










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.60 no.2 Medellín July/Dec. 2007
VARIABILIDAD MORFOLOGICA DE LA COLECCIÓN COLOMBIANA DE LULO (Solanum quitoense Lam.) Y ESPECIES ELACIONADAS DE LA SECCIÓN Lasiocarpa
MORPHOLOGICAL VARIABILITY OF THE COLOMBIAN COLLECTION OF LULO (Solanum quitoense Lam.) AND RELATED LASIOCARPA SECTION SPECIES
Mario Lobo Arias1; Clara Inés Medina Cano2; Oscar Arturo Delgado Paz3 y Armando Bermeo Giraldo4
1 Investigador Titular. CORPOICA, Centro de Investigación La Selva. Km 7 Vía las Palmas. Rionegro, Colombia y Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779. Medellín, Colombia. <mlobo@corpoica.org.co>
2 Investigador Asistente. CORPOICA, Centro de Investigación La Selva. Km 7 Vía las Palmas. Rionegro, Colombia. <cmedina@corpoica.org.co>
3 Investigador Asistente. CORPOICA, Centro de Investigación La Selva. Km 7 Vía las Palmas. Rionegro, Colombia. <odelgado@corpoica.org.co>
4 Ingeniero Agrónomo. Universidad Nacional de Colombia, Sede Medellín. A.A. 1779, Medellín, Colombia. <abermeo@unalmed.edu.co>
Recibido: Septiembre 4 de 2007; aceptado: Noviembre 10 de 2007
RESUMEN
Se categorizó la variabilidad morfológica de la colección colombiana de lulo Solanum quitoense Lam, especies relacionadas de la sección Lasiocarpa y entidades biológicas Solanaceae de otras secciones, como grupo externo. Se encontró amplia variabilidad morfológica cualitativa y cuantitativa en las dos variedades botánicas del lulo y las otras entidades biológicas de Lasiocarpa. Los 58 atributos morfológicos cualitativos fueron polimórficos en éste conjunto, con presencia de 73,9% del total de estados incluidos en los descriptores y 4,2 morfoalelos por variable. En el caso de S. quitoense, 52 de los 58 caracteres cualitativos exhibieron variabilidad, con 60% de las variantes totales de la lista de descriptores y 3,4 morfoalelos por característica. Igualmente, se encontró polimorfismo considerable en S. hirtum, S. pseudolulo y potencial de utilización, en dos materiales estudiados de S. vestissimum. A nivel cuantitativo se determinó diversidad en todas las variables estudiadas. El análisis de componentes principales de los caracteres cuantitativos reveló una alta contribución de las variables de fruto a la explicación de la variabilidad total de esta índole. Los fenogramas cualitativo, cuantitativo y cualitativo-cuantitativo, no detectaron materiales idénticos en los taxa estudiados. La mayor consistencia taxonómica fue exhibida por el fenograma obtenido con todas las variables cualitativas y cuantitativas.
Palabras claves: Naranjilla, recursos genéticos, caracterización, evaluación, agrupamientos.
ABSTRACT
The morphologic variability of the Colombian collection of lulo, Solanum quitoense Lam, related species of section Lasiocarpa and Solanaceae taxa of other sections, as outgroup, was studied. Ample qualitative and quantitative morphologic variability was found in the two botanical varieties of lulo and the other studied Lasiocarpa species. The 58 qualitative morphologic attributes were polymorphic in this set of taxa, with 73,9% of the total qualitative states included in the descriptor list up today, with 4,2 morphoalleles by variable. In the case of S. quitoense, 52 of the 58 qualitative characters exhibited variability, with 60% of the total variants of the list and 3,4 morphoalleles by characteristic. Also, considerable polymorphism was found in S. hirtum, S. pseudolulo, with potentiality of use of the two studied accessions of S. vestissimum. At quantitative level diversity in all the studied variables was determined. The analysis of main components of the quantitative characters revealed a high contribution of the fruit variables to the total variability of this nature. Qualitative, quantitative and qualitative-quantitative phenograms did not detect identical materials in the studied taxa. The largest taxonomic consistency was exhibited by the phenogram obtained with all the qualitative and quantitative variables.
Key words: Genetic resources, characterization, evaluation, clustering pattern
 MATERIALES Y MÉTODOS
MATERIALES Y MÉTODOS
RESULTADOS Y DISCUSIÓN
CONCLUSIONES
AGRADECIMIENTOS
BIBLIOGRAFÍA
El lulo o naranjilla (Solanum quitoense Lam.), Solanaceae, es un frutal andino considerado como promisorio desde hace más de 80 años (Popenoe, 1924). Este fue incluido en un conjunto de plantas subutilizadas con potencial económico, en la década de los 70 (National Academy of Sciences, 1975). Actualmente, la especie ha adquirido desarrollo en Colombia y el Ecuador, con un área de siembra estimada para el año 2006, en Colombia, del orden de 5467 hectáreas y una oferta de fruta de 41064 toneladas (Arias, Támara y Arbeláez, 2006). En el país hay una demanda insatisfecha, por lo cual se realizan importaciones a partir del Ecuador, las que se consideraron, en el año 2000, que correspondían al 21,7% del consumo nacional (Torres, 2002).
Las posibilidades de un mayor cre-cimiento de los sistemas productivos con el taxón se derivan de una serie de aspectos puntualizados por Lobo (2000, 2004), los cuales incluyen: existencia en la zona andina de variabilidad a nivel de la especie cultivada y taxa relacionados; nichos apropiados para su siembra; aceptación de las bayas; déficit de con-sumo de frutas frescas en Latinoamérica; potencial agroindustrial; factibilidad de producir desarrollo económico a nivel de los productores; bienestar social como fuente de empleo tanto a nivel de los cultivos como en su industrialización; alternativa para el reemplazo de cultivos ilícitos y competencia prácticamente nula con otras zonas del mundo para su producción.
La planta pertenece a la sección Lasiocarpa, clase que incluye entre 11 y 13 especies (Whalen, Costich y Heiser, 1981; Heiser, 1996; Bohs, 2004), distribuidas principalmente en el noroeste de Suramérica, con presencia de un taxón, Solanum ferox, en el continente Asiático (Whalen, Costich y Heiser, 1981; Heiser y Anderson, 1999) y de algunos de los taxa en el escudo de las Guyanas y el norte de Brasil (Bohs, 2004). Del conjunto de entidades biológicas anteriores, el lulo Solanum quitoense y la cocona, cubiu o lulo amazónico, Solanum sessiliflorum son cultivos importantes en América Latina (Heiser, 1969, 1985). De éstos, el primer frutal, se introdujo y es cultivado en Venezuela, Panamá, Guatemala y Costa Rica (Heiser y Anderson, 1999; Lobo y Medina, 2000; Bohs, 2004). El lulo fue encontrado y descrito por los conquistadores españoles en Ecuador y Colombia (Patiño, 1962); sin ubicación de registros arqueológicos para esta especie (Heiser, 1985).
El centro primario de diversidad genética del taxón comprende Colombia, Ecuador y Perú, encontrándose entre los 1200 y 2300 msnm (Heiser y Anderson, 1999; Lobo y Medina, 2000). Heiser (1993) indicó que las plantas cultivadas actuales no deben diferir mucho de las halladas por los conquistadores al llegar a América por el poco trabajo de selección y mejoramiento que se ha practicado con esta entidad biológica; a lo cual adicionaron Heiser (1972); Whalen, Costich y Heiser (1981), que a pesar del grado de domesticación de la planta, ésta exhibe una variabilidad escasa.
Lobo (2004), con base en argumentos genéticos y lingüísticos, considera que el centro nuclear de la especie es Colombia; lo cual se basa en la mayor presencia de algunos atributos ancestrales en la poblaciones colombianas, como son las espinas y que en Colombia, a diferencia del Ecuador, donde la planta es conocida con la palabra española naranjilla, se le denomina con el vocablo lulo de origen quechua (Patiño, 1962, 2002) y con otros nombres ancestrales, como son: machakve, en el dialecto Kamsá (Schultes, 1949) y Monai en el lenguaje Tunebo (Lobo, 2004).
El lulo no corresponde a una planta completamente domesticada (Lobo, 1991). Lo anterior es soportado por una serie de atributos existentes en éste, los cuales corresponden al complejo silvestres-malezas (arvenses), como son: alogamia; adaptación estrecha en las poblaciones espontáneas y locales; espinas en los tallos, las ramas y las hojas; antocianinas en diversos órganos; frutos recubiertos por tricomas; oxidación rápida de los jugos; elevado número de semillas por baya (Lobo 2000); latencia en éstas (Cárdenas, Zuluaga y Lobo, 2004); andromonoecia no plástica en la especie cultivada (Miller y Diggle, 2003) y hojas con ideoblastos que contienen cristales de oxalato de calcio (Medina 2003), el cual es un mecanismo de defensa contra la herbivoría en las poblaciones naturales. Todo lo anterior está en consonancia con lo expresado por Gepts (2002), quien señaló que los frutales usualmente presentan un síndrome de domesticación incompleto. Lo expuesto, sobre la presencia de espinas en los materiales colombianos y el empleo de un nombre indígena para la especie, llevó a Heiser (1979), a sugerir que la domesticación de la especie comenzó en el país.
La siembra del lulo, en Colombia, se lleva a cabo generalmente con materiales locales, que reciben nombres diferentes o la misma denominación para ecotipos distintos, con asignación del apelativo Lulo de Castilla a poblaciones cultivadas en diversas zonas del país (Chacón, Cardona y Ariza, 1996; Cabezas y Novoa, 2000; Franco et al., 2002; Gómez et al., 2004; Ríos, et al., 2004). El material de propagación se obtiene a partir de semilla extraída por los productores, intercambio con otros agricultores o compra de material vegetativo a partir de viveros locales, sin certificación alguna, con existencia de un sólo cultivar registrado, el lulo La Selva (Bernal, Lobo y Londoño, 1998). En el sentido anterior, Hodgkin et al. (2007), indicaron que en el siglo XX los gobiernos dedicaron esfuerzos para promover la oferta de cultivares mejorados, pese a lo cual, las comunidades locales de los países en desarrollo continúan obteniendo el material para la siembra principalmente a partir de fuentes tradicionales o informales, aspecto que ha sido puntualizado por diversos autores (Gaifani, 1992; Hardon y de Boef, 1993; Mellas, 2000; Tripp, 2001; Bellon y Risopoulos, 2001; Badstue et al., 2002).
Para apoyar la implementación de un programa de oferta de materiales para la siembra, se conformó una colección de la especie cultivada y taxa relacionados, la cual entró a formar parte del Sistema de Bancos de Germoplasma, para la Alimentación y la Agricultura, de la Nación Colombiana. Esta se derivó de las accesiones existentes a partir de expediciones de colecta en diversas zonas, las cuales, de acuerdo con los libros de campo, datos de pasaporte e informes publicados, provienen de los departamentos de Antioquia, Boyacá, Caldas, Cauca, Cundinamarca, Huila, Magdalena, Nariño, Norte de Santander, Putumayo, Quindio, Tolima, Santander y Valle del Cauca (García y García, 1985; Estrada, García y García, 1986; Marín y Hernández, 1988), a las que se adicionaron entradas de otras localidades del país e introducciones a partir de jardines botánicos, investigadores y bancos de germoplasma de otras áreas del mundo (Lobo et al., 2002).
El potencial de utilización de la colección depende del conocimiento que se tenga del conjunto germoplásmico, por diferentes aproximaciones como son los procesos de conocimiento fenotípico y genotípico. Al respecto, se ha afirmado que una de las causas para el uso reducido de los bancos de germoplasma es la falta de información sobre la variabilidad genética de las accesiones en conservación (Ordás, Malvar y de Ron, 1994) y que la utilización depende de una adecuada clasificación de la variabilidad (Van Beuningen y Busch, 1997) y de un conocimiento detallado sobre los atributos presentes en los materiales de las colecciones (Beuselink y Steiner, 1992). Con base en lo anterior, el Plan Mundial de Acción para la Conservación y la Utilización Sostenible de los Recursos Filogenéticos para la Alimentación y la Agricultura, incluyó como una de sus prioridades, la realización de procesos de caracterización y evaluación, dado que la mayoría de las accesiones carecen de esta información, lo cual causa baja utilización de los recursos genéticos, y resulta en costos elevados en la conservación en relación con los beneficios derivados (FAO, 1996). Con algunos de los materiales, incluidos en la colección, se realizaron, en forma previa al estudio actual, dos procesos de evaluación y caracterización morfológica por parte de algunos de los autores de la investigación presente (datos no publicados) y de Sahaza y Henao (2001), en los cuales se encontró polimorfismo morfológico amplio, al igual que una evaluación de la vida de poscosecha de las bayas y su relación con la actividad de la enzima poligalacturonasa (Giraldo y Gil, 2004).
Tomando como referente todo lo anterior, se ejecutó el estudio actual de caracterización y evaluación morfológicas en un conjunto amplio de los materiales de la colección, los que incluyeron la especie cultivada y taxa relacionados de la sección Lasiocarpa, al igual que otras Solanaceae no pertenecientes a la sección anterior, como grupo externo de comparación. Complementariamente, y en forma simultánea, se llevó a cabo un trabajo de caracterización molecular, con los materiales de la investigación presente (Fory, 2005; Fory et al., 2007). Con los resultados de la caracterización actual, presentados y discutidos en el escrito, se busca promover el empleo de la colección, a través del conocimiento de los atributos disponibles requeridos para programas de mejoramiento. Además, se pretende utilizar éstos para la obtención, en principio, de relaciones putativas con los datos obtenidos en el estudio molecular. Esto, puede apoyar la integración de la información molecular y fenotípica, lo cual ha sido enfatizado por Sobral (2002), en la era que él denomina como de la investigación biológica industrial.
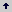
Localización. El trabajo se llevó a cabo en el Centro de Investigación, C.I. La Selva, de la Corporación Colombiana de Investigación Agropecuaria, CORPOICA, ubicado en el municipio de Rionegro, Antioquia a 2120 msnm, con una temperatura promedio de 17°C, una precipitación anual de 1800 mm y una humedad relativa media de 75%. La zona, donde se encuentra el Centro de investigación pertenece a la formación ecológica bosque húmedo montano bajo.
Material biológico. En el estudio se incluyeron en total 116 accesiones, de las dos variedades botánicas del taxón cultivado Solanum quitoense var. quitoense (sin espinas) y S. quitoense var. septentrionale (con espinas), demes de siete especies relacionadas de la sección Lasiocarpa: S. hirtum, S. pseudolulo, S. vestissimum, S. pectinatum, S. sessiliflorum, S. stramonifolium y S. ferox y poblaciones de cinco taxa de la familia Solanaceae, S. capsicoides, S. jilo, S. marginatum, S. mamosum y S. athropurpureum, de secciones diferentes a Lasiocarpa, para su empleo como grupo de referencia.
En la Tabla 1, se incluyen las entradas de la evaluación, por especie, las cuales están vinculadas al Sistema de Bancos de Germoplasma Vegetal de la Nación Colombiana, para la Alimentación y la Agricultura, a cargo de CORPOICA. Los materiales se sembraron por transplante en un lote experimental del C.I. La Selva, con establecimiento de parcelas de cinco plantas por población, a una distancia de 3 m entre surcos y 2 m entre individuos.
Tabla 1. Accesiones de lulo Solanum quitoense y taxa relacionados de la sección Lasiocarpa y otras entidades de Solanaceae, como grupo de comparación (GC), incluidas en el estudio.
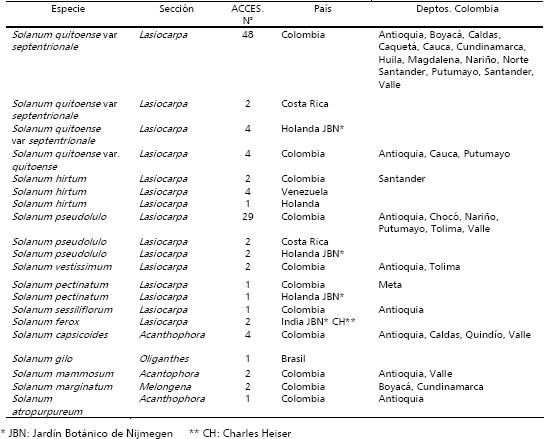
Descriptores y análisis de la información. La información se registró, en forma individual, en cinco plantas por accesión, en plena competencia, mediante la utilización de un listado de descriptores desarrollados inicialmente por investigadores de la Universidad Nacional de Colombia, sede Palmira, el cual fue revisado y modificado por el equipo de Recursos Genéticos de CORPOICA, del C.I. La Selva, Rionegro, Antioquia Este comprende 83 variables, de las cuales 58 son de naturaleza cualitativa y 25 cuantitativa. En la Tabla 2, se incluye el listado de atributos, con la categorización, de cada uno de los caracteres evaluados. Esta se refiere, en el caso de los cualitativos a variables binarias, multiestado con orden lógico y multiestado con orden no lógico y en caso de las cuantitativas, características continuas y discretas.
Tabla 2. Variables cualitativas y cuantitativas registradas en la colección colombiana de lulo Solanum quitoense, especies relacionadas de la sección Lasiocarpa y otros taxa de la familia Solanaceae
Los datos obtenidos se incorporaron a una hoja electrónica del programa Excel, con diferenciación de éstas como cualitativas y cuantitativas.
En el caso de los rasgos cualitativos, codificados numéricamente, para cada uno de éstos, se obtuvo la moda de la población, el número de estados polimórficos, el porcentaje de polimorfismo por característica y el número promedio de estados por atributo (morfoalelos).
Con los aspectos cualitativos con variabilidad, se llevó a cabo un análisis de agrupamientos, con base en matrices de similitud de las modas, entre pares de materiales, a través del empleo del Coeficiente Simple de Similaridad, propuesto por Sokal y Michener (1958). Con esto se generó un fenograma, mediante el algoritmo de las medias no ponderadas UPGMA, con el paquete de programación NTSYS, versión 2.0 Rohlf (1998).
En el caso de las características cuantitativas, se realizó un análisis univariado para cada una de éstas, el cual permitió detectar el promedio, la desviación estándar, el coeficiente de variación, y los valores máximos y mínimos por rasgo y se realizó un estudio de Componentes Principales con el fin de determinar las características con mayor contribución a la explicación de la diversidad de ésta índole. Luego, con aquellas que exhibieron variabilidad, se efectuó un estudio de agrupamientos, con valores convertidos a unidades estándar, con el fin de que cada variable tuviera un peso igual en el procedimiento (Sneath y Sokal, 1953; Crisci y López, 1983). Este, se completó con el uso del coeficiente de distancia, conocido como la Diferencia Promedio entre Atributos, propuesto por Cain y Harrison (1958), con obtención de un fenograma cuantitativo por el algoritmo de las medias no ponderadas UPGMA, versión 2.0 (Rohlf, 1998).
También se realizó un análisis conjunto de variabilidad cuantitativa y cualitativa, para lo cual se empleo el coeficiente de similitud propuesto por Gower (1971), el cual permite unir los atributos de las categorías anteriores. Con este coeficiente, se obtuvo un fenograma a través del empleo del algoritmo de las medias no ponderadas UPGMA.
Variabilidad cualitativa. Como puede apreciarse en la Tabla 3, se encontró polimorfismo para los 58 atributos cualitativos registrados, tomando en consideración todas las especies caracterizadas, al igual que los taxa de la sección Lasiocarpa. En el caso de las variedades botánicas septentrionale y quitoense, del taxón cultivado Solanum quitoense, las cuales se caracterizan por la presencia de espinas en el primer clade y su ausencia en el segundo (Heiser, 1972; Schultes y Romero-Castañeda, 1962; Schultes y Cuatrecasas, 1958), se detectó variabilidad en 55 de los 58 caracteres de la categoría anterior, En el sentido anterior, Sahaza y Henao 2001, reportaron amplia variabilidad cualitativa en materiales de la colección colombiana de lulo, lo cual concuerda con lo obtenido en el trabajo presente. El resultado no coincide con informes de baja variabilidad morfológica reportada en poblaciones de lulo o naranjilla del Ecuador (Whalen, Costich y Heiser, 1981), lo cual fue atribuido a un efecto fundador asociado con la dispersión antrópica del material a partir de la parte Central de Colombia, lugar que es considerado por Heiser (1969) como el sitio de domesticación de la especie; patrón que se extiende desde el sur de Colombia hacia el Ecuador (Whalen, Costich y Heiser, 1981).
Tabla 3. Polimorfismo cualitativo encontrado en lulo, Solanum quitoense, taxa de la sección Lasiocarpa y otras especies de Solanum, Solanaceae, en 330 estados de 58 características.
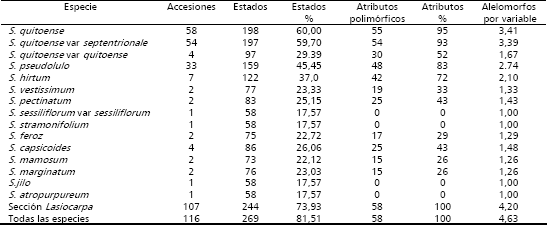
El potencial de utilización de cualquier colección de germoplasma depende de la variabilidad existente en ésta. En el contexto, el conjunto de materiales estudiados de la sección Lasiocarpa, además de presentar polimorfismo para todos los atributos cualitativos, exhibió el 73,9% de todos los estados incluidos en el listado de descriptores, con un promedio de 4,2 morfoalelos por variable (Tabla 3), lo que indica que en el germoplasma existe un rango de atributos que posibilitan atender las demandas de atributos específicos requeridos por los investigadores, agricultores y consumidores. La utilización de la variabilidad cualitativa, de la colección caracterizada, parte de la búsqueda de características importantes dentro de las dos variedades botánicas de la especie S. quitoense septentrionale y quitoense y luego en los taxa de la sección Lasiocarpa, compatibles con ésta. En el primer caso, en el taxón cultivado, se aprecio en las accesiones de sus dos variedades botánicas, que éstas presentaban el 60% de los atributos totales descritos, con 3,4 variantes por cada uno de éstos. En cuanto a las especies relacionadas de la sección Lasiocarpa, una primera alternativa, como fuente de características morfológicas, es la entidad biológica Solanum hirtum, la cual presentó diversidad en 42 de las variables, con 2,1 morfoalelos por aspecto y 37,0% de las variantes totales (Tabla 3). Esta es compatible con S. quitoense, con obtención de híbridos fértiles (Whalen, Costich y Heiser, 1981; Heiser, 1985, 1989; Lobo, 2007). Otro taxón, a partir del cual se pueden transferir características, por métodos convencionales, es S. pseudolulo, especie endémica de Colombia (Whalen, Costich y Heiser, 1981), en el cual se obtuvo variabilidad en 48 caracteres, con 2,7 morfoalelos por propiedad y 45,4% de los estados (Tabla 3); el traspaso de atributos de S. pseudolulo, a S. quitoense es posible realizarlo mediante puentes genéticos a través de S. hirtum, los cuales, de acuerdo con Heiser (1989), precisan de rescate de embriones, con reporte de obtención de un híbrido entre S. hirtum y S. pseudolulo, sin la necesidad de llevar a cabo el procedimiento (Lobo, 2007).
También es factible el traslado de atributos genéticos, al lulo, a partir de S. vestissimum, entidad que exteriorizó diversidad en 19 características con 1,3 variantes por aspecto y 23,3% de los morfoalelos totales, en las dos accesiones de ésta, incluídas en la caracterización. El taxón puede hibridarse con S. quitoense, lo cual demanda rescate de embriones (Heiser, 1989). La utilización de esta entidad implica colecta, a partir de diversas zonas, para incrementar su representatividad en el conjunto de germoplasma en conservación en Colombia. En el país, la especie ha sido reportada en la Cordillera Oriental desde Boyacá hasta Norte de Santander, en la Cordillera Central en Antioquia y en la Sierra Nevada de Santa Marta (Whalen, Costich y Heiser, 1981). Otro cruzamiento viable del lulo se logra con S. sessiliflorum, entidad de la cual solo se incluyó una población; la hibridación demanda el empleo, de S. quitoense como progenitor femenino, con logro de híbridos sin semilla, estériles (Heiser, 1989, 1993), lo que implica utilización de éstos como tal. Al respecto, en el Ecuador se siembran dos híbridos interespecíficos: Puyo y Palora, entre los dos taxa anteriores, los cuales se propagan vegetativamente (Heiser, 1993).
Cabe señalar que las características cualitativas corresponden en alto grado a la expresión de alelos de genes específicos, o sea, que pueden emplearse como marcadores genéticos (Van Hintum, 1995) y que dichos alelos marcan igualmente genes ligados a éstos, por lo cual la variabilidad se extiende a características genéticas ubicadas a corta distancia de los loci de atributos cualitativos en los cromosomas (Medina y Lobo, 2001; Rosso, Medina y Lobo, 2004;), cuya utilización se favorece con la presencia de desequilibrio de ligamiento.
En la Figura 1, se incluye el fenograma cualitativo obtenido a partir del coeficiente de similaridad, con inclusión del valor de disimilaridad en el eje de las Y (1- similaridad), para efectos de la discusión. En éste puede verse una mayor variabilidad morfológica en el conjunto de accesiones del taxón cultivado S. quitoense, considerando las formas septentrionale y quitoense, en comparación con los de S. pseudolulo, S. vestissimum, y S. hirtum. con separación de los grupos de las dos variedades botánicas de S. quitoense, entre las cuales se ubicaron los conglomerados de los taxa silvestres mencionados. Esto difiere de los resultados obtenidos por Fory et al. (2007), quienes indicaron, en un estudio realizado con marcadores AFLP, con materiales de la investigación presente, una mayor variabilidad de las tres especies no cultivadas S. pseudolulo, S. vestissimum y S. hirtum, que la obtenida con S. quitoense. Otro aspecto derivado del árbol de variabilidad es el hecho de éste tuvo valor taxonómico desde la óptica de los agrupamientos por especie, en la sección Lasiocarpa, con ligero intercalamiento de accesiones de diferentes especies, con excepción de S. quitoense, en el cual, como se anotó, no hubo continuidad entre los clades de las dos variedades botánicas. Adicionalmente, no fue evidente la separación espacial entre los taxa andinos: S. quitoense, S. pseudolulo, S. vestissimum, S. hirtum y S. pectinatum, como conjunto, al de las entidades biológicas de la Amazonia: S. sessiliflorum y S. stramonifolium, Al respecto, en el estudio de Fory et al. 2007, se obtuvo discriminación sistemática por taxa e igualmente entre las especies Andinas y Amazónicas.
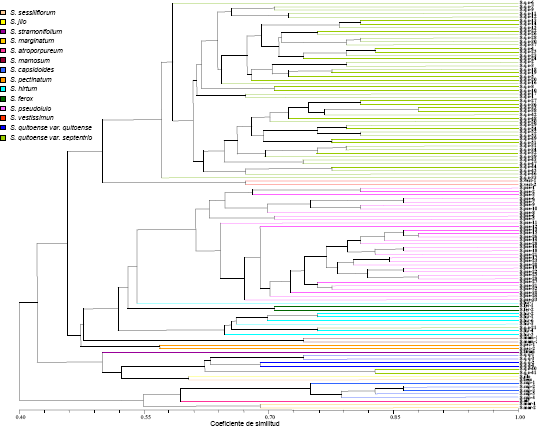
Figura 1. Fenograma cualitativo de la colección de lulo Solanum quitoense, taxa relacionados de la sección Lasiocarpa y otras Solanaceae
El distanciamiento morfológico, de las ramas de las variedades botánicas septentrionale y quitoense, puede atribuirse al hecho de que los materiales de la segunda entidad, provienen de procesos de domesticación, iniciados con la selección de materiales sin espinas (Heiser, 1979), lo cual causó un efecto fundador, con selección antrópica posterior, por atributos importantes y para su siembra en condiciones de mayor luminosidad que las soportadas por los materiales de la forma septentrionale. Las diferencias de comportamiento a la intensidad lumínica entre los dos materiales han sido informadas por Medina (2003) y Medina et al. (2006).
Por su parte, la accesión asiática de la sección, S. ferox, se entremezcló con S. hirtum. Al respecto, inicialmente, se consideraron dos especies de la sección Lasiocarpa en Asia, estas correspondían a S. lasiocarpum y S. repandum (Whalen, Costich y Heiser (1981); de éstas, Whalen, Costich y Heiser (1981) y Heiser (1987) indicaron que la primera, estaba íntimamente relacionada con S. candidum, taxón de los Andes, no incluido en el estudio actual, con el cual se obtienen híbridos fértiles con S. lasiocarpum. La denominación anterior de S. lasiocarpum era S. ferox, nombre cuya reinstalación se propuso para esta entidad biológica conjuntamente con S. repandum (Heiser, 1996b), con información, por parte de Bohs (2004), a partir de un estudio, con ADN de los cloroplastos, de que S. lasiocarpum y S. repandum eran muy similares y que agrupaban con las especies del nuevo mundo S. candidum y S. pseudolulo. Con relación al agrupamiento de S. ferox con S. hirtum, Whalen y Caruso (1983) indicaron, como consecuencia de una investigación filogenética y de congruencia entre caracterizaciones morfológicas y moleculares, que las especies S. lasiocarpum, S candidum, S. quitoense,S. hirtum y S. pseudolulo, agrupaban en una misma rama. La ubicación de S. ferox, intermezclado en un extremo del clade de S. hirtum, podría atribuirse a la amplia variabilidad morfológica de éste último, por ser la especie Andina, de la sección Lasiocarpa, de más amplia distribución y variabilidad, la cual se extiende desde México, a través de América Central, donde posiblemente fue introducida, hasta Colombia y Venezuela, con demes en Trinidad (Whalen, Heiser y Costich, 1981).
El potencial de utilización de las especies S. hirtum, S. pseudolulo y S vestissimum, para transferir atributos cualitativos no presentes en S. quitoense, pude apreciarse por las disimilitud entre los conjuntos de las especies en mención, incluidas, en el fenograma de la Figura 1. En este se puede apreciar una no semejanza del 58% entre los agrupamientos de las dos variedades botánicas de S. quitoense y los clades de S. hirtum, S. pseudolulo y S. vestissimum. Como se indicó hay compatibilidad genética entre S. quitoense y S. hirtum (Whalen, Costich y Heiser, 1981; Heiser, 1985, 1989; Lobo, 2007) y se puede transferir genes de S. pseudolulo a S. quitoense por la vía de un puente genético entre S. hirtum y S. pseudolulo (Heiser, 1989; Lobo, 2007), aspecto que se potencia por una disimilitud del 48% entre las dos últimas especies por lo cual, los híbridos entre ellas pueden estar enriquecidos con atributos morfológicos y loci ligados estrechamente a éstos, presentes en las dos entidades. Complementariamente, hay posibilidades de incremento de la variabilidad morfológica, de S. quitoense, mediante cruzamientos interespecíficos con S. vestissimum; éstos son factibles con el empleo de rescate de embriones (Heiser, 1989).
La máxima disimilitud, entre accesiones de S. quitoense, fue del 58% (Figura 1); éste valor unido a 95% de atributos variables, con la presencia del 60% de los atributos cualitativos totales, indica que hay rearreglos diferentes de características de ésta índole en los materiales del taxón. Lo anterior señala posibilidades de encontrar combinación de caracteres deseables en algunas accesiones. Esto facilitaría su evaluación per se en zonas con condiciones ecológicas similares a las de colecta de los materiales con conjuntos de atributos demandados, con base en los datos de pasaporte disponibles. En el caso presente no se dispone de información de pasaporte en algunas de las poblaciones evaluadas o ésta es parcial. Las deficiencias en datos de la índole anterior fueron puntualizadas en el Primer Informe del Estado de los Recursos Filogenéticos para la Alimentación y la Agricultura (FAO, 1996a) y en la formulación del Plan Mundial de Acción para la Conservación y la Utilización de los Recursos Genéticos para la Alimentación y la Agricultura (FAO, 1996b). Al respecto, Plucknett et al. (1987), estimaron que al menos el 65% de las accesiones en bancos de germoplasma carecen de este tipo de conocimiento y Williams (1989), Beuselinck y Steiner (1992), Van Hintum y Knüpffer (1995) y Hazekamp (2002) señalaron limitaciones y errores en el la reseña de éstos.
En el fenograma, Figura 1, se puede apreciar que de las especies de la familia Solanaceae, empleadas como testigos de comparación; S. capsicoides y S. atropurpureum, de la sección Acanthopora y S. marginatum de la sección Melongena, se ubicaron al final del dendrograma, con agrupamientos intraespecíficos, en el caso de los taxa con más de una entrada. Por su parte las entidades S. mammosum de la sección Acanthopora y S. gilo de la Oliganthes, se localizaron, con agrupamientos intraespecíficos de sus demes y con algún intercalamiento con materiales de Lasiocarpa. Las asociaciones intraespecíficas y la ubicación al final del árbol morfológico, con las mayores disimilitudes, de algunas de ellas, brindan confiabilidad al procedimiento jerárquico realizado a partir de las variables cualitativas.
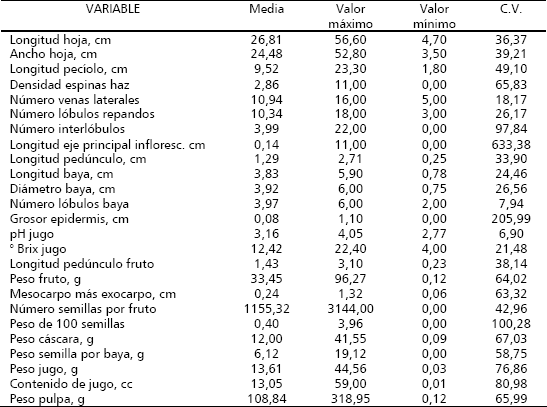
Variabilidad cuantitativa. En la Tabla 4, se incluye la información de promedios, valores máximos y mínimos obtenidos con las características cuantitativas registradas en el estudio con la especies de la sección Lasiocarpa. Como puede apreciarse todos los atributos exhibieron polimorfismo, con coeficientes de variabilidad que fluctuaron entre 6,9% para pH del jugo y 633,4% para la longitud del eje principal de la inflorescencia.
Tabla 4. Promedio, valor máximo, valor mínimo y coeficiente de variación (C.V.) para cada una de las variables cuantitativas registradas en la colección de lulo, Solanum quitoense y especies relacionadas de la sección Lasiocarpa.
En un análisis de componentes principales, los 8 primeros, exhibieron valores característicos superiores a uno, con explicación del 79,2% de la variabilidad total cuantitativa y con contribución de un conjunto amplio de atributos registrados, a la diversidad de la colección de lulo, especies relacionadas de la sección Lasiocarpa y taxa de la familia Solanaceae de las secciones Oliganthes, Acantophora y Melongena. El procedimiento permitió visualizar que las variables con mayor contribución a la explicación de la variabilidad, estuvieron relacionadas con atributos del fruto; de éstos, los pesos total, de jugo, y de la pulpa fueron los de mayor significado para el primer componente, con una contribución del 41,6% a la diversidad total; con aporte de atributos de las bayas a los 7 primeros componentes, los cuales explicaron el 70,5% de polimorfismo. Esto conduce a pensar, en el caso de las especies silvestres, en el papel del fruto en la producción de semilla y procesos de dispersión, para la supervivencia de las especies y en el de los frutales cultivados, en selección antrópica, por ser el órgano de valor de la planta. Al respecto, Brown y Hodgkin (2007) escribieron, con relación a los materiales locales, que los agricultores, en forma tradicional, han manipulado, seleccionado y usado las diferencias que ellos perciben entre y dentro de las especies, las cuales pueden ser, entre otras, de morfología, productividad, calidad, resistencia a pestes y variación útil, lo que puede nos ser evidente para el común de la gente.
El fenograma cuantitativo, derivado de coeficientes de distancia, se incluye en la Figura 2. En este puede apreciarse algún agrupamiento por especies, con accesiones que se intercalan en los conjuntos de otros taxa. A diferencia del árbol cualitativo, las poblaciones de S. quitoense var. quitoense agruparon con las de S. quitoense var. septentrionale. Los conglomerados cuantitativos y cualitativos señalaron una baja coincidencia a nivel de los taxa de la sección Lasiocarpa. Al respecto se ha indicado que los atributos cuantitativos son afectados por el ambiente y que los cualitativos están relacionados con genes con interferencia ambiental baja o nula (Van Hintum, 1995). Resultados similares, de baja congruencia cuantitativa y cualitativa, han sido obtenidos en estudios previos de caracterización y evaluación morfológica con otras especies (Medina y Lobo, 2002; Rosso, Medina y Lobo, 2004; Vásquez, Medina y Lobo, 2004).
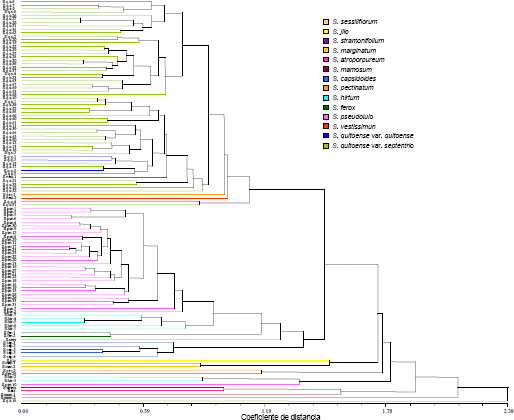
Figura 2. Fenograma cuantitativo de la colección de lulo Solanum quitoense, taxa relacionados de la sección Lasiocarpa y otras Solanaceae.
Desde la óptica del aprovechamiento de los recursos genéticos, para incremento de la productividad, las distancias cuantitativas son importantes para la selección de parentales, con literatura sobre el efecto de la distancia morfológica en la heterosis de los híbridos obtenidos (Cox y Murphy, 1990; Riday et al., 2003; Geleta, Labuschagne y Vijoen, 2004). Tomando como referente lo anterior, el dendrograma, asociado con atributos cuantitativos, podría emplearse en la búsqueda de heterosis en la generación F1. Esto parte de la búsqueda de vigor híbrido dentro de la especie cultivada S. quitoense, con utilización de los materiales de las variedades botánicas septentrionale y quitoense. Para lo anterior, el fenograma es una herramienta valiosa. A partir de éste se podrían seleccionar parentales distantes, de los cuales el más alejado de todas las accesiones es Sqs 14, material colectado en Guarne, Antioquia, perteneciente a la forma septentrionale, el cual no agrupó en el conglomerado de los S. quitoense y exhibió la máxima distancia cuantitativa con todos los materiales estudiados, con ubicación en la rama jerárquica superior del fenograma. Sin embargo, es importante estudiar la separación óptima, entre accesiones, para el logro de vigor híbrido ya que se ha indicado que hay una distancia a partir de la cual se produce depresión en éste, lo que se conoce como disgenesia híbrida (Spillane y Gepts, 2001). Otra alternativa es la acumulación de Atributos de Loci Cuantitativos (QTL), mediante hibridación interespecífica, aspecto que ha sido realizado con éxito en diversas especies. Así, Fulton et al. (1998), reportaron resultados de trabajos de hibridación interespecífica en tomate y en arroz, con presencia de un 25% de alelos favorables, en los cruzamientos con la primera especie, provenientes del taxón no cultivado Lycopersicon peruvianum e incremento en el rendimiento en la segunda especie por parte de los atributos cuantitativos derivados del taxón silvestre Oryza rufipogum. Esto fue atribuido por Miflin (2000), al hecho de que la domesticación de los cultivos se ha llevado a cabo con bases genéticas estrechas de las poblaciones silvestres. En consonancia con lo anterior, Lobo (datos sin publicar), ha obtenido productividades elevadas en hibridaciones entre S. quitoense y S. hirtum, en procesos iniciados por el autor (Lobo et al., 2002; Lobo, 2004, 2007), para crear una base heredable amplia como sustento de programas de entrega de cultivares mejorados de la especie. En el contexto, también se considera importante evaluar el potencial de los taxa S. pseudolulo y S. vestissimum, para aportar loci de atributos cuantitativos relacionados con productividad y adaptación.
Variabilidad cualitativa y cuantitativa. En la Figura 3, se incluye el fenograma obtenido con el procedimiento de Gower (1971), basado en un coeficiente de similitud que permite analizar datos mixtos cualitativos: binarios y multiestado e información cuantitativa, generando medias de proximidad entre pares de individuos. En éste se consiguió una separación entre los taxa, de la sección Lasiocarpa de los Andes: S. quitoense, S. vestissimun, S. pseudolulo, S. hirtum y S. pectinatum, y las especies Amazónicas: S. sessiliflorum y S. stramonifolium, las cuales se ubicaron en el conglomerado superior del árbol obtenido, intercaladas con la accesión de S. gilo, conocida también como S. aethoipicum, planta comestible cultivada en el Africa por sus frutos (Fatokun, 1989), la cual está relacionada en forma cercana con la berenjena (Isshiki y Taura, 2003). De ésta se incluyó, como testigo de comparación, una accesión obtenida en Brasil, país al cual se introdujo y siembra este taxón. En consonancia con lo precedente Fory et al. (2007) indicaron separación, entre las especies, de la sección Lasiocarpa, de los Andes y la Amazonía, con materiales del trabajo actual, mediante el dendrograma obtenido con marcadores AFLP.
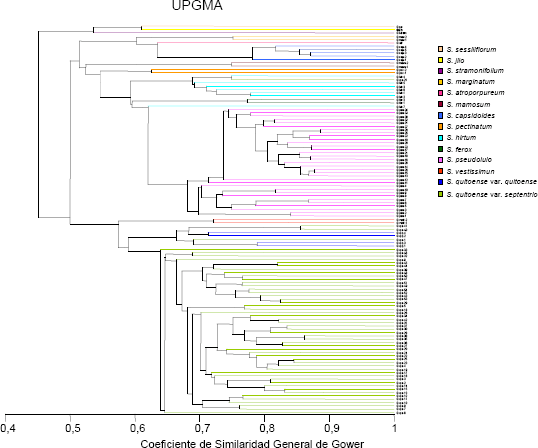
Figura 3. Fenograma cualitativo-cuantitativo de la colección de lulo Solanum quitoense, taxa relacionados de la sección Lasiocarpa y otras Solanaceae.
Igualmente, en el fenograma es evidente que el procedimiento tiene valor taxonómico a nivel intraespecífico e interespecífico, tanto en los taxa de la sección Lasiocarpa, como en los de las entidades biológicas de las otras secciones, con agrupamiento de los tres taxa de la sección Acanthophora: S, atropurpureum, S. capsicoides y S. mammosum. El único caso de intercalamiento de materiales de dos taxa, en el análisis realizado con el procedimiento de Gower (1971), fue el de las dos accesiones de S. ferox dentro del conglomerado de S. hirtum, lo cual también ocurrió en el fenograma cualitativo, con conformación de un conjunto entre las dos entidades en el árbol cuantitativo sin mezcla de los materiales de las dos especies. El valor sistemático interespecífico fue reportado igualmente por Fory et al. (2007), en el estudio molecular. En el árbol logrado con los dos tipos de información, se apreció una ubicación topológica cercana de S. sessiliflorum, S. stramonifoliumy S. pectinatum, con amplia disimilitud entre éstos, con base en una accesión en cada uno de los dos primeros taxa y dos en el último. Al respecto, Bohs (2004) indicó que, en un estudio filogenético de las sección Lasiocarpa, que las tres especies formaban uno de los tres clades de la sección.
El aspecto que no fue evidente en el estudio cualitativo-cuantitativo, fue la separación entre las variedades botánicas septentrionale y quitoense de S. quitoense, las cuales exhibieron intercalamiento, patrón que se obtuvo igualmente en el árbol cuantitativo, con agrupamiento de las dos variedades botánicas con otras taxa en el cualitativo. En contraste con lo obtenido en el estudio actual, Medina y Lobo (2001), indicaron, en un estudio realizado con el tomate pajarito Lycopersicon esculentum var. cerasiforme, separación de las variedades botánicas cerasiforme y esculentum, al emplear datos cualitativos y cuantitativos en forma conjunta, lo cual no se obtuvo con las dos categorías en forma aislada.
Whalen y Caruso 1983, en estudios hechos con atributos morfológicos e isoenzimáticos con especies de la sección Lasiocarpa, a través del empleo de metódos fenéticos y cladísticos, indicaron la obtención de dendrogramas muy similares, en forma independiente de la metodología empleada y afirmaron que las clasificaciones fenéticas, utilizadas en la investigación presente, eran ligeramente más estables que las cladísticas. Igualmente, los autores señalaron que S. stramonifolium,y S. sessiliflorum eran ramas aisladas de la sección Lasiocarpa, aspecto, este último, que fue evidente en el fenograma cualitativo-cuantitativo.
La mayor consistencia sistemática, aparente, del fenograma cualitativo-cuantitativo, indica que este puede servir para la selección de posibles parentales con estados cualitativos deseados y que agreguen loci cuantitativos de productividad, ya que éste tiene tanto la información de de atributos cuantitativos, como de segmentos de esta índole, ligados a los loci cualitativos. Esto puede permitir sumar loci con genética aditiva para rendimiento, lo que es factible obtener tanto dentro de la especie cultivada S. quitoense var septentrionale y quitoense, como en híbridos interespecíficos de ésta con los taxa S. hirtum, S. pseudolulo y S. vestissimum, entidades con las cuales es factible lograr transferencia de genes (Whalen, Costich y Heiser, 1981; Heiser, 1985, 1989; Lobo, 2007), sin empleo de procedimientos de transformación genética. Al respecto, Bernardillo, Heiser y Piazzano (1994), indicaron que la diferenciación morfológica entre las especies de la sección Lasiocarpa, no siempre ocurrió con divergencia cromosómica. Esto indica las posibilidades de realizar hibridación interespecífica entre algunos de los taxa, sin dificultades mayores. Como se señaló, es importante determinar la distancia óptima para la obtención de valores heteróticos, para evitar la presencia de disgenesia híbrida en los cruzamientos, tópico puntualizado por Spillane y Gepts (2001).
Se encontró polimorfismo cualitativo para los 58 atributos registrados, a nivel de todos los materiales, al igual que en las accesiones de especies de la sección Lasiocarpa. En esta última se detectaron 4,2 morfoalelos por característica y 74% de los estados totales del descriptor.
Solanum quitoense, exhibió en las poblaciones de sus dos variedades botánicas, 55 caracteres cualitativos con diversidad, con 3,4 alelomorfos por variable, lo cual señala polimorfismo disponible para procesos de mejoramiento.
Las especies S. hirtum, S. pseudolulo y S. vestissimum , exhibieron 42%, 48% y 19% de caracteres cualitativos variables, con 2,1, 2,7 y 1,3 alelomorfos por característica. Esto señala el potencial de búsqueda de estados deseables en estos taxa, no presentes en el lulo para su transferencia a la entidad biológica.
En el estudio de conglomerados, se obtuvo una mayor variabilidad morfológica de S. quitoense, en comparación S. hirtum, S. pseudoluloy S. vestissimum.
El árbol cualitativo, no indicó una separación clara entre los taxa de Lasiocarpa originarios de los Andes de aquellos provenientes de la Amazonía.
Todas las variables cuantitativas exhibieron variabilidad, con una contribución importante de los atributos del fruto a la diversidad total, lo cual fue evidente a través de un análisis de componentes principales.
Se apreció una amplia dispersión de las accesiones en el fenograma cuantitativo, con agrupamiento e intercalamiento de los materiales de las dos variedades botánicas de S. quitoense y una población de la entidad biológica, Sqs 14, que exhibió la máxima distancia con todos los demes estudiados.
Los agrupamientos cualitativo-cuantitativos, obtenidos con el coeficiente de Gower, indicaron valor taxonómico a nivel inter e intraespecífico y separación, dentro de la sección Lasiocarpa, de los taxa Andinos y los Amazónicos, aspecto, este último, que no fue evidente en los conglomerados obtenidos con cada uno de los dos tipos de variables.
El fenograma cualitativo-cuantitativo, por su mayor consistencia sistemática, puede emplearse para la selección de parentales con atributos cualitativos deseados y amplia distancia, lo cual suma a los loci cuantitativos, otros posibles segmentos de ésta índole ligados estrechamente a los cualitativos, con desequilibrio de ligamiento.
La presente investigación se realizó gracias al apoyo financiero de COLCIENCIAS, en el marco del proyecto: Conocimiento de la variabilidad genética del lulo (Solanum quitoense Lam.), como apoyo a la implementación de programas de mejoramiento, Código 7106-12-11582, contrato 153, 2001.
Arias, C. L., L. Támara y F. Arbeláez. 2006. Apuesta exportadora agropecuaria 2006-2020. República de Colombia. Ministerio de Agricultura y Desarrollo Rural, 119 p. [ Links ]
Badstue, L.B., M. Bellon, X. Juárez, I. Manuel and A.M. Solano. 2002. Social relations and seed transactions among small-scale maize farmers in the Central Valleys Oaxaca, Mexico: preliminary findings. CIMMYT, Mexico, D.F. 21 p. (Economic Working Paper 02-02.) [ Links ]
Bellon, M.R. and J.Risopoulos. 2001. Small-scale farmers expand the benefits of improved maize germplasm: A case study from Chiapas, Mexico . World Dev. 29(5):799-811. [ Links ]
Bernal, J., M. Lobo y M. Londoño. 1998. Documento de presentación del material Lulo La Selva. Corpoica, Rionegro, Antioquia, Colombia. 77p. [ Links ]
Bernardello, L.M. C.B. Heiser and M. Piazzano. 1994. Karyotypic studies in Solanum section Lasiocarpa (Solanaceae). Amer. J. B. 81(1):95-103. [ Links ]
Beuselinck, P.R. and J.J. Steiner. 1992. A proposed framework for identifying quantifying and using plant germplasm resources. Field Crops Res. 29:261-272. [ Links ]
Bohs, L. 2004. A chloroplast DNA phylogenie of Solanum section Lasiocarpa. Systematic Bot. 29(1):177-187. [ Links ]
Brown, A.H.D. and T. Hodgkin. 2007. Measuring, managing, and maintaining crop genetic diversity on farm. pp. 13-33. In: Jarvis, D.I., C. Padoch and H.D. Cooper. (eds.). Managing biodiversity in agricultural ecosystems. bioversity. Columbia University Press, New York, USA . [ Links ]
Cabezas, M. and D. Novoa. 2000. Efecto de la remoción de hojas y frutos en la relación fuente demanda en lulo (Solanum quitoense Lam.). pp. 176-181. En: Memorias III Seminario Frutales de Clima Frío Moderado. Centro de Desarrollo Tecnológico de Frutales, CDTF, Manizales, Colombia. [ Links ]
Cain, A.J. and G.A. Harrison. 1958. An analysis of the taxonomists judgement of affinity. Proc. Zool. Soc. London. 131:85-98. [ Links ]
Cárdenas, W., M.L. Zuluaga and M. Lobo. 2004. Latencia en semillas de lulo (Solanum quitoense Lam.) y tomate de árbol (Cyphomandra betacea (Solanum betaceum) Cav. Sendt.) como aspecto básico para la conservación y el monitoreo de viabilidad de las colecciones. Plant Genet. Res. Newsl. (139):31-41. [ Links ]
Chacón, R.C., M.M. Cardona y H.J. Ariza, 1996. Caracterización físico-química de tres híbridos de lulo y lulo de castilla, producido bajo sol y sombra. pp. 81-87. En: En: Memorias I Seminario Frutales de Clima Frío Moderado. Centro de Desarrollo Tecnológico de Frutales, CDTF, Manizales, Colombia. [ Links ]
Cox T.S. and J.P. Murphy. 1990. The effect of parental divergence on F2 heterosis in winter wheat crosses. Theor. Appl. Genet. 79(2):169-171. [ Links ]
Crisci, J.V., y M.F. López. 1983. Introducción a la teoría y práctica de la taxonomía numérica. Serie de Biología. Secretaria General de la OEA. Programa Regional de Desarrollo Científico y Tecnológico, Washington, D.C. 132 p. (Monografía No 26). [ Links ]
Estrada, E.I., H.E. García y M. García. 1986. Colección y establecimiento de un banco de germoplasma en lulo Solanum quitoense Lam. y especies relacionadas en el suroccidente colombiano. p. 25-27. En: Memorias III Seminario de Cultivos Promisorios. Universidad Nacional de Colombia, Medellín. [ Links ]
Fatokun, C.A. 1989. Cytogenetical studies of the F1 interspecific hybrid between Solanum aethiopicum and S. gilo Raddi. Cytologia. 54:425-428. [ Links ]
Fory, P. 2005. Caracterización y análisis molecular de la diversidad genética de la colección colombiana de lulo (Solanum quitoense Lam) y seis especies relacionadas de la sección Lasiocarpa. Tesis Magíster en Ciencias, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Palmira. 78 p. [ Links ]
Fory, P., I. Sánchez, A. Bohórquez, H. Ramírez, C.I. Medina and M. Lobo, 2007. Genetic variability of a colombian collection of lulo (Solanum quitoense Lam.) and related species of section Lasiocarpa. Plant Genet. Resources Newsl. (Submitted). [ Links ]
Franco, G., J. Bernal, J.L. Gallego, J.E. Rodríguez, N. Guevara, M. Giraldo y M. Londoño, 2002. Generalidades del cultivo del lulo. Asohofrucol, Corpoica, Fondo Nacional de Fomento Hortofrutícola, Bogotá, Colombia. 97 p. [ Links ]
Fulton, T.M., T. Beck-Bunn, D. Emmatty, Y. Eshed, J. Lopez, C. Petiard, J. Uhlig, D. Zamir, and S.D. Tanksley, 1997. QTL analysis of an advanced backcross of Lycopersicon peruvianum to the cultivated tomato and comparisons with QTLs found in other wild species. Theoretical Appl. Genet. 95(5):881-894. [ Links ]
Gaifani, A. 1992. Developing local seed production in Mozambique . En: Cooper, D., R. Vellvé and H. Hobbelink (eds.). Growing diversity: genetic resources and local food security. Intermediate Technol. Publ., London. 195 p. [ Links ]
García, E.H. y M.A. García. 1985. Colección y establecimiento de un banco de germoplasma en lulo, Solanum quitoense Lam., y especies relacionadas en el suroeste colombiano. Trabajo de grado Facultad de Ciencias Agropecuarias Universidad Nacional de Colombia. Palmira. 100 p. [ Links ]
Geleta, L.F., M.T. Labuschagne and C.D. Viljoen. 2004. Relationship between heterosis and genetic distance based on morphological traits and AFLP markers in pepper. Plant Breed. 123(5): 467-473. [ Links ]
Gepts, P. 2002. A comparison between crop domestication, classical plant breeding and genetic engineering. Crop Sci. 42(6):1780-790. [ Links ]
Giraldo, B. E. y M. Gil. 2004. Cuantificación de la vida de almacenamiento de los frutos de Solanum quitoense Lam. y su relación con la actividad de la enzima poligalacturonasa. Trabajo de grado Facultad de Ciencias Agropecuarias. Universidad Nacional de Colombia. Medellín. 22 p. [ Links ]
Gómez, L.M., J.M. Rojas, G.E. Aristizábal, A.E. Peñuela, M.C. Chaparro, A. López, y J.M. Naranjo. 2004. Caracterización y normalización del lulo de castilla (Solanum quitoense Lam.). pp. 161-168. En: Memorias V Seminario Nacional e Internacional de Frutales. Centro de Desarrollo Tecnológico de Frutales, CDTF, Manizales, Colombia. [ Links ]
Gower, J.C. 1971. A General coefficient of similarity and some of its properties. Biometrics 27(4): 857-871. [ Links ]
Hardon, J. and de Boef, W. 1993. Linking farmers and breeders in local crop development. pp. 64-71. In: Boef, W. Amanor, K. Wellard, K. and Bebbington, A., eds. Cultivating knowledge: genetic diversity, farmer experimentation and crop research. Intermediate Technol. Publ., London. [ Links ]
Hazekamp, Th. 2002. The potential role of passport data in the conservation and use of plant genetic resources. pp. 185-194. In: (Engels, J.M.M., R. Ramantha, A.H.D. Brown, M.T. Jackson (eds.). Managing plant genetic diversity. Chapter 18. Oxford University Press. 514 p. [ Links ]
Heiser, C.B. 1969. Nightshades: the paradoxical plants. W.H. Freeman, San Francisco, USA . 301p. [ Links ]
Heiser, C.B. 1972. The relationship of the naranjilla S. quitoense. Biotropica. 4(2):77-84. [ Links ]
Heiser, C.B. 1979. Origins of some cultivated new world plants. Annu. Rev. Ecol. Syst. 10:309-326. [ Links ]
Heiser, C.B. 1985. Ethnobotany of the naranjilla (Solanum quitoense) and its relatives. Econ. Bot. 74:1045-1048. [ Links ]
Heiser, C.B. 1987. Origins of Solanum lasiocarpum and S. repandum. Amer. J. Bot. 74(7):1045-1048. [ Links ]
Heiser, C.B. 1989. Artificial hybrids in Solanum sect. Lasiocarpa. Syst. Bot. 14(1):3-6. [ Links ]
Heiser, C.B. 1993. The naranjilla (Solanum quitoense), the cocona (Solanum sessiliflorum) and their hybrid. pp. 29-34. In: Gustafson, J. et al. (eds.). Gene conservation and exploitation. Plenum Press, New York, USA . [ Links ]
Heiser, C.B. and Anderson, 1999. New solanums. p. 379-384. In: Janick, J. (ed.). Perspectives of new crops and new uses. ASHS Press, Alexandria, Virginia, USA . [ Links ]
Heiser, C.B. 1996a. The naranjilla Solanum quitoense and relatives after 38 years. Soc. Econ. Bot. Newsl. Plants and People 14:4-5. [ Links ]
Heiser, C.B. 1996b. Reappraisal of Solanum ferox, S. lasiocarpum, and S.repandum. Solanaceae Newsl. 4(2):44-50. [ Links ]
Hodgkin, T., R. Rana, J. Tuxill, D. Balma, A. Subedi, I. Mar, D. Karamura, R. Valdivia, L. Collado, L. Latournerie, M. Sadiki, M. Sawadogo, A.H.D. Brown and D. I. Jarvis. 2007. Seed systems and crop genetic diversity in agroecosystems. p. 77-116. In: Jarvis, D.I., C. Padoch and H.D. Cooper (eds.). Managing biodiversity in agri-cultural ecosystems. Bioversity International,Columbia University Press, New York, USA . [ Links ]
Isshiki, S. and T. Taura. 2003. Fertility restoration of hybrids between Solanum melongena L. and S. aethiopicum L. Gilo group by chromosome doubling and cytoplasmic effect on pollen fertility. Euphytica 134(2):195–201. [ Links ]
Lobo, M. 1991. Pespectivas de la siembra del lulo o naranjilla (Solanum quitoense Lam.). Bol. Téc. Univ. Nal. de Colombia. Fac. Cien. Agro. Palmira. 2(2):125-130. [ Links ]
Lobo, M. 2000. Papel de la variabilidad genética en el desarrollo de los frutales andinos como alternativa productiva. p. 1-13. En: En: Memorias III Seminario Frutales de Clima Frío Moderado. Centro de Desarrollo Tecnológico de Frutales, CDTF, Manizales, Colombia. [ Links ]
Lobo, M. y Medina, C. I. 2000. Lulo (Solanum quitoense Lam.). En: Caracterização de frutas nativas de América Latina. Serie: Frutas nativas de América Latina. Edição Comemorativa do 30° aniversario da Sociedade Brasileira de Fruticultura. pp. 41-43. [ Links ]
Lobo, M., C. I. Medina, O. Delgado, M. L. Zuluaga, M. Cardona, y A. Osório, 2002. Recursos genéticos de frutales andinos em el sistema de bancos de germoplasma del estado colombiano. pp. 43-48. En: Memorias IV Seminario Frutales de Clima Frío Moderado. Corpoica, UPB, C.D.T.F., Medellín, Colombia. [ Links ]
Lobo, M. 2004. Recursos genéticos de especies frutales. pp. 1-13. En: Memorias VIII Congreso Venezolano de Fruticultura. LUZ, INIA, Corpozulia, UCLA, Maracaibo, Venezuela. [ Links ]
Lobo, M. 2007. Recursos genéticos y mejoramiento de frutales andinos. Visión conceptual. Revista Corpoica (sometido). [ Links ]
Marin, R. y M. Hernández, 1988. Colección, descripción preliminar y montaje de un Banco de Germoplasma de materiales de lulo (Solanum quitoense Lam.) y especies relacionadas. Bogotá. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 249 p. [ Links ]
Medina, C.I. y M. Lobo. 2002. Variabilidad morfológica en el tomate pajarito (Lycopersicon esculentum Var. Cerasiforme), precursor del tomate cultivado. Rev. Corpoica 3(2):39-50. [ Links ]
Medina, C.I. 2003. Estudio de algunos aspectos fisiológicos del lulo (Solanum quitoense Lam.) en el bosque húmedo montano bajo del Oriente Antioqueño. Medellín. Tesis Magister en Ciencias Agrarias, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín. 249 p. [ Links ]
Medina, C.I., E. Martínez, M. Lobo y J.C. López, 2006. Comportamiento bioquímico y del intercambio gaseoso del lulo (Solanum quitoense Lam.) a plena exposición solar en el bosque húmedo montano bajo del Oriente Antioqueño Colombiano. En: Rev. Fac. Nal. Agr. Medellín 59(1): 3123-3146. [ Links ]
Mellas, H. 2000. Morocco . Seed supply systems: Data collection and analysis. p. 155-156. In: Jarvis,
Miflin, B. 2000. Crop improvement in the 21st century. J. Expt. Bot. 51(342):1-8. [ Links ]
Miller, J. S. and P. Diggler. 2003. Diversification of andromonoecy in Solanum section Lasiocarpa; the roles of phenotypic plasticity and architecture. Amer. J. Bot. 90(5):707-715. [ Links ]
National Academy of Sciences. 1975. Underexploited tropical plants with promising economic value. Part IV. Fruits. Naranjilla. NAS, Washington, D.C. USA . 189 p. [ Links ]
Ordás, A., R.A. Malvar and A.M. De Ron. 1994. Relationships among american and spanish populations of maize. Euphytica 79(1-2):149-161. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación. 1996a. Informe sobre el estado de los recursos filogenéticos en el mundo. FAO, Roma, Italia. 75 p. [ Links ]
Organización de las Naciones Unidas para la Agricultura y la Alimentación. 1996 b. Plan de acción mundial para la conservación y utilización sostenible de los recursos filogenéticos para la alimentación y la agricultura. FAO, Roma, Italia. 64 p. [ Links ]
Patiño, V.M. 1962. Edible fruits of Solanum in South American historic and geographic referents. Botl. Museum Lflt. Harvard Univ. 19:215-234. [ Links ]
Patiño, V.M. 2002. Historia y dispersión de los frutales nativos del geotrópico. Centro Internacional de Agricultura Tropical, CIAT, Palmira, Colombia. 655 p. [ Links ]
Plucknett, D.L., N.J.H. Smith, J.T. Williams and N.M. Anishetty.1987. Genebanks and the worlds food. Princeton Univ. Press, NJ, USA . 246 p. [ Links ]
Popenoe, W. 1924. Economic fruit bearing plants of Ecuador . Contributions U.S. Natl. Herbarium 24(5):101-134. [ Links ]
Riday, H., E.C. Brummer, T.A. Cambell and D. Luth. 2003. Comparison of genetic and morphological distance with heterosis between Medicago sativa and subsp. falcata. Euphytica 131(1):37-45. [ Links ]
Ríos, G., M. Romero, M.I. Botero, G. Franco, J.C. Pérez, J.E. Morales, J.L. Gallego y D.I. Echeverri. 2004. Zonificación, caracterización y tipificación de los sistemas de producción de lulo (Solanum quitoense Lam.) en el eje cafetero. Rev. Corpoica. 5(1):22-30. [ Links ]
Rohlf, F.J. 1998. Ntsys. numerical taxonomy and multivariate analysis system. User guide. Exeter software. Department of Ecology and Evolution, State University of New York, Stony, Brook, NY, USA , 31 p. [ Links ]
Roso, C.A., C.I. Medina, and M. Lobo. 2002. Morphologic characterization and agronomic evaluation of a colombian collection of arracacha (Arracacia xanthorrhiza Bancroft). Plant Genet. Resources Nwsl. 132:22-29. [ Links ]
Sahaza, D.P. y M.L. Henao. 2001. Evaluación y caracterización morfológica del lulo (Solanum quitoense Lam.). Trabajo de grado, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín. 63 p. [ Links ]
Schultes, R.E. 1949. Plantae colombianae XII. De plantis principaliter amazoniae colombianae. Botl. Museum Lflt. Harvard Univ. 14(2):21-47. [ Links ]
Schultes, R.E. and J. Cuatrecasas. 1953. Notes on the cultivated lulo. Botl. Museum Lflt. Harvard Univ. 16(5):97-234. [ Links ]
Schultes, R.E. and R. Romero-Castañeda. 1962. Edible fruits of Solanum in Colombia . Botl. Museum Lflt. Harvard Univ. 19(8):235-286. [ Links ]
Sneath, P.H. and R.R. Sokal. 1973. Numerical taxonomy. W. H. Freeman, San Francisco, California, USA. 573 p. [ Links ]
Sokal, R. R. and Michener, C. D. 1958. A statistical method for evaluating systematic relationships. Univ. Kansas Sci. Bul. (38):1409-1438. [ Links ]
Spillane, C. and P. Gepts. 2001. Evolutionary and genetic perspectives on the dynamics of crop genepools. p. 25-71. In: Cooper, H.D., C. Spillane, and T. Hodgkin (eds.). Broadening the genetic base of crop production. CABI Publishing, New York, USA . 480 p. [ Links ]
Torres, A. 2002. Inteligencia de mercados para el lulo, mora y uchuva. p. 326-331. En: Memorias IV Seminario Nacional de Frutales de Clima Frío Moderado. Corpoica, U.P.B., C.D.T.F., Medellín, Colombia. [ Links ]
Tripp, R. 2001. Seed provision and agricultural development. Overseas Dev. Inst. London, 174 p. [ Links ]
Van Beuningen, L.T. and R.H. Busch. 1997. Genetic diversity among North American spring wheat cultivars: Part III. Cluster analysis based on quantitative morphological traits. Crop Sci. 37(3):981-988. [ Links ]
Van Hintum, Th.J.L. 1995. Hierarchical approaches to the analysis of genetic diversity in crop plants. p. 23-34. In: Hodgkin, T. et al. (eds.). Core collections of plant genetic resources. Wiley, New York. [ Links ]
Van Hintum, Th.J.L. and H. Knüpffer.1995. Duplication within and between germplasm collections: Identifying duplication on the basis of passport data. Genet. Resources Crop Evolution. 42(2):127-133. [ Links ]
Vásquez, N., C.I. Medina, y M. Lobo. 2004. Caracterización morfológica de la colección colombiana (Tolima, Huila, Boyacá, Cauca) de arracacha (Arracacia xanthorrhiza). Raíces Andinas: Contribuciones al conocimiento y a la capacitación. Serie: Conservación y uso de la biodiversidad de raíces y tubérculos andinos: Una década de investigación para el desarrollo (1993-2003). (6):165-178. [ Links ]
Whalen, M.D., D.E. Costich and C.B. Heiser. 1981. Taxonomy of Solanum section Lasiocapa. Gentes Herbarum. 12(2):41-129. [ Links ]
Whalen, M.D. and E.E. Caruso. 1983. Phylogeny in Solanum sect. Lasiocarpa (Solanaceae): congruence of morphological and molecular data. Systematic Bot. 8(4):369-380. [ Links ]
Williams, J.T. 1989. Practical considerations relevant to effective evaluation. In: Brown, A.H.D., O.H. Frankel, D.R. Marshall and J.T. Williams (eds.). The use of plant genetic resources. Cambridge University Press, Cambridge, U.K. pp. 235-244. [ Links ]

