










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.59 no.2 Medellín July/Dec. 2006
PRODUCCIÓN Y CALIDAD DE FRUTA EN CULTIVARES DE FRESA (Fragaria sp.) AFECTADOS POR ESTRÉS SALINO
YIELD AND FRUIT QUALITY OF SALT-STRESSED STRAWBERRY CULTIVARS (Fragaria sp.)
Fánor Casierra-Posada 1 y Nehidy García Riaño2
1 Profesor Asociado. Universidad Pedagógica y Tecnológica de Colombia. Facultad de Ciencias Agropecuarias A.A. 661, Tunja, Colombia. <fcasierra@tunja.uptc.edu.co>
2 Bióloga, Universidad Pedagógica y Tecnológica de Colombia. A.A. 661, Tunja, Colombia. <nehidydgr@hotmail.com>
Recibido: Noviembre 17 de 2005; aceptado: Septiembre 29 de 2006.
RESUMEN
Dado que el agua es muy buen solvente, las aguas que se usan para riego contienen algunas sales disueltas. La salinidad restringe la disponibilidad de este líquido para las plantas mediante la reducción del potencial hídrico en el suelo. La salinidad también tiene un impacto sobre la fisiología de la producción de las plantas. Es posible observar lesiones en plantas expuestas a niveles altos de salinidad. Este ensayo fue realizado para evaluar la relación entre la salinidad por NaCl, la producción y la calidad de fruta, en los cultivares de fresa Chandler, Sweet Charlie y Camarosa cultivados en invernadero, en materas con suelo salinizado. Se compararon cinco niveles de la sal (0, 20, 40, 60 y 80 mM de NaCl) los cuales indujeron diferentes valores de conductividad eléctrica (EC) (2,68; 6,08; 8,24; 9,98 y 12,62 dS.m-1, respectivamente). Los diversos niveles de EC se obtuvieron agregando el NaCl al suelo, progresivamente. La producción y la calidad de la fruta fueron afectadas negativamente, por los tratamientos probados. De igual manera, el contenido de azúcares en los frutos fue influido por los diferentes niveles de sal. Los contenidos de sólidos solubles totales y la acidez titulable en frutas se incrementaron con el aumento en la EC. Mientras que la acumulación de la materia seca en las raíces mostró un aumento proporcional con el nivel del NaCl en el substrato, los frutos presentaron una tendencia opuesta. Aunque los síntomas típicos del estrés tales como retraso del crecimiento y la producción reducida de fruta fueron observados en los cultivares de fresa evaluados, Sweet Charlie fue menos sensible a la salinidad.
Palabras clave: Salinidad, estrés osmótico, sólidos solubles totales, acidez titulable, distribución de materia seca, diámetro de frutos.
ABSTRACT
Since water is a very good solvent, all irrigation waters contain some dissolved salts. Salinity restricts the availability of water to plants by lowering the total water potential in the soil. Salinity also has an impact on crop physiology and yield. Visible injury can occur at high salinity levels. This trial was carried out to investigate the relationship between NaCl-salt stress, fruit quality and yield on the strawberries cultivars Sweet Carlie Chandler and Camarosa grown in pots filled with salinized soil in glasshouse. Five salt levels were compared (0, 20, 40, 60 and 80 mM NaCl) which induced different electrical conductivity (EC) levels (2,68; 6,08; 8,24; 9,98 y 12,62 dS.m-1 respectly). The different EC levels were obtained by adding NaCl progressively to the soil. Fruit yield and quality were negatively influenced by the tested treatments. Sugar content in fruits was influenced by the treatments. Total soluble solids and tritrable acidity in fruits grew as EC levels grew. While the accumulation of dry matter in the roots showed a proportional increase with the level of NaCl in the substrate, the fruits showed an opposite tendency. Although typical stress symptoms such as growth retardation and poor fruit yield were observed in the strawberry tested cultivars, Sweet Charlie was less sensitive to the salinity.
Key words: Total soluble solids, tritrable acid, dry matter partitioning, fruit diameter.
 MATERIALES Y MÉTODOS
MATERIALES Y MÉTODOS
RESULTADOS Y DISCUSIÓN
AGRADECIMIENTOS
BIBLIOGRAFÍA
La salinidad en áreas alejadas del mar, en el interior del continente, se hace más frecuente bajo condiciones áridas. Sin embargo, en zonas húmedas se forman parches salinos como consecuencia de depósitos fósiles de sales o del afloramiento de manantiales salinos (Willadino y Camara, 2004). El 10% del área cultivada del planeta presenta suelos afectados por la salinidad (Tanjii 1990) y en las superficies irrigadas, el área salinizada representa del 20 al 27% (Ghassemi et al. 1995). El estrés por salinidad tiene un fuerte impacto sobre la producción comercial de cultivos hortícolas que se desarrollan en condiciones hidropónicas (Sonneveld et al. 1999); sin embargo, los efectos de la salinidad son especialmente agudos en regiones áridas (Kuznetsov y Shevyakova 1997), y la respuesta de los vegetales puede variar considerablemente en función de la tolerancia a la salinidad inherente (Savvas y Lenz 2000). Los efectos nocivos de la salinidad se asocian al déficit de agua, al desbalance iónico, al comportamiento de los estomas, a la eficiencia fotosintética y a la distribución y asimilación del carbono (Greenway y Munns 1980; Bohnert et al. 1995).
El efecto inicial de la salinidad, especialmente a concentraciones de sal moderadas, se debe al efecto osmótico (Munns y Termaat 1986, Jacoby 1994). Las concentraciones específicas de iones pueden modificarse en los tejidos vegetales como consecuencia del estrés por sales, lo que resulta en desbalances por exceso o por deficiencia de iones (Bernstein et al. 1974). Además, la salinidad causa muchos problemas para el crecimiento y desarrollo del vegetal, especialmente para plantas glicófitas, mediante la inducción de mal funcionamiento de procesos fisiológicos (Shannon et al., 1994). La exposición de las plantas al NaCl afecta también el transporte de iones y agua en los vegetales (Lauchli y Epstein, 1990). Estas anormalidades resultan del efecto de la salinidad sobre la disponibilidad de nutrientes, antagonismos y competencia, transporte y distribución en los tejidos del vegetal (Grattan y Grieve, 1999; CasierraPosada et al., 2000; Casierra-Posada et al., 2000a). Por tanto, se han desarrollado muchos ensayos con el objeto de estudiar el efecto de la salinidad sobre los parámetros comúnmente utilizados para medir el crecimiento y la producción en plantas cultivadas (Chartzoulakis y Klapaki, 2000; Perez-Alfosea et al., 1996; Kaya et al., 2002). La salinidad afecta las plantas cultivadas tanto en la fase vegetativa como en la reproductiva, por tanto, causa reducción en la acumulación de biomasa como en la producción (Ebert et al. 1999, Aslam et al. 1993).
Debido a que la mayoría de los representantes del género Fragaria son muy sensible a las condiciones de estrés y a que sus frutos tienen alto valor comercial, se han desarrollado muchas investigaciones para evaluar el comportamiento de las plantas cultivadas bajo diferentes factores causantes de estrés, como presencia de metales pesados (Treder y Cieslinski 2005), sequía (Pomper y Breen 1997, McDonald y Archbold 1998), temperaturas bajas (Nestby y Bjorgum 1999, Rajashekar et al. 1999, Nestby et al. 2001, Lindeen et al. 2002, Houde et al. 2004) y temperaturas altas (Gulen y Eris 2004, Gulen y Eris 2004a, Ledesma y Sugiyama 2005).
Con el incremento del interés por la producción de fresas en sistemas hidropónicos, se ha investigado el efecto de la salinidad por NaCl, con miras a lograr un balance en el crecimiento vegetativo y reproductivo y a mejorar la calidad de fruta (Awang et al. 1993; Awang et al. 1993a). Por otro lado, este cultivo se maneja normalmente bajo condiciones de riego localizado, lo que incrementa la acumulación de sales en la zona de mojado y como consecuencia, se reduce la presión osmótica en la rizósfera, con efectos negativos sobre el crecimiento y la producción de las plantas (Casierra-Posada y García 2005). Como consecuencia de la salinidad en el sustrato se ha reportado un incremento en el contenido de sólidos solubles totales (SST) y de la acidez titulable no sólo en fresa sino también en otros frutales como guayaba (Walker et al. 1979), manzana (Schreiner y Lüdders 1992, ElSiddig y Lüdders 1994), además en tomates (Mizrahi et al. 1988, Adams 1991), pimiento dulce (Janse 1989) y pepino (Chartzoulakis 1992).
El objetivo de el presente estudio fue la evaluación de tres cultivares de fresa cultivados en invernadero bajo condiciones de estrés salino por NaCl, en cuanto a la producción, a la calidad de la fruta, y a la acumulación y distribución de materia seca en raíces, coronas, hojas, flores y frutos.
Para determinar el efecto de la salinidad por NaCl sobre la producción y la calidad de fruta en plantas de fresa, se tomaron los cultivares de fresa Chandler, Camarosa y Sweet Charlie, los cuales se cultivan comúnmente en la zona de estudio. Antes de la siembra, los estolones de fresa se mantuvieron a temperatura de 7 °C durante dos semanas y posteriormente se colocaron en materas plásticas con capacidad para dos kg. de suelo. Una vez las plantas iniciaron la brotación, se aplicó al suelo de cada matera una mezcla de fertilizante comercial de alta solubilidad (Los valores mencionados se adicionaron en g.kg-1de suelo seco): N: 0,4, P2O5: 0,03, K2O: 0,05, CaO: 0,0005, MgO: 0,0013, S: 0,00137, B: 0,0002, Cu: 0,00014, Fe: 0,00012, Mn: 0,0013, Mo: 0,00005 y Zn: 0,0002. Los tratamientos para inducir la salinidad fueron 0, 20, 40, 60 y 80 mM de NaCl. kg-1 de suelo seco al ambiente, los cuales indujeron una conductividad eléctrica de 2,68; 6,08; 8,24; 9,98 y 12,62 dS.m-1, respectivamente, determinados en extracto de saturación. El NaCl se aplicó gradualmente a cada matera, de manera que la cantidad total de la sal se terminara de aplicar en un periodo cercano a cuatro semanas, según la metodología implementada por Alpaslan et al. 1999. A lo largo del ensayo, el suelo de las materas se mantuvo cercano a capacidad de campo. La cantidad de agua a aplicar a las materas se calculó con base en el peso de la matera y en la pérdida de agua con respecto al peso de la misma el día anterior, de manera que las plantas no llegaran al punto de marchitez temporal y que el suelo se mantuviera a capacidad de campo, evitando que hubiera pérdidas de la solución del suelo, por agua gravitacional. Luego de 16 semanas después de terminar los tratamientos con la sal, se cosecharon las plantas y se determinó la acumulación de biomasa en hojas, flores, coronas, frutos y raíces. En frutos se evaluó el peso fresco, la producción total de frutos por planta, el diámetro del fruto, la acidez titulable expresada como porcentaje de ácido cítrico y el contenido de Sólidos Solubles Totales (SST) expresado en °Brix. El arreglo estadístico fue completamente al azar, con cinco reppeticiones por tratamiento. Los resultados obtenidos se analizaron mediante una tabla ANAVA para determinar la significancia. La diferencia entre promedios se determinó mediante la prueba Tukey (P < 0,05). Los análisis estadísticos se realizaron con la versión 11,5 de SPSS (Statistical Package for the Social Sciences - SPSS, Inc., Chicago, Illinois). Las figuras se presentan en formato de barras con su respectiva desviación estándar.
Los tres cultivares evaluados presentaron un comportamiento diferencial en cuanto a la acumulación de peso seco en los diferentes órganos. Es así como Sweet Charlie se vio menos afectado por la salinidad que los demás cultivares respecto a la acumulación de materia seca en los frutos. Por otro lado, la salinidad afectó en menor grado al cultivar Camarosa en cuanto al peso seco de las hojas y raíces que los demás cultivares evaluados (Figura 1 A – E). Respecto a la producción y acumulación de materia seca en hojas, raíces, coronas, flores y frutos en plantas de fresa, la salinidad por NaCl tuvo una fuerte influencia sobre la acumulación de materia seca básicamente en raíces y frutos, en donde hubo diferencias altamente significativas para los factores, concentración de NaCl en el sustrato y cultivar. En las coronas hubo diferencia significativa para el factor variedad. En lo relacionado con la acumulación de peso seco en las flores no se presentó diferencia para ninguno de los factores evaluados (Figura 1 F).
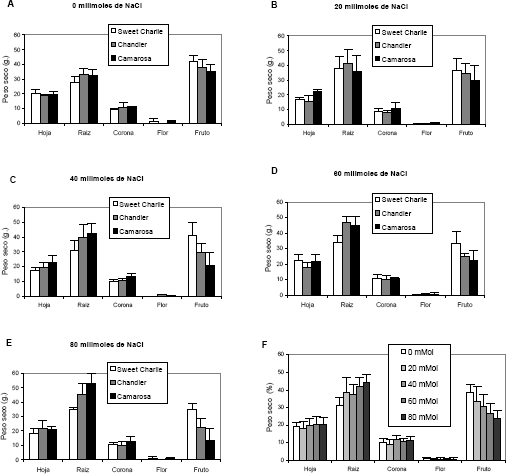
Figura 1. Peso seco en plantas de fresa (Fragaria sp.) cultivadas bajo estrés salino por NaCl. Las figuras A a E, representan el peso seco acumulado en los órganos de los cultivares probados bajo las diferentes concentraciones de NaCl. F, representa la distribución porcentual del peso seco del promedio de los tres cultivares en cada uno de los órganos de la planta.
De igual manera, en plantas de Rubus sp. (Casierra-Posada y Hernández 2006), Agropyron elongatum, Leymus triticoides, Medicago sativum, Paspalum vaginatum, Cynodon dactylum (Suyama et al. 2005), y Fragaria sp. (Casierra-Posada y García 2005) se encontró una reducción de la producción de materia seca bajo condiciones de salinidad. Según otras investigaciones sobre el tema, se ha concluido que la reducción en la fotosíntesis en plantas expuestas a la salinidad depende no sólo de la reducción del CO2 disponible, por el cierre de los estomas, sino también de los efectos acumulativos del contenido de agua y del potencial osmótico en la hoja, de la conductancia estomática, de la tasa de transpiración, del contenido relativo de agua en la hoja, y de componentes bioquímicos tales como pigmentos fotosintéticos, carbohidratos solubles, y proteínas. (Heuvelink, et al. 2003, Sultana et al. 1999, Brugnoli y Lauteri 1991).
Kepenek y Koyuncu 2002, encontraron que las plantas de fresa expuestas a la salinidad responden con una reducción en el crecimiento vegetativo expresado en términos de peso fresco y seco de la planta, número de estolones, área foliar, altura de planta, longitud de la raíz principal y número de hojas; sin embargo, la respuesta de las plantas de fresa al efecto de la salinidad es una consecuencia de la posibilidad de tolerancia de los diferentes cultivares evaluados. Por su parte, Kaya et al. 2002, encontraron también, un componente varietal relacionado con la producción de materia seca en cultivares de fresa sometidos a estrés por NaCl; es así, como la reducción en la acumulación total de fitomasa fue mayor en el cultivar Camarosa que en Oso Grande.
Los resultados encontrados respecto a la disminución diferencial del peso seco en las variedades evaluadas se puede relacionar con diferencias en la asimilación neta de los materiales genéticos, dado que según la información obtenida por Saied et al. 2003, el cultivar de fresa Elsanta mostró menor disminución en la eficiencia de la carboxilación y en la tasa máxima de asimilación que el cultivar Korona expuestos a salinidad por NaCl. Como justificación, afirman estos autores, que la tasa máxima de asimilación es un indicador de la limitación de la fotosíntesis neta bajo saturación de luz y concentración adecuada de CO2, la eficiencia de la carboxilación refleja la actividad de la Ribulosa-1,5 bifosfato carboxilasa/oxigenasa (Rubisco), una enzima esencial en el ciclo de Calvin durante las reacciones oscuras de la fotosíntesis, lo que indica que la fotosíntesis se ve más afectada en los cultivares de fresa que producen menor cantidad de materia seca en sus diferentes órganos; además, en los cultivares de fresa expuestos a condiciones de salinidad por NaCl las reacciones oscuras de la fotosíntesis, son relativamente menos afectadas por la salinidad que las reacciones lumínicas.
Por su parte, Viswanathan y Zhu (2003) discuten que el potencial del plasmalema de las células vegetales favorece el transporte pasivo del Na+ hacia el interior de las células, especialmente bajo condiciones de alto contenido de Na+ extracelular; sin embargo, el exceso de Na+ extracelular entra a la célula a través del transportador HKT1 y de los canales transportadores de cationes no selectivos, lo cual resulta en una disminución de la relación K+/Na+ en el citosol, que tiene como consecuencia la alteración de los procesos metabólicos en los cuales está implicado el K+ y por tanto, el crecimiento, expresado en términos de acumulación de materia seca, se ve seriamente afectado.
Cuando plantas glicofitas se exponen a la salinidad se reduce el crecimiento, asociado con disminución en los valores de algunos de los factores determinantes del proceso fotosintético. El efecto del estrés salino sobre la reducción de la anhidrasa carbónica (CA) puede ser uno de los factores que limitan fotosíntesis (Syeed y Khan, 2004). Es de resaltar que la CA cataliza la conversión reversible del HCO3- y del CO2 y proporciona el CO2 a la enzima Rubisco para la carboxilación.
Se ha encontrado que en condiciones salinas, la disminución de la fotosíntesis puede resultar también debido a la inhibición en la biosíntesis de clorofilas, en relación con el aumento de etileno que afecta la actividad de la enzima ácido-1-amino ciclopropano-1-carboxíloco oxidasa (ACC-oxidasa) (Cramer 1992, Saha y Gupta 1999). De igual manera, el contenido de clorofilas, bajo condiciones de estrés salino, se puede afectar por deficiencia de magnesio (Ali-Dinar et al. 1999), resultante del exceso en la toma de Na+, cuya concentración incrementa la relación Na/Mg en las hojas (Casierra-Posada et al. 2000; Casierra-Posada et al. 2000a).
La producción total de fruta se redujo con el incremento de la concentración de NaCl en el sustrato (Figura 2). Para esta variable hubo diferencias altamente significativas para los factores concentración de NaCl en el sustrato y para cultivar. De hecho, el incremento en la conductividad eléctrica como consecuencia de la adición de NaCl al sustrato conducen a un desbalance metabólico que se refleja en la reducción de la fruta cosechada a partir de plantas cultivadas bajo estrés salino, comparadas con los testigos. El metabolismo anormal causado por la salinidad en plantas de fresa es una consecuencia de las alteraciones en la toma de nutrientes y agua, debidas al potencial osmótico bajo en el suelo. Por su parte, bajo condiciones de salinidad por NaCl, el Cl- y el Na+ son los iones dominantes en la solución del suelo. A pesar de que ambos son esenciales para el buen desarrollo los vegetales, su concentración en suelos salinos excede la demanda, por tanto, inducen toxicidad en plantas sensibles a la salinidad (Marschner 1995).
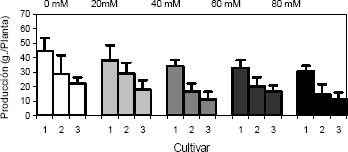
Figura 2. Producción total de fruta en plantas de fresa (Fragaria sp.) cultivadas bajo estrés salino por NaCl. Cultivar 1: Sweet Carlie, cultivar 2: Chandler, cultivar 3: Camarosa.
A pesar de que la fresa es una planta sensible a la salinidad (Maas 1992), en este trabajo se encontraron diferencias altamente significativas en cuanto a la producción de fruta en los tres cultivares probados, de los cuales el cultivar Sweet Charlie se vio menos afectado en cuanto a la producción de fruta que los cultivares Camarosa y Chandler. Al respecto, Kepenek y Koyuncu 2002 y 2002a, evaluaron la tolerancia a la salinidad de diferentes cultivares nativos y foráneos de fresa, de los cuales, los nativos se vieron menos afectados por la salinidad inducida mediante la adición de NaCl al sustrato. Dado que los cultivares de fresa Camarosa y Oso Grande evaluados por Kaya et al. 2002, presentaron diferencias en cuanto a la reducción en la producción de fruta, estos autores le atribuyen la reducción en la producción de fresa a diferencias en la permeabilidad en la membrana celular en los cultivares, puesto que la presencia de NaCl en el sustrato induce una mayor permeabilidad en el plasmalema. Por otro lado, encontraron que tanto el nitrógeno como el calcio se redujeron en los tejidos vegetales como consecuencia de la salinidad, lo cual, para el caso del nitrógeno, es un efecto del antagonismo ente el cloro y la toma de nitratos en el sistema radical (Marschner 1995), lo que induce una disminución en el contenido de clorofila (Kaya et al. 2002), en la tasa de asimilación de las plantas de fresa y en la eficiencia de la carboxilación, tomados como indicadores de la fotosíntesis neta bajo condiciones de salinidad en el sustrato, saturación de luz y concentración de CO2 no limitante (Saied et al. 2003). Como consecuencia, se presenta un descenso en la producción de fruta de las plantas de fresa bajo condiciones de salinidad por NaCl.
Se encontraron diferencias significativas para la variable peso fresco por fruto, de acuerdo con los factores concentración de NaCl en el sustrato y cultivar. Al igual que para la variable comentada anteriormente, el peso seco tuvo un comportamiento inversamente proporcional a la concentración de sal. Con respecto al cultivar, Sweet Carlie mostró un mayor peso de frutos en comparación con los demás cultivares evaluados (Figura 3). Este resultado coincide con lo hallado por DAnna et al. 2003, quienes encontraron que el peso máximo por fruto en las plantas de fresa se obtuvo cuando éstas se sometieron a una conductividad eléctrica de 2,5 mS.cm-1. Valores superiores en la conductividad eléctrica redujeron el peso por fruto cosechado.
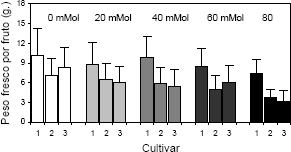
Figura 3. Peso fresco por fruto en plantas de fresa (Fragaria sp.) cultivadas bajo estrés salino por NaCl. Cultivar 1: Sweet Carlie, cultivar 2: Chandler, cultivar 3: Camarosa.
El diámetro longitudinal y ecuatorial de los frutos de las plantas de fresa sometidas a salinidad por NaCl, de igual manera se redujo (Figura 4). Las plantas tratadas con concentraciones elevadas de la sal, produjeron frutos de menor diámetro. Al respecto se encontraron diferencias altamente significativas para las variables diámetro longitudinal y diámetro transversal de frutos con respecto a los factores concentración de NaCl en el sustrato y cultivar.
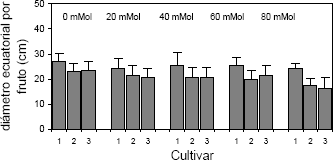
Figura 4. Diámetro ecuatorial por fruto en plantas de fresa (Fragaria sp.) cultivadas bajo estrés salino por NaCl. Cultivar 1: Sweet Carlie, cultivar 2: Chandler, cultivar 3: Camarosa.
La cantidad de sólidos solubles totales en frutos de las plantas de fresa cultivadas bajo condiciones de salinidad por NaCl, se incrementó proporcionalmente con la concentración de la sal en el sustrato (Figura 5). Se presentaron diferencias significativas para los factores concentración de NaCl en el sustrato y cultivar.
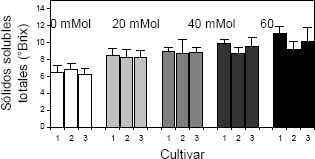
Figura 5. Concentración de sólidos solubles totales (SST) en frutos de fresa (Fragaria sp.) cultivadas bajo estrés salino por NaCl. Cultivar 1: Sweet Carlie, cultivar 2: Chandler, cultivar 3: Camarosa.
En cuanto a este parámetro, Keutgen y Keutgen 2003, encontraron que, mientras en el cultivar de fresa Elsanta, la salinidad indujo una reducción en el contenido de sólidos solubles totales, en el cultivar Korona el contenido de éstos se incrementó; sin embargo, en comparación con el contenido de ácidos orgánicos, la reducción en los azucares reductores en la fruta fresca, fue más marcada, produciendo como resultado la tendencia, comúnmente observada en cuanto al incremento en la calidad de la fruta producida cuando se exponen las especies frutícolas a condiciones de salinidad.
El estrés osmótico induce un incremento en la toma de cationes (K+ y Na+) y su posterior secuestro en las vacuolas, así como la síntesis de osmoprotectantes, que son solutos orgánicos compatibles con la planta, como la prolina, la betaina, polioles y azucares solubles (Larcher 2001, Willadino y Camara 2004), lo que puede explicar el incremento en el contenido de sólidos solubles en los frutos. Estos solutos compatibles, actúan tanto en el ajuste osmótico como en la osmoprotección. En el ajuste cumplen el papel de osmolitos, facilitando la retención de agua en el citosol y posibilitando el secuestro del Na+ en la vacuola o en el apoplasto. Por otro lado, pueden tener también la función de protección de las estructuras celulares a través de interacciones con membranas, enzimas o complejos proteicos (Bohnert et al. 1995, Kreeb 1996). Sin embargo, a pesar de los beneficios de la producción de osmoprotectantes, la síntesis de solutos compatibles implica, por otro lado, un desvío del carbono que, en un principio, se utilizaría en los procesos biosintéticos del metabolismo primario, de los cuales resulta el crecimiento de la planta (Raven 1985), lo que implica una disminución en la acumulación de materia seca.
De igual manera que para los sólidos solubles totales, la cantidad de ácidos orgánicos contenidos en frutos de las plantas de fresa cultivadas bajo condiciones de salinidad por NaCl, se incrementó, proporcionalmente, con la concentración de la sal en el sustrato (Figura 6). Con diferencias significativas para los factores, concentración de NaCl en el sustrato y cultivar. Por el contrario, Kepenek y Koyuncu 2002a, manifiestan que la salinidad induce una reducción, tanto en la acidez titulable como en la cantidad de sólidos solubles totales en los frutos. Además, estos autores expresan que existe un fuerte componente varietal que determina la respuesta de las plantas de fresa a la salinidad, en lo relacionado con la calidad de fruta; por tanto, el comportamiento de las plantas de fresa, expuestas a niveles crecientes de salinidad en el sustrato, depende de los cultivares en estudio y de la capacidad inherente a cada cultivar para tolerar, hasta cierto límite, el efecto negativo de la salinidad.
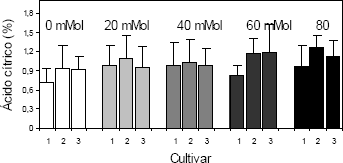
Figura 6. Acidez titulable en frutos de fresa (Fragaria sp.) cultivadas bajo estrés salino por NaCl. Cultivar 1: ‘Sweet Carlie’, cultivar 2: ‘Chandler’, cultivar 3: ‘Camarosa’.
Mientras algunos cultivares, como los evaluados por Kepenek y Koyuncu 2002a, reducen el contenido de ácidos orgánicos en los frutos, cuando se exponen a una conductividad eléctrica elevada como consecuencia de la concentración de sales en el sustrato, otros cultivares como Sweet Carlie, Chandler y Camarosa, estudiados en el presente trabajo incrementan tanto la acidez titulable como los sólidos solubles totales en los frutos, dado que las posibilidades de tolerancia de los diferentes materiales vegetales de fresa es distinta.
Con base en los resultados y discusión expuestos, se puede concluir que las plantas de fresa son muy sensibles a la salinidad por NaCl, a pesar de lo cual, su respuesta, medida en términos de crecimiento del vegetal, es altamente dependiente del material genético en cuestión, pues hay diferencias muy notorias entre cultivares en cuanto a su tolerancia a los efectos negativos de la salinidad.
La salinidad indujo modificaciones en el patrón de acumulación de materia seca en los diferentes órganos de las plantas. De este modo, la fitomasa se acumuló en las raíces, de manera directamente proporcional a la salinidad, mientras que en los frutos, sucedió lo contrario.
La calidad de fruta expresada como peso fresco y diámetro de frutos se vio fuertemente disminuida por el incremento en la conductividad eléctrica, resultante de la salinidad; mientras que, por el contrario, los sólidos solubles totales y la acidez titulable tuvieron un incremento directamente proporcional a la salinidad.
Este estudio fue desarrollado con recursos del Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología Francisco José de Caldas COLCIENCIAS y de la Dirección de Investigaciones (DIN) de la Universidad Pedagógica y Tecnológica de Colombia, en el marco del proyecto DIN-No 9001.059 Evaluación de la tolerancia de plantas de fresa (Fragaria sp.) al estrés osmótico.
BIBLIOGRAFÍA
Adams, P. 1991. Effect of increasing the salinity of nutrient solution with major nutrients or sodium chloride on yield, quality and composition of tomatoes grown in rockwool. En: Journal of Horticultural Science. Vol. 66, no. 6; p. 201-207. [ Links ]
Ali-Dinar, H.M.; Ebert, G. and Lüdders, P. 1999. Growth, chlorophyll content, photosynthesis and water relations in guava (Psidium guajava L.) under salinity and different nitrogen supply. En: Garten-bauwissenschaft. Vol 64, no. 2; p. 54-59. [ Links ]
Alpaslan, M., Inal, A., Günes, A., Çikili, Y. and Özcan, H. 1999. Effect of treatment on alleviation of sodium and chloride injury in tomato (Lycopersicum esculen-tum (L.) Mill. Cv. Lale) grown under salinity. En: Turkish Journal of Botany. Vol. 23; p. 1-6. [ Links ]
Aslam, M.; Qureshi, R. H. and Ahmad, N. 1993. A rapid screening technique for salt tolerance in rice (Oriza sativa L.). En: Plant and Soil. Vol. 150; p. 99-107. [ Links ]
Awang, Y. B.; Atherton, J. G. and Taylor, A. J. 1993. Salinity effects on strawberry plants grown in rockwool. I. Growth and leaf relations. Journal of Horticultural Science, 68: 783-790. [ Links ]
Awang, Y. B.; Atherton, J. G. and Taylor, A. J. 1993a. Salinity effects on strawberry plants grown in rockwool. Part II. Fruit quality. En: Journal of Horticultural Science. Vol. 68; p. 791-795. [ Links ]
Bernstein, L.; Francois, L. E. and Clark, R. A. 1974. Interactive effects of salinity and fertility on yield of grains and vegetables. En: Agronomy Journal. Vol. 66; p. 661-667. [ Links ]
Bohnert , H. J.; Nelson, D.E. and Jensen, R.G. 1995. Adaptations to environmental stress. En: Plant and Cell. Vol. 7, no.7; p. 1109-1111. [ Links ]
Brugnoli, E. and Lauteri, M. 1991. Effects of salinity on stomatal conductance, photosynthetic capacity, and carbon isotope discrimination of salt- tolerant (Gossypium hirsutum L.) and salt-sensitive (Phaseolus vulgaris L.) C3 non-halophytes. En: Plant Physiology. Vol. 95; p. 628-635. [ Links ]
Casierra–Posada, F.; Ebert, G. y Lüdders, P. 2000. Efecto de la salinidad por cloruro de sodio sobre el balance de nutrientes en plantas de lulo. En: Agronomía Colombiana. Vol. 17, no. 1-3; p. 85-90. [ Links ]
________,Ebert, G. y Lüdders, P. 2000a. Acumulación y distribución de iones en tejidos de lulo (Solanum quitoense) en condiciones de salinidad por cloruro de sodio. p. 76-80. III Seminario de Frutales de Clima Frío Moderado. (3°: 2000: Manizales, Colombia). Memorias. Manizales: Centro de Desarrollo Tecnológico de Frutales. [ Links ]
________ y García, N. 2005. Crecimiento y distribución de materia seca en cultivares de fresa (Fragaria sp.) bajo estrés salino. En: Agronomía Colombiana. Vol. 23, no.1; p. 83-90. [ Links ]
________ y Hernández, H. L. 2006. Evapotranspiración y distribución de materia seca en plantas de mora (Rubus sp.) bajo estrés salino. En: Revista Actualidad y Divulgación Científica U.D.C.A. Vol. 9, no.1; p. 85-95. [ Links ]
Chartzoulakis, K. 1992. Effects of NaCl salinity on germination, growth and yield of greenhouse cucumber. En: Journal of Horticultural Science. Vol. 67, no. 3; p. 115-119. [ Links ]
Chartzoulakis, K. and Klapaki, G. 2000. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. En: Scientia Horticulturae. Vol. 86; no. 4; p. 247-260. [ Links ]
Cramer, G. R. 1992. Kinetics of maize leaf elongation. III Silver thiosulphate increases the yield of salt stressed plants, but ethylene is not involved. En: Plant Physiology. Vol. 100, no. 2; p. 1044-1047. [ Links ]
DAnna, F.; Incalcaterra. G.; Moncada, A. and Miceli, A. 2003. Effects of different electrical conductivity on strawberry grown in soilless culture. En: Acta Horticulturae. Vol. 609; p. 355-360. [ Links ]
Ebert, G.; Casierra, F. and Lüdders, P. 1999. Influence of NaCl salinity on growth and mineral uptake of lulo (Solanum quitoense). En: Angewandte Botanik. Vol. 73; p. 31-33. [ Links ]
El-Siddig, K. and Lüdders, P. 1994. Interactive effects of nitrogen nutrition and salinity on reproductive growth of apples trees. En: Gartenbauwissen-schaft. Vol. 59, no. 2; p. 58-61. [ Links ]
Ghassemi, F., Jakeman, A. J. and Nik, H. A. 1995. Salinization of land and water resources. Human causes, extent, management and case studies. Australia: New South Wales Press, 526 p. [ Links ]
Grattan, S. R. and Grieve, C. M. 1999. Salinity mineral nutrient relations in horticultural crops. En: Scientia Horticulturae. Vol. 78, no. 1-4; p. 127-157. [ Links ]
Greenway, H, and Munns, R. 1980. Mechanisms of salt tolerance in non halophytes. Annual Review in Plant Physiology. Vol. 31; p. 149-190. [ Links ]
Gulen, H. and Eris A. 2004. Some physiological changes in strawberry (Fragaria x Ananassa 'Camarosa') plants under heat stress. En: Journal of Horticultural Science and Biotechnology. Vol. 78, no. 6; p. 894-898 [ Links ]
Gulen, H. and Eris, A. 2004a. Effect of heat stress on peroxidase activity and total protein content in strawberry plants. En: Plant Science. Vol. 166, no. 3; p. 739-744. [ Links ]
Heuvelink, E. P., Bakker, M. and Stanghellini, C. 2003. Salinity effects on fruit yield in vegetable crops: a simulation study. En: Acta Horticulturae. Vol. 609; p. 133-140. [ Links ]
Houde, M., Dallaire, S., N'Dong, D. and Sarhan, F. 2004. Overexpression of the acidic dehydrin WCOR410 improves freezing tolerance in transgenic strawberry leaves. En: Plant Biotechnology Journal. Vol. 2, no. 5; p. 381-387 [ Links ]
Jacoby, B. 1994. Mechanisms involved in salt tolerance by plants. p. 97-123. En: Pessarali, M., ed. Handbook of plant and crop stress. Nueva York Marcel Dekker. [ Links ]
Janse, J. 1989. Effects of humidity, temperature and concentration of the nutrient solution on firmness, shelflife and flavor of sweet pepper fruits (Capsicum annuum L.). En: Acta Horticulturae. Vol. 244; p. 123-132. [ Links ]
Kaya, C., Erol, B., Higgs, D. and Murillo-Amador, B. 2002. Influence of foliar-applied calcium nitrate on strawberry plants grown under salt-stressed conditions. Australian Journal of Experimental Agriculture. Vol. 42, no. 5; p. 631-636. [ Links ]
Kepenek, K. and Koyuncu, F. 2002. Effect of salt expression of resistance in some domestic foreign strawberry cultivars. Acta Horticulturae. Vol. 573; p. 289-295. [ Links ]
Kepenek, K. and Koyuncu, F. 2002a. Studies on the salt tolerance of some strawberry cultivars under glasshouse. En: Acta Horticulturae. Vol. 573; p. 297-304. [ Links ]
Keutgen, A. J. and Keutgen, N. 2003. Influence of NaCl salinty on stress fruit quality in strawberry. En: Acta Horticulturae. Vol. 609; p. 155-157. [ Links ]
Kreeb, K. H. 1996. Salzstress. p. 149-172. En: Brunold, CH. , Rüegesegger, A. und Brändle, R., eds. Stress bei Pflanzen. Stuttgart (Alemania): UTB für Wissenschaft, Paul Haupt. [ Links ]
Kuznetsov, V. V. and Shevyakova, N. I. 1997. Stress responses of tobacco cells to high temperature and salinity. Proline accumulation and phosphorilation of polypeptides. En: Physiologia Plantarum. 100, no. 2; p. 320-326. [ Links ]
Larcher, W. 2001. Ökophysiologie der Pflanzen. Stuttgart (Alemania): Eugen Ulmer. p. 341-349. [ Links ]
Lauchli, A. and Epstein, E. 1990. Plant responses to saline and sodic conditions. p. 113-137. En: Tanji, P. K. K., ed. Agricultural salinity assessment and management. (Manuals Rep. On. Eng. Practice No 71). [ Links ]
Ledesma, N. and Sugiyama, N. 2005. Pollen quality and performance in strawberry plants exposed to high-temperature stress. En: Journal of the American Society for Horticultural Science. Vol. 130, no. 3; p. 341-347. [ Links ]
Lindeen, L., Palonen, P., and Hytonen T. 2002. Evaluation of three methods to assess winter hardiness of strawberry genotypes. En: Journal of Horticultural Science and Biotechnology. Vol. 77, no. 5; p. 580-588. [ Links ]
Maas, J. L. 1992. Compendium of strawberry diseases. St. Paul, Minnesota, USA: American Phytopathological Society. APS Press. 138 p. [ Links ]
Marschner, H. 1995. Mineral nutrition of higher plants. Londres: Academic Press. p. 657-680. [ Links ]
McDonald, S. S. and Archbold,
Mizrahi, Y., Taleisnik, E., Kagan-Zur, V., Zohar, Y., Offenbach, R., Matan, E. and Golan, R. 1988. A saline irrigation regime for improving tomato fruit quality without reducing yield. En: Journal of the American Society for Horticultural Science, 113, no. 2; p. 202-205. [ Links ]
Munns, R. and Termaat, A. 1986. Whole plant response to salinity. En: Australian Journal of Plant Physiology. Vol. 13, no. 1; p. 143-160. [ Links ]
Nestby, R. and Bjorgum, R. 1999. Freeze injury to strawberry plants as evaluated by crown tissue browning, regrowth and yield parameters. En: Scientia Horticulturae. Vol. 81, no. 3; p. 321-329. [ Links ]
Nestby, R., Bjorgum, R., Nes, A., Wikdahl, T. and Hageberg, B. 2001. Reactions of strawberry plants to longterm freezing and alternate freezing and thawing. En: Journal of Horticultural Science and Biotechnology. Vol. 76, no. 3; p. 280-285. [ Links ]
Perez-Alfocea, F., Balibrea, M. E., Santa Cruz, A. and Estan, M. T. 1996. Agronomical and physiological characterization of salinity tolerance in a commercial tomato hybrid. En: Plant and Soil. Vol. 180, no. 2; p. 251-257. [ Links ]
Pomper K. W. and Breen P. J. 1997. Expansion and osmotic adjustment of strawberry fruit during water stress. En: Journal of American Society for Horticultural Science. Vol. 122, no. 2; p. 183-189. [ Links ]
Rajashekar, C. B., Zhou, H., Marcum, K. B. and Prakash, O. 1999. Glycine betaine accumulation and induction of cold tolerance in strawberry (Fragaria x ananassa Duch.) plants. En: Plant Science. Vol. 148, no. 2; p. 175-183. [ Links ]
Raven, J, A. 1985. Regulation of pH and generation of osmolarity in vascular plants: a costbenefit analysis in relation to efficiency of use of energy, nitrogen and water. En: New Phytologist. Vol. 101, no. 1; p. 25-77. [ Links ]
Saha, K. and Gupta, K. 1999. Effect of NaCl salinity on ethylene production and metabolism in sunflower seedlings. En: Plant Physiology and Biochemistry Vol. 2; p. 127-130. [ Links ]
Saied, A. S., Keutgen, N. and Noga, G. 2003. Effects of NaCl on leaf growth, photosynthesis and ionic contents of strawberry cvs Elsanta and Korona. En: Acta Horticulturae. Vol. 609; p. 67-73. [ Links ]
Savvas, D. and Lenz, F. 2000. Effects of NaCl or nutrient induced salinity on growth, yield and composition of eggplants grown in rock wool. En: Scientia Horticulturae. Vol. 84, no. 1-2; p. 37-47. [ Links ]
Schreiner, M. und Lüdders, P. 1992. Einfluss von Na-Salzen auf die Na[+]-, K[+]-, Cl[−]- und SO[4][2−]-Gehalte in Blättern von Golden Delicious'- Apfelbäumen bei unterschiedlicher K-Ernährung. En: Angewandte Botanik. Vol. 66, no.1-2; p. 1-6. [ Links ]
Shannon, M. C., Grieve, C. M. and Francois, L. E. 1994. Whole-plant to salinity. p. 199-244. En: Wilkinson, R. E., ed. Plant-environment interactions. Nueva York: Marcel Dekker. [ Links ]
Sonneveld, C., Baas, R., Nussen, H. M. C. and De Hoog, J. 1999. Salt tolerance of flower crops in soilless culture. En: Journal of Plant Nutrition. Vol. 22, no. 6; p. 1033-1048. [ Links ]
Sultana, N., Ikeda, T. and Itoh, R. 1999. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. En: Environmental and Experimental Botany. Vol. 42, no. 3; p. 211-220. [ Links ]
Suyama, H., Benes, S., Robinson, P., Getachew, G., Grattan, S. R. and Grieve, C. M. 2005. Biomass production and nutritional value of forages irrigated with saline-sodic drainage water in a green-house study. p. 175-178. En: Proceedings of the International Salinity Forum, Managing Saline Soils and Water. Riverside, CA: Science, Technology, and Soil Issues. [ Links ]
Syeed, S. and Khan, N. A. 2004. Activities of carbonic anhydrase, catalase and ACC oxidades of mung bean (Vigna radiata) are differentially affected by salinity stress. En: Food, Agriculture & Environment. Vol. 2, no. 2; p. 241-249. [ Links ]
Tanjii, K. K. 1990. Nature and extent of agricultural salinity. p. 1-17. En: Tanjii, K. K., ed. Agricultural salinity assessment and management, Nueva York: American Society of Civil Engineers. [ Links ]
Treder, W. and Cieslinski, G. 2005. Effect of silicon application on cadmium uptake and distribution in strawberry plants grown on contaminated soils. En: Journal of Plant Nutrition. Vol. 28, no. 6; p. 917-929. [ Links ]
Maas, J. L.. 1992. Compendium of strawberry diseases. St. Paul, Minnesota-USA: American Phytopathological Society. APS Press. 138 p. [ Links ]
Viswanathan, C. and Zhu, J. K. 2003. Plant salt tolerance. p. 241-270. En: Hirt, H. y Schinozaki, K., ed. Plant responses to abiotic stress. Topics in current genetics, v. 4. Berlin: Springer-Verlag. [ Links ]
Walker, R. R., Kriedmann, P. E. and Maggs, D. H. 1979. Growth, leaf physiology and fruit development in salt-stressed guavas. En: Australian Journal of Agricultural Research. Vol. 30, no. 3; p. 477-488. [ Links ]
Willadino, L. and Camara, T. 2004. Origen y naturaleza de los ambientes salinos. p. 303-330. En: Reigosa, M. J.; Pedrol, N. Y Sánchez, A., eds. La ecofisiología vegetal. Una ciencia de síntesis. Madrid: Thompson. [ Links ]

