol. 48, no. 3/4; p. 46-50. [ Links ]
Vasal, S. K., Srinivasan, G., Beck, D. L., Crossa, J. L., Pandey, S. and De Leon, S. 1992. Heterosis and combining cv ability of CIMMYTs tropical late white maize germoplasm. En: Maydica. Vol. 37, no. 1; p. 217-223. [ Links ]
Vencovsky, R. and Barrica, P. 1992. Genética biometrica no fitomelhoramento. Sao Paulo, Brasil: Sociedade Brasileira de Genética. 256 p. [ Links ]
Whitaker, J. and Davis, G. 1962. Cucurbits: botany, cultivation and utilization. New York: Interscience. 2510 p. [ Links ]

