










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.59 no.2 Medellín July/Dec. 2006
PROPAGACIÓN POR ESTACAS JUVENILES DEL BALSO BLANCO (Heliocarpus americanus L. Sin. H. popayanensis) UTILIZANDO PROPAGADORES DE SUBIRRIGACIÓN
PROPAGATION OF WHITE BALSUM (Heliocarpus americanus L. Sin. H. popayanensis) CUTTINGS USING NONMIST PROPAGATORS
Carolina Vásquez Restrepo1; Ana María Gutiérrez Uribe2 y Jorge Iván Álvarez González 3
1 Ingeniera Forestal. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <cvr48@hotmail.com>
2 Instructora Asociada, Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <amgutieru@unal.edu.co>
3 Profesor Asociado. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <jialvare@unal.edu.co>
Recibido: Junio 14 de 2005; aceptado: Julio 11 de 2006.
RESUMEN
El mercado mundial de edulcorantes orgánicos es una oportunidad para la panela, producto básico de la canasta familiar que representa ingresos importantes para la población rural colombiana. El balso blanco (Heliocarpus americanus L. Sin. H. popayanensis Hook & Arn.) es la especie más usada en procesos de clarificación de la panela en Antioquia y las zonas cañeras húmedas colombianas. Pero la extracción de la corteza de árboles obtenidos de la regeneración natural comienza a ser insostenible, por el daño causado a los individuos y la presión creciente a este recurso. La prohibición del uso de sustancias químicas en procesos de clarificación de panela ha aumentado la demanda de la corteza del balso blanco. En este trabajo se estimó el enraizamiento de estacas juveniles de balso blanco, por propagadores de subirrigación. Se realizaron dos experimentos, utilizando ácido anaftalenacético (ANA). En el primero se evaluó el efecto del transporte, cicatrizante y sustrato sobre el enraizamiento de las estacas de balso blanco. El mejor medio de transporte fue en cristales de hidrogel para mantener la humedad de las estacas, sin utilizar cicatrizante y sembrándolas en el sustrato tierra (55 % de enraizamiento). En el segundo se analizó la influencia de la intensidad lumínica y el área foliar en el porcentaje de enraizamiento de las estacas. El más alto enraizamiento se obtuvo con el tratamiento de doble sombra y un área foliar de 20 cm2 (25 % de enraizamiento). Aunque los mejores resultados indican un relativo éxito en el uso de medios de enraizamiento, es una primera aproximación para propagar esta especie que necesita ser protegida.
Palabras claves: Ácido anafatalenacético (ANA), área foliar, cristales de hidrogel, irradiación, sustratos, intensidad luminosa.
ABSTRACT
The world market of organic edulcorants is an opportunity for panela, a basic domestic consumption product that provides important income to rural population. White balsum (Heliocarpus americanus L. Sin. H. popayanensis Hook & Arn.) is the most used species in the panela clarification processes in Antioquia and in the humid Colombian sugar cane zones. But the extraction of bark from naturally regenerated trees eventually becomes unsustainable, due to the damage caused to individuals and the increasing demand for this resource. The prohibition of using chemical products in the panela clarification process has increasing the demand for white balsum bark. This research estimates rooting in softwood cuttings with the use of nonmist propagators. Two experiments were conducted using anaftalenacetic acid (ANA). The first experiment evaluated the effects of transportation, cicatrizant and substrates on the rooting of white balsum cuttings. The best means of transportation was in hydrogel cristals to maintain the humidity of the cuttings, without using cicatrizant, and planting them in soil substrates (55% rooting). The second analyzed the influence of illumination intensity and foliar area on rooting in the cuttings. The highest rooting was obtained with double shadow treatment and a 20 cm2 foliar area (25% rooting). Although the best results only yielded average rooting percentages (55%), this provides a first approximation to propagating this species, that need be protected.
Key words: anaftalenacetic acid (ANA), foliar area, hydrogel cristals, illumination intensity, substrates.
 MATERIALES Y MÉTODOS
MATERIALES Y MÉTODOS
RESULTADOS Y DISCUSIÓN
CONCLUSIONES
AGRADECIMIENTOS
BIBLIOGRAFÍA
En Colombia, la industria de la panela produce 25 millones de jornales al año y es la segunda generadora de empleo a nivel rural, después del café. De esta actividad subsisten 350.000 familias campesinas (12 % de la población rural económicamente activa) y 30.000 comerciantes de forma indirecta (Ministerio de Agricultura y Desarrollo Rural 2003).
En el año 2001, Colombia fue el principal consumidor de panela con 19,6 kg/ persona/año, seguido de lejos por Myanmar (antes Birmania) con 10 kg/persona/año (FAO 2004). Las cifras del Ministerio de Agricultura y Desarrollo Rural 2003, informan de un consumo aparente de 33,4 kg/persona/año. Este edulcorante es, además, una fuente de vitaminas y minerales especialmente para el sector más pobre, que actualmente se estima en un 60 % de la población del país, aproximadamente. El consumo de la panela representa el 2,18 % del gasto de alimentos de la población colombiana (2,94 % en los estratos bajos y 1,6 % en los estratos medios). La prohibición del uso de sustancias con propiedades blanqueadoras de la panela, como el hidrosulfito de sodio (Ministerio de Salud, Resolución 2284 de 1995), ha estimulado el interés por la recuperación de especies nativas utilizadas en el proceso de clarificación o descachazamiento de los jugos de caña para la elaboración de panela.
La industria de la panela en Colombia utiliza el mucílago de la corteza de tallo y raíces de varias especies del orden de las Malvales para la clarificación de la panela. En muchas regiones del país se ha empleado tradicionalmente el balso blanco (Heliocarpus americanus Sin. H. popayanensis) (Figura 1), siendo la especie de más amplio uso en Antioquia.
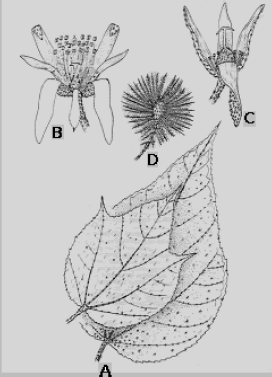
Figura 1. Heliocarpus americanus L. Sin. H. popayanensis. A. Hojas, envés; B. Flor hermafrodita; C. Flor femenina; D. Fruto (Robyns, 1964).
Esta especie se distribuye desde el sureste de México hasta noreste de Argentina, en una altitud entre 500 y 2.500 msnm en bosques secundarios (Robyns 1964). Es un árbol ginomonóico (flores femeninas o pistiladas y flores hermafroditas en la misma planta) (Croat 1978).
En regiones como el nordeste antioqueño la especie es escasa y por eso el 69 % de los trapiches deben hacer la clarificación con mezclas de balso blanco y otras especies. Ningún trapiche del nordeste antioqueño es autosuficiente en fuentes de mucílago, por lo que deben comprar todo o parte en el mercado (López y Osorio, 2003).
En la hoya del río Suárez (Boyacá, Santander, Colombia) se ha investigado la posibilidad de industrializar los polímeros que contiene la corteza del balso blanco, pero se requerirían de 26 toneladas de corteza para producir 2 toneladas al mes del extracto del polímero liofilizado ( La Rotta 1988) y aún no se ha evaluado la sostenibilidad de la producción.
López y Osorio (en imprenta) han realizado un estudio agronómico para evaluar la producción de los floculantes en algunas especies del orden de las Malvales, entre ellas el San Joaquín (Malvaviscus pendulifloreis Dc.) y algunas especies arvenses. Las evaluaciones con balso blanco tomarán más tiempo. Hasta el momento encontraron una germinación del 100 % en semillas del balso blanco, pero el enraizamiento de las estacas fue del 1 %.
Se ha demostrado que las estacas de árboles juveniles enraizan mejor que estacas de árboles adultos (varios árboles tropicales) Mesén 1998; Dalbergia sissoo Roxb., Fabaceae, Gupta, Kumar y Negi 1993; Tectona grandis Linn. F., Verbenaceae, Nautiyal, Sing y Gurumurti 1991; Pinus radiata Dist., Pinaceae, Penman 1988; Picea sitchensis (Bong.), Pinaceae, Kennedy y Selby 1984; Eucalyptus stuartiana F. Muell. ex Miq., E. gunni Hook., E. cinerea F. Muell. ex Benth., Myrtaceae, Curir y Sulis 1982; cacao, Sterculiaceae (Rendón 1952).
Aunque la regeneración natural del balso blanco es abundante, la producción vegetativa se justifica, porque los extractores de corteza de diferentes regiones de Colombia distinguen dos calidades de mucílago. Lo mismo ocurre en Costa Rica, donde el Heliocarpus appendiculatus Turcz. se le llama burío blanco o colorado, depeniendo del color del mucílago, que puede ser transparente o rojizo (Lay 1949). En ambos países se prefieren el mucílago transparente para los procesos de elaboración de la panela, pues flocula más rápidamente las impurezas. La propagación vegetativa sería un primer paso para establecer si estas diferencias son debidas a factores genéticos, fisiológicos o ambientales.
Esta investigación se plantea como hipótesis el que es posible propagar el balso blanco por estacas de árboles juveniles, utilizando una tecnología sencilla de propagadores de subirrigación, desarrollada en el Instituto de Ecología Terrestre de Escocia (ITE, por sus siglas en inglés), al alcance de cualquier campesino (Leakey et al. 1990).
Este trabajo se enmarcó dentro de los objetivos del Grupo de Investigación en Panela de la Universidad Nacional de Colombia, GIPUN, que pretende ser un Centro de Innovación Tecnológica para la mediana y pequeña industria de este sector.
Hace parte de la línea de conservación de los sistemas de producción de panela, al tratarse de obtener una fuente confiable de mucílago natural, propagada vegetativamente. Los resultados servirían para proponer su uso como especie cicatrizante después de un aprovechamiento forestal o en distintos arreglos agroforestales y no para el establecimiento de zonas de reserva, como lo propone el Instituto de Estudios Ambientales de la Universidad Nacional de Colombia, IDEA (González et al. 2004). Esta especie heliofita efímera requiere de claros grandes para su regeneración e impide la continuidad de la sucesión, por lo que no es recomendable mantenerla dentro del bosque (Peters 1994).
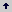
Para la recolección del material vegetal se realizaron dos visitas de inspección en diciembre de 2004 y enero de 2005 por la vía que conduce al municipio de Amagá y al Centro Nacional Agropecuario (SENA) La Salada, municipio de Caldas, departamento de Antioquia, Colombia. Estos sitios están ubicados a una altura aproximada de 1800 msnm, con una precipitación de 2444 mm anuales, correspondiente a la zona de vida del sistema de Holdridge bosque muy húmedo premontano (bmh-PM) (Espinal 1992).
Se buscaron 10 árboles juveniles, con diámetros menores de 10 cm, separados 100 m uno del otro, para garantizar la variabilidad genética y de los cuales se pudieran obtener al menos 50 estacas. Esto se basó en los criterios para estudios de semillas y procedencias, por tratarse de una especie monóica y de sitios con condiciones homogéneas (Kemp 1979). Los árboles fueron marcados con pintura y su diámetro y espesor de corteza fueron medidos a 1,30 m (d.a.p). Además se hicieron observaciones del mucílago en la corteza del tallo principal. Los árboles se establecieron por regeneración natural, a la orilla de carreteras y caminos, muy cerca de drenajes y quebradas. En diciembre de 2004 los árboles adultos presentaron botones florales y frutos verdes y en enero de 2005 se presentaron árboles con frutos verdes y maduros, mientras que los árboles juveniles no presentaron estos eventos. Esto coincide con el período de menor precipitación para la región.
Se utilizó una tecnología sencilla de propagadores de subirrigación (Figura 2) que ha sido probada con éxito en Centro América y África (Leakey et al. 1990), es barata fácil de adoptar, propone el uso de materiales disponibles localmente y puede usarse a pequeña o gran escala (Longman 1993).
Figura 2. Propagador de subirrigación. (Longman, 1993).
Los propagadores de subirrigación son un invernadero en miniatura, los cuales tienen la función de proveer de agua por capilaridad a los diferentes sustratos y evitar su evaporación. Para ello se forma un filtro. Éste consiste en una capa de 20- 25 cm de espesor formada con piedras (de 6 a 15 cm de diámetro), cubiertas con gravilla y una capa de 5 a 10 cm de sustrato desinfectado (Mesén 1998). Antes del llenado de los propagadores, se colocó un tubo de 2,5 pulgadas de diámetro y 50 cm de longitud verticalmente en la mitad del propagador, hasta sobresalir del sustrato.
Esto se hace para evitar erosión en el sustrato cuando se llena con agua por primera vez, o durante la realización de un experimento. Por último el filtro se llena de agua hasta la mitad, a través del tubo, y se observa el nivel de agua en los laterales del propagador. Los sustratos sólo se riegan abundantemente antes de la siembra y se procura mantener siempre el mismo nivel de agua.
Los propagadores se ubicaron en los predios de la Universidad Nacional de Colombia, Sede Medellín. Se soportaron sobre un lecho de aserrín y se protegieron con tela de sombrío del 65 % a 2 m de altura sobre el nivel del suelo. La recolección del material vegetal se hizo durante las primeras horas de la mañana y se sembraron lo más rápido posible para evitar su deshidratación y la cavitación (embolias) de los tejidos del xilema (Loach 1977).
En ambos experimentos, se trabajó únicamente con la primera estaca obtenida de cada rama, eliminando el ápice. Esto se hace para evitar la pudrición de estos tejidos meristemáticos (Mesén 1998).
En Triplochiton scleroxylon K. Schum. (Sterculiaceae), la capacidad de enraizamiento es máxima en la primera estaca subapical (Leakey 1985). Las estacas se trataron con ácido anaftalenacético al 0,4 % y se cortaron con una longitud de 20 cm. Los datos se evaluaron estadísticamente con el programa SAS® versión 8.2 (TS2M0), USA.
Efecto del sustrato, transporte y cicatrizante en el enraizamiento de estacas de balso blanco. Para analizar el enraizamiento se estableció un diseño completamente aleatorizado de efectos fijos. Los efectos fijos evaluados fueron dos condiciones de transporte del material vegetal (agua asperjada y cristales de hidrogel), aplicando o no un cicatrizante hormonal en la base y se sembraron en cuatro sustratos (arena, tierra, mezclas de arena:tierra y arena:aserrín descompuesto en proporción 1:1 en volumen). El cristal de hidrogel (Stockosorb®) es un copolímero de acrilamida y ácido acrílico como sal potásica. La arena se lavó con agua para eliminar los limos, arcillas e impurezas y se desinfectó con agua caliente; la tierra se zarandeó para eliminar el material grueso y las impurezas, además se desinfectó al cubrirla con aserrín que se mantuvo en combustión durante una hora. Se establecieron tres repeticiones que correspondían a tres propagadores. Cada propagador contenía 16 unidades de muestreo resultantes de la combinación de los factores analizados, distribuidos aleatoriamente.
Para asegurar la aleatoriedad de cada individuo se recolectaron 48 estacas de con 25- 30 cm de longitud, que se distribuyeron en partes iguales en cuatro neveras de icopor, en las cuales se aplicaron los siguientes tratamientos: cristales de hidrogel con la aplicación de cicatrizante en la base de cada estaca, cristales de hidrogel sin la utilización de cicatrizante, agua asperjada con la aplicación de cicatrizante en la base de cada estaca y agua asperjada sin el uso de cicatrizante.
La preparación de las estacas se hizo en el sitio donde se ubicaron los propagadores, aproximadamente una hora después de terminar la recolección, bajo sombra y manteniendo húmedo el material recolectado. La lámina foliar se redujo hasta 25 cm2 y se recortaron a20 cm de longitud eliminando completamente el cicatrizante de las estacas que lo tenían. En la base de las estacas se aplicó el ácido anaftalenacético (ANA) y se sembraron inmediatamente en el sustrato correspondiente. Se emplearon en total 160 estacas en cada repetición (propagador) y 480 para todo el experimento. Las hojas de las estacas y las paredes del propagador fueron asperjadas manualmente a las 9 a.m. y 3 p.m. diariamente. El periodo de análisis del experimento fue de seis semanas, haciendo una evaluación en la cuarta y sexta semana. Solamente se presentan los resultados de la sexta semana. El área donde se ubicaron los propagadores se protegió lateralmente con la tela de sombrío en los costados sur y occidente, además del sombrío superior permanente. Esto se hizo para evitar la alta radiación en las horas de la tarde por el ángulo de incidencia del sol. Los resultados obtenidos de este primer experimento indicaron la mejor forma de transporte de las estacas y el mejor sustrato de enraizamiento para realizar el segundo experimento.
Efecto del área foliar y la luz en el enraizamiento de estacas de balso blanco. Se analizó la influencia de la intensidad lumínica y el área foliar en el enraizamiento de las estacas de balso blanco, utilizando un diseño de bloques al azar con efectos fijos, donde la intensidad lumínica es el bloque.
Para evaluar la luminosidad se procedió a tapar al azar la mitad de los propagadores con tela de sombrío, ambos propagadores se protegieron por encima, con tela de sombrío a 2 m de altura para disminuir la luminosidad durante el medio día. Los propagadores se llenaron con tierra desinfectada con calor. De las 16 divisiones presentes en cada propagador, no se utilizaron las cuatro divisiones centrales, dos a cada lado del eje del propagador, para evitar efectos de borde por la incidencia de la luz solar en las diferentes horas del día. Se midió en la sexta semana la intensidad lumínica en µmol m-2 s-1 y la temperatura del aire en ºC con un (LI1000, LICOR, Lincoln, NE).
Se establecieron dos repeticiones para cada condición de luz. Para evaluar el efecto de área foliar se procedió con la disminución de la única lámina foliar a 0, 5, 10, 15, 20 y 25 cm2. Cada unidad de muestreo estuvo formada por 12 ó 13 estacas escogidas aleatoriamente de los mismos 10 árboles del primer experimento. Se recolectaron de 32 a 40 estacas por árbol y se transportaron en neveras de icopor con cristales de hidrogel, sin la adición de cicatrizante. Las estacas se cubrieron hasta 1 cm con ácido anaftalenacético (ANA) en talco y se sembraron inmediatamente en el sustrato tierra. Se emplearon 296 estacas en todo el ensayo.
El interés de propagar esta especie es la producción del mucílago a partir de la corteza. Las observaciones hechas en los árboles cosechados no mostraron diferencias en el mucílago. Al momento de hacer el corte se observó transparente y a los pocos minutos se oxidaba tomando una coloración rojiza.
Efecto del sustrato, transporte y cicatrizante en el enraizamiento de estacas de balso blanco. Al observar los resultados, dos de los propagadores mostraron un promedio de 22,5 % de enraizamiento en todos los tratamientos, mientras que uno de los propagadores mostró un promedio de 10,6 %, por lo cual se debió eliminar esta repetición.
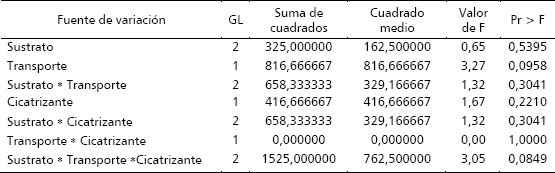
Así mismo, fue necesario descartar el nivel arena: aserrín puesto que no mostró ninguna variabilidad en su respuesta y además obtuvo un enraizamiento nulo. Con las modificaciones mencionadas, los factores explicaron en un 59,5 % el modelo evaluado para determinar la capacidad de enraizamiento y garantizan la normalidad de los residuales. Los resultados arrojados por el programa mostraron que estadísticamente el factor transporte del material vegetal (con cristales de hidrogel o con agua asperjada) fue significativo (p = 0,1) para el enraizamiento, y la triple interacción entre los factores sustrato, transporte y el cicatrizante fue significativa con un nivel de confianza mayor del 90 % (p = 0,09) (Tabla 1), siendo esta última la variable que más aportó al modelo.
Tabla 1. Análisis de varianza para el enraizamiento (%) de estacas de balso blanco (Heliocarpus americanus L.): efecto del sustrato, transporte y cicatrizante.
Debido a que la triple interacción fue significativa, y por jerarquía, para escoger el mejor tratamiento se recurrió a la interacción triple entre los factores sustrato, transporte y cicatrizante. En la Figura 3a se observa que la mejor respuesta se obtiene con el sustrato tierra, utilizando cristales de hidrogel como medio de transporte y sin la adición de cicatrizante en el material vegetal del que se obtuvieron las estacas (55 % de enraizamiento).
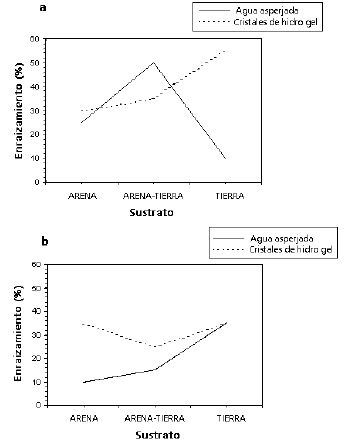
Figura 3. Respuesta del enraizamiento (%) del balso blanco Heliocarpus americanus L. debida a la interacción triple entre los factores sustrato, transporte y cicatrizante. a. Sin adición de cicatrizante, b. Con adición de cicatrizante.
En la Figura 3b no se muestran diferencias entre los sustratos arena y tierra, con la adición de cicatrizante (35 % de enraizamiento).
De los resultados de este experimento se deduce que hubo una mejor respuesta en el enraizamiento de estacas del balso blanco (55 %) frente al 1 % obtenido por López y Osorio (2003). Esta diferencia se explica porque se utilizaron estacas de árboles juveniles, de máximo un centímetro de diámetro y porque fueron estacas subapicales, que se espera tengan mayor capacidad de enraizamiento (Leakey 1985 y Mesén 1998).
Sin embargo, a nivel comercial un enraizamiento por debajo del 70 %, no se considera adecuado para ninguna especie (Leakey 1987).
En Heliocarpus appendiculatus el mejor enraizamiento (46,6%) se obtuvo en sustrato arena aserrín utilizando ácido indol-3-butírico (AIB) (Gutiérrez 2003). En el experimento con balso blanco el enraizamiento obtenido con el sustrato arena aserrín fue nulo. Sin embargo, en ambos experimentos el sustrato no mostró un efecto importante sobre el enraizamiento. En otra especie del orden de las Malvales, como el cacao, el mejor sustrato de enraizamiento fue cáscara de arroz, tanto fresca como descompuesta (84,5 % y 86,5 % respectivamente) utilizando propagadores de irrigación tipo Trinidad (Otto 1954). Los sustratos orgánicos siempre han resultado mejores para el cacao, pero muchos horticultores prefieren sustratos inertes para evitar contaminación por agentes biológicos (Andersen 1986).
El transporte del material vegetal para fines de propagación es un aspecto crítico en cualquier especie. Por ejemplo, Rumbos 2003, atribuyó la disminución en la producción de algunos viñedos jóvenes en Grecia al transporte inadecuado de las estacas de uva. Al utilizar como medio de transporte la inmersión del material en agua, Gutiérrez 2003, encontró que esta situación hacía brotar en abundancia el mucílago de la especie H. appendiculatus, por esta razón era necesario limpiar la base de las estacas para poder aplicar la hormona. En los experimentos siguientes el material vegetal fue empacado en bolsas y asperjados con agua durante la recolección y preparación de las estacas.
Debido a los resultados obtenidos por Gutiérrez 2003, en el transporte de estacas, se quiso comparar el método transporte utilizando agua asperjada con el uso de los cristales de hidrogel, que es el método usado para el transporte de palmas en la Universidad Nacional de Colombia, sede Medellín. No se encuentra información para comparar esta forma de transporte de material vegetal usando los cristales de hidrogel, que demostró tener ventajas frente al transporte de estacas con agua asperjada. La casa productora recomienda su uso para evitar el estrés durante el transplante.
El cicatrizante normalmente se usa para que las yemas foliares formen ramas en lugares diferentes al sitio del corte. En el caso del experimento con balso blanco, se adicionó el cicatrizante en la base del material vegetal del cual se obtuvieron las estacas para impedir la salida del mucílago y la deshidratación durante el transporte. Se retiró la zona que contenía el cicatrizante para que éste no ejerciera una barrera que impidiera el ingreso de la hormona en la estaca; al observar los resultados, el cicatrizante no tuvo ningún efecto en el enraizamiento.
La producción de mucílagos y látex dificultan la capacidad de enraizamiento, como ocurre con la especie Pouteria sapota, Sapotaceae (Lobato 1998). Durante la preparación del material no se observó la salida del mucílago en abundancia, sin embargo, este mucílago y la formación de un gran callo en respuesta al corte, pudieron haber sido un impedimento para la formación de raíces (Longman 1993 y Loach 1977).
En el corte transversal se apreció que la médula ocupa la mayor parte del área. La presencia de parénquima en la médula, la corteza y el xilema pueden provocar la formación de un callo abultado antes de la desdiferenciación de células, que no siempre resulta en la formación de raíces en estas especies (Flores 1999, Hartmann et al. 1997 y Hackett 1988). Aunque MacDonald 1990, encontró que un medio poco aireado tiende a producir formaciones callosas que emergen de las lenticelas en la parte inferior de las estacas, en el balso blanco se encontró la formación de callo en todos los tratamientos, característica propia de especies de difícil enraizamiento. Los sustratos con arena presentaron las formaciones callosas más grandes, a pesar de que fueron los sustratos más aireados. Al final del experimento con balso blanco, las estacas que presentaron callos continuaron con sus funciones metabólicas, pero es probable que una vez se terminen las reservas de sus tejidos mueran (Mesén, Newton y Leakey 1987).
En especies no leñosas, se observa que un mejor enraizamiento depende de una interacción entre estacas provenientes de plantas juveniles y que crezcan bajo sombra (Andersen 1986). En Triplochiton scleroxylon (Sterculiaceae) el porcentaje de enraizamiento se mejora cuando las plantas madres crecen bajo poca irradiación solar, al parecer porque bajo estas condiciones el contenido de almidones es bajo (Leakey 1985).
En general, las hojas de sombra tienen bajo contenido de almidones y la respiración se reduce, pero la fotosíntesis y el crecimiento son más lentos (Salisbury y Ross 1994). Los árboles de los cuales se obtuvieron las estacas para este experimento se encontraban a plena exposición, lo cual pudo afectar los resultados de enraizamiento.
Efecto del área foliar y la luz en el enraizamiento de estacas de balso blanco. Los factores intensidad lumínica y área foliar explicaron en un 77 % el modelo evaluado para determinar la capacidad de enraizamiento. Los datos se comportaron normalmente (p = 0,054). Los resultados arrojados por el programa mostraron que estadísticaente el factor área foliar y la doble interacción entre los factores intensidad lumínica y área foliar fueron significativos con un nivel de confianza mayor del 95 % (p < 0,05) (Tabla 2), siendo la interacción, la variable que más aportó al modelo.
Tabla 2. Análisis de varianza para el Área foliar * Sombra de enraizamiento de estacas de balso blanco Heliocarpus americanus L.: efecto de la intensidad lumínica y el área foliar.

Por jerarquía, aunque el factor intensidad lumínica (bloques) no aportó variación en la respuesta debe considerarse porque la interacción de ambos factores analizados fue significativa. El porcentaje más alto de enraizamiento se obtuvo en el tratamiento con doble sombra y un área foliar de 20 cm2 (25 %), los demás tratamientos presentaron un comportamiento muy similar (inferior al 10 %) (Figura 4).
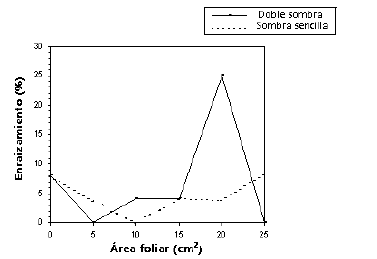
Figura 4. Respuesta del enraizamiento (%) del balso blanco Heliocarpus americanus L. debida a la interacción doble entre los factores intensidad lumínica y área foliar.
Para el cacao, las estacas enraizan mejor si se recolectan en épocas de sequía (Rendón 1952). En este experimento el enraizamiento se redujo más de la mitad en comparación con el primero (55 %), esto puede ser debido al inicio de la época de lluvias al momento de recolectar las estacas del segundo experimento, mientras que el primero se recolectó el material en época seca.
Los registros de temperatura y luz se hicieron durante 24 horas en los dos bloques y en ambas repeticiones, comenzando a las 10:35 de la mañana y terminando al siguiente día a la misma hora (Figuras 5 y 6). Se presentaron fluctuaciones considerables debidas al comportamiento climático, tiempo en el cual se presentaron días soleados y lluviosos. No se pudieron hacer promedios porque sólo se contaba con un sensor de luz y uno de temperatura para medir en cada tratamiento.
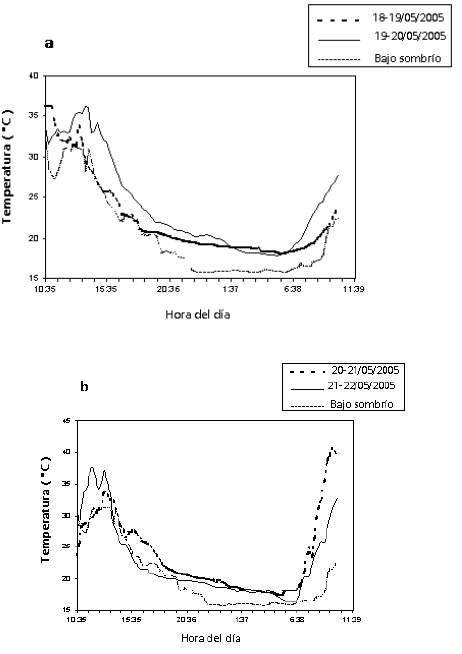
Figura 5. Comportamiento de la temperatura (ºC) bajo dos condiciones de luz en enraizadores de estacas de balso blanco Heliocarpus americanus L.N.C. 5a. Sombra sencilla, 5b. Doble sombra.
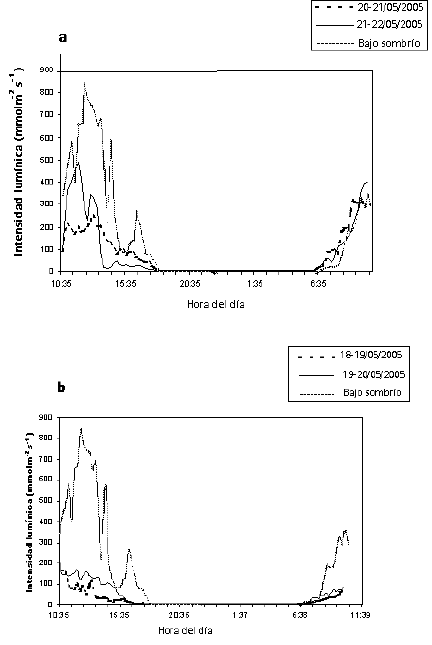
Figura 6. Comportamiento de la intensidad lumínica (µmol m-2 s-1) bajo dos condiciones de luz en enraizadores de estacas de balso blanco Heliocarpus americanus L. 6a. Sombra sencilla, 6b. Doble sombra.
La temperatura es el factor más crítico para la propagación (Hartmann et al., 1997). Para los propagadores de subirrigación ésta debe mantenerse en un rango de 28 a 33 ºC (Longman, 1993). Entre las 10:30 a.m. y 3:00 p.m. se presentaron temperaturas por encima de 30 ºC. En el tratamiento con sombra sencilla se observaron temperaturas máximas promedias de 40 ºC a las 10:20 a.m. en una de las repeticiones (Figura 5a), y aproximadamente a la misma hora se observaron temperaturas máximas promedias de 36 ºC en el tratamiento con doble sombra (Figura 5b). El tratamiento con doble sombra evita fluctuaciones rápidas en la temperatura dentro del propagador, mientras que en el tratamiento con sombra sencilla responde inmediatamente a un cambio en la temperatura externa al propagador y por debajo del sombrío.
La intensidad lumínica presentó los valores mayores al medio día (Figura 6). En un día soleado se reciben aproximadamente 2000 µmol m-2s-1 de radiación fotosintéticamente activa (RFA), al medio día (Hartmann et al. 1997). En el tratamiento con sombra sencilla se obtuvo un valor máximo promedio de 490 µmol m-2s-1 (Figura 6 a), mientras que en el tratamiento con doble sombra el valor máximo promedio fue de 168 µmol m-2s-1 (Figura 6 b). En el tratamiento con sombra sencilla, el sombrío utilizado más la lámina de polietileno que cubre los propagadores filtraron aproximadamente el 62 % de la luz, por lo tanto en un día soleado se podría recibir un máximo 760 µmol m-2s-1 al medio día.
En plántulas H. appendiculatus el punto de saturación lumínica se presentó a 500 µmol m-2s-1 de RFA (Fetcher et al. 1987). En un experimento de enraizamiento con esta misma especie, bajo dos condiciones de luz ensayadas, la mejor res puesta se obtuvo con la mayor radiación (máximas promedias de 476 µmol m-2s-1 de RFA) al medio día (Gutiérrez, Mesen y Sequeiro 2004). En el experimento con el balso blanco se obtuvo una intensidad lumínica cercana a 500 µmolm-2s-1 con sombra sencilla, pero este tratamiento se logró un menor porcentaje de enraizamiento.
Aunque en doble sombra dio mejor porcentaje de enraizamiento, las diferencias entre ambas condiciones de luz no fueron significativas. El mismo resultado se presentó en estacas de Cordia alliodora. Esto fue atribuido al comportamiento parabólico del enraizamiento como efecto de la cantidad de luz, donde las diferencias de irradiación podrían haber estado por encima o por debajo de las condiciones óptimas (Mesén 1993). Se recomienda ensayar valores intermedios cercanos a 400 µmol m-2s-1 de RFA, que es una radiación apropiada para la mayoría de las especies (Grange y Loach 1983), ya que las copas de la especie H. appendiculatus en condiciones naturales se orientan preferiblemente hacia la radiación difusa más bien que hacia la radiación directa (Ackerly y Bazzaz, 1995). En Tilia cordata, T. europaea, y T. tomentosa, familia Tiliaceae, se recomienda la exclusión de luz para mejorar el enraizamiento (Maynard y Bassuk, 1988).
En otro estudio con Cordia alliodora la interacción entre iluminación y área foliar produjo diferencias en el enraizamiento. Con poca iluminación son mejores las áreas foliares grandes, y con alta iluminación es mejor tener áreas foliares pequeñas (Mesén, Newton y Leakey 1997). Para el caso del balso blanco la interacción entre estos factores fue significativa, pero debido a la caída masiva de las hojas y a la alta mortalidad presentada en este experimento los resultados no permiten concluir cuál es la radiación óptima para las estacas de esta especie, pues la temperatura fue el factor de mayor peso y está estrechamente relacionado con la iluminación y el riego por aspersión que se debe realizar diariamente (Figura 4). El único caso en que se presenta esta diferencia es para el área foliar de 20 cm2 (25 % de enraizamiento), pero los demás tratamientos presentaron un enraizamiento similar o inferior al tratamiento de estacas sin hoja o con área foliar cero (8,33 %).
En el tratamiento de doble sombra y área foliar de 20 cm2 ocurrió el mejor porcentaje de enraizamiento, simplemente porque obtuvo el mayor porcentaje de sobrevivencia. En cada tratamiento se observó que la tendencia del enraizamiento fue la misma que la sobrevivencia.
De acuerdo a las observaciones fenológicas hechas al momento de obtener el material vegetal para el segundo experimento, los árboles presentaron una defoliación aproximada del 25 % del área de la copa. Esto puede explicar la caída masiva de las hojas al cuarto día de la siembra; o que pudo haber una interacción entre las condiciones ambientales y las fitohormonas presentes en las hojas (ácido absícico o etileno) que participan en este proceso (Hartmann et al. 1997).
La aspersión de las hojas busca disminuir la temperatura. Debido a que el sustrato tierra se observó demasiado húmedo, se tomó la decisión de hacer un solo riego a partir de la cuarta semana, lo que pudo afectar la expresión del enraizamiento. Estos resultados son el inicio para proponer sistemas que permitan hacer aproximaciones a sistemas sostenibles de aprovechamiento de una fuente natural de mucílago a partir de la corteza de árboles de balso blanco. Sin embargo, es necesario continuar con las investigaciones, ya que existen evidencias que el espesor de corteza en árboles de Pinus radiata provenientes de estacas es menor, comparado con árboles provenientes de semillas de la misma edad (Pennman 1988).
La mejor respuesta de enraizamiento se obtuvo utilizando cristales de hidrogel como medio de transporte sin la adición de cicatrizante y sembrando las estacas en el sustrato tierra, teniendo cuidado con la correcta desinfección del medio de enraizamiento.
Al evaluar el efecto del área foliar, el porcentaje más alto de enraizamiento se obtuvo con el tratamiento de doble sombra y un área foliar de 20 cm2, pero es necesario hacer ajustes que permitan encontrar condiciones de temperatura e intensidad lumínica óptimas para el Heliocarpus americanus L. Sin. H. popayanensis
Es necesario tener en cuenta la época de recolección del material vegetal, ya que en la época seca las estacas recolectadas duplicaron el porcentaje de enraizamiento. Aunque la formación de callo no es un buen criterio para seleccionar el mejor tratamiento, se observó que las estacas recolectadas en la época seca que no desarrollaron raíces, sobrevivieron.
Se requiere observar si existe alguna interacción entre las condiciones ambientales, en el momento de recolección y durante el desarrollo del experimento, que induzcan a la producción del callo, para tratar de corregir este problema y que se llegue a la formación de raíces.
El uso de propagadores de subirrigación, es una tecnología sencilla que permite propagar vegetativamente el balso blanco, pero es necesario seguir haciendo investigaciones para encontrar las condiciones ambientales óptimas, para la recolección y el enraizamiento de las estacas.
Para próximas investigaciones se recomienda controlar los niveles de temperatura y radiación, teniendo en cuenta las variaciones del ángulo de inclinación del sol durante el año.
Al Departamento de Investigación Sede Medellín (DIME) que económicamente fue el apoyo para el desarrollo de esta investigación. Al Laboratorio de Productos Forestales y al Laboratorio de Ecología y Conservación Ambiental (LECA), por el apoyo logístico prestado durante todo el desarrollo de los experimentos. A Diana Marcela Pérez Valencia, por su valiosa ayuda en los análisis estadísticos.
Ackerly, D. D. y Bazzaz, F. A. 1995. Seedling crow orientation and interception of diffuse radiation in tropical forest gaps. En: Ecology. Vol. 76, no. 4; p. 1134-1146. [ Links ]
Andersen, A. S. 1986. Environmental influences on adventitious rooting in cuttings of nonwood species: p. 223-253. En: Jackson, M., ed. New root formation in plants and cuttings. Dordrecht, Holanda: Martinus Nijhoff Publishers. 261 p. [ Links ]
Croat, T. B. 1978. Flora of Barro Colorado. Stanford University Press. CA. 943p. [ Links ]
Curir, P. and Sulis, S. 1982. Studio sulla rizogenesis in talee di Eucalyptus stuartiana, E. gunni, E. cinerea. En: Annali dell'Istituto Sperimentale per la Floricoltura. Vol. 13, no. 1; p. 63-78. [ Links ]
Espinal, L. S. 1992. Geografía ecológica de Antioquia: zonas de vida. Medellín: Lealon. 101 p. [ Links ]
FAO, 2004. Estadísticas de consumo de alimentos: sugar noncentrifugal Roma, Italia. FAO. Disponible en Internet http://apps.fao.org/faostat/servlet/XteServ let3?Areas=%3E862=y Domain=FSyItem Types=FS.CropsAndProductsylanguage=EN. [Consultada:13 ago. 2004.] [ Links ]
Fetcher, N., Oberbauer, S. F., Rojas, G. y Strain, B. J. 1987. Efectos del régimen de luz sobre la fotosíntesis y el crecimiento en plántulas de árboles de un lluvioso tropical de Costa Rica. En: Revista de Biología Tropical. Vol. 35, Supl.1; p. 97-110. [ Links ]
Flores Vindas, E. 1999. Parénquima y colénquima: p. 151-171. En: La planta. Estructura y función. Cartago, Costa Rica: Libro Universitario Regional. v. I y II. 367 p. [ Links ]
González Castrillón, E., Velásquez, H. I., Velásquez, H. C. y Álvarez, J. I. 2004. Evaluación preliminar del impacto ambiental en la producción de panela en Antioquia. Medellín: Universidad Nacional de Colombia, IDEA/GIA/GIPUN. S.p. 80 p. [ Links ]
Grange, R. I. and Loach, K. 1983. Environmental factors affecting water loss from leafy cuttings indifferent propagation systems. En: Journal of Horticultural Science. Vol. 58, no.1; p. 1-7. [ Links ]
Gupta, B. B.; Kumar, A. and. Negi, D. S. 1993. Vegetative propagation through braneh cuttings in Dalbergia sissoo Roxb. En: Indian Forester. Vol. 119, no.5; p. 381-387. [ Links ]
Gutiérrez Uribe, A. M. 2003. Propagación del burío (Heliocarpus appendiculatus Turcz.) por semillas, estacas y acodos. Turrialba, Costa Rica. 106 h. Tesis Maestría en Ciencias Forestales. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Departamento Manejo de Recursos Forestales. [ Links ]
________, Mesén Sequeiro, F. y Villalobos Soto, R. 2004. Propagación del burío (Heliocarpus appendiculatus Turcz.), un recurso no maderable del bosque tropical útil para la producción del dulce y azúcar orgánicos. En: Recursos Naturales y Ambiente. No. 41; p. 80-87. [ Links ]
Hartmann, H. T., Kester, D. E., Davis, F. T. and Geneve, R. L. 1997. Plant propagation: principles and practices. 6ed. New Jersey: Prentice Hall. 770 p. [ Links ]
Hackett, W. P. 1988. Donor plant maturation & adventitious root formation: p. 11-28. En: Davis, D. V.; Haissig, B. and Sankhla, N., eds. Adventitious root formation in cuttings. En: Advances in Plant Sciences Series. Portland. Oregon: Dioscorides Press. [ Links ]
Kemp, R. H. 1979. La obtención de semilla para las investigaciones de especies y procedencias. En: Burley, J. y Wod, P. J. Manual sobre investigaciones de especies y procedencias con referencia especial a los trópicos. Oxford, UK : Commonwealth Forestry Institute (Tropical Forestry Paper no. 10 y 10 A). [ Links ]
Kennedy, S. J. and Selby, C. 1984. Propagation of sitka spruce by stem cuttings. En: Record of Agricultural Research. Vol. 32; p. 61-70. [ Links ]
Küppers, M., Timm, H., Orth, F., Stergemann, J. Stöber, R. Schneider, H., Paliwal, K., Karunaichamy, K. and Ortiz, R. 1996. Effects of light environment and successional status on light fleck use by under story trees of temperature and tropical forest. En: Tree Physiology. vol. 16; p. 69-80. [ Links ]
La Rotta García, H. 1988. Extracción y conservación del polímero del balso. Bogotá. 104 h. Trabajo de grado Ingeniero Químico. Universidad Nacional de Colombia. Facultad de Ingeniería. [ Links ]
Lay, K. K. 1949. A revision of the genus Heliocarpus L. En: Annals of the Missouri Botanical Garden. Vol. 36, no. 4; p. 507-541. [ Links ]
Leakey, R. R. B. 1985. The capacity for vegetative propagation in trees: p. 110-133. En: Cannell, M. G. R. and. Jackson E., eds. Attributes of trees as crop plants. Midlothian, Scotland: Institute of Terrestrial Ecology. 315 p. [ Links ]
Leakey, R. R. B. 1987. Clonal forestry in the tropic: a review of developments, strategies and opportunities. En: Commonwealth Forestry Review. Vol. 66, no.1; p. 61-75. [ Links ]
Leakey, R. R. B., Mesén, J. F., Tchoundjeu, Z., Longman, K. A., Dick, Mcp. J., Newton, A., Matin, A., Grace, J., Munro, R. C. and Muthoka, P. N. 1990. Low technology techniques for vegetative propagation of tropical trees. En: Commonwealth Forestry Rewiew. Vol. 66, no.1; p. 61-75. [ Links ]
Loach, K. 1977. Leaf water potencial and the rooting of cuttings under mist and polythene. En: Physiology Plantarum. No. 40; p. 191-197. [ Links ]
Lobato Artiga, S. D. 1998. Desarrollo de métodos de propagación para la conservación y propagación ex situ de especies de Sapotáceaes: Pauteria sapota (Jacq). Turrialba, Costa Rica. 131 h. Tesis Maestría en Ciencias Forestales Centro Agronómico Tropical de Investigación y Enseñanza (CATIE). Departamento Manejo de Recursos Forestales. [ Links ]
Longman, K. A. 1993. Árboles tropicales: manuales de propagación. Enraizamiento de estacas de árboles tropicales. London, United Kingdom: Commonwealth Science Council. 131p. [ Links ]
López Osorio, J. G. y Osorio Cadavid, G. 2003. Evaluación y conservación de las especies aglutinantes o floculantes utilizadas en la agroindustria panelera de los municipios situados en jurisdicción de CORANTIOQUIA: informe parcial de resultados. 30 p. En: X Seminario Nacional y IV Internacional sobre especies promisorias: biodiversidad y bioprospección. (2003: Medellín, Colombia). Memorias. Medellín: IPGRI/ Universidad Nacional de Colombia, Sede Medellín. Corporación Autónoma Regional del Centro de Antioquia, CORANTIOQUIA – Corporación Colombiana de Investigación Agropecuaria, CORPOICA. [ Links ]
MacDonald, B. 1990. Practical woody plant propagation for nursery growers: rooting hormones. Portland, Oregon: Timber Press. 351 p. [ Links ]
Maynard, B. K. and Bassuk, N. L. 1988. Etiolation and banding effects on adventitious root formation. p. 29-46. In: Davis, T. D., Haissig, B. E. y Sankhla, N., eds. Adventitious root formation in cuttings. En: Advances in Plant Sciences Series, Portland, Oregon: Dioscorides Press. [ Links ]
Mesén, J. F. 1993. Vegetative propagation in Central America hardwoods. Scotland . 231 h. Ph. D. Thesis University of Edinburgh. [ Links ]
________, Newton, A. C. and Leakey, R. B. 1997. The effects of propagation environment and foliar area on the rooting physiology of Cordia alliodora (Ruiz & Pavon) Oken cuttings. En: Trees Structure and Function. Vol. 11, no. 7; p. 404-411. [ Links ]
________. 1998. Enraizamiento de estacas juveniles de especies forestales: uso de propagadores de subirrigación. Turrialba, CR.: CATIE. 36 p. (Serie Técnica, Manual Técnico no. 30). [ Links ]
Nautiyal, S., Sing, U. and Gurumurti, K. 1991. Rooting response of branch cuttings of teak (Tectona grandis) as influenced by season and growth hormones. En: Indian Forester. Vol. 117, no. 49; p. 249-255. [ Links ]
Otto Pérez, J. M. 1954. Experimento comparativo entre siete medios de enraizamiento para estacas de cacao. Palmira. 35 h. Trabajo de grado Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
Penman, J. T. D. 1988. Volume, taper and bark thickness in seedlings and cuttings from Mamaku forest, New Zealand . En: New Zealand Journal of Forestry Science. Vol. 18, no. 3; p. 311-317. [ Links ]
Peters, C. M. 1994. Sustainable harvest of nontimber plant resources in tropical moist forest: an ecological primer. Washington D. C.: WWF/The Nature Conservancy World Resources Institute. 45 p. [ Links ]
Rendón Araujo, P. 1952. Influencia de las estaciones y del estado de la estaca sobre el enraizamiento de estacas de cacao. Palmira. 42 h. Trabajo de grado Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. [ Links ]
Ministerio de Agricultura y Desarrollo Rural. 2003. La Cadena agroindustrial de la panela en Colombia: una mirada global de su estructura y dinámica 1991-2005 Observatorio Agrocadenas Colombia. Disponible en Internet: http://www.agrocadenas.gov.co/home. htm. [Consultada: 12 may. 2004]. [ Links ]
Ministerio de Salud. 1995. Resolución 2284 del 27 de junio de 1995: medidas sanitarias sobre la panela. Santafé de Bogotá D. C. Colombia: El Ministerio. 20 p. [ Links ]
Robyns, A. 1964. Flora of Panama : part IV, family 114. Tiliaceae. En: Annals of the Missouri Botanical Garden. Vol. 51, no. 1-4; p. 3-35. [ Links ]
Rumbos, I. C. 2003. Young grapevine decline. Associated with defective propagated material. En: Bulletin OILB/ SROP. Vol. 26, no. 8-1; p. 45-47. [ Links ]
Salisbury, F. B. y Ross, C. W. 1994. Fisiología Vegetal. México: Interamericana McGraw-Hill. 759 p. [ Links ]

