










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.60 no.2 Medellín July/Dec. 2007
ANÁLISIS ECOFISIOLÓGICO DEL CULTIVO ASOCIADO MAÍZ (Zea mays L.) – FRÍJOL VOLUBLE (Phaselus vulgaris L.)
ECOPHYSIOLOGICAL ANALYSIS OF CORN (Zea mays L.) – CLIMBIMG COMMON BEAN (Phaselus vulgaris L.) INTERCROPPING.
León Darío Vélez Vargas1, Jairo Clavijo Porras2 y Gustavo Adolfo Ligarreto Moreno3
1 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <ldvelez@unalmed.edu.co>
2 Profesor Especial. Universidad Nacional de Colombia, Sede Bogotá. Facultad de Agronomía. A.A. 14490, Bogotá, Colombia. <jclavijop@unal.edu.co>
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Bogotá. Facultad de Agronomía. A.A. 14490, Bogotá, Colombia. <galigarretom@unal.edu.co>
Recibido: Noviembre 21 de 2006; aceptado: Noviembre 5 de 2007.
RESUMEN
El objetivo es analizar el estado del conocimiento del cultivo asociado maíz-fríjol voluble trepador (MxFv), desde una perspectiva ecofisiológica. Se parte de la revisión de las variables evaluadas en las investigaciones consultadas y su clasificación en variables descriptivas, explicativas y condicionantes. Esta clasificación permitió establecer el carácter predominantemente descriptivo de la mayoría de las investigaciones con respecto a los efectos de la competencia por recursos del suelo y luz sobre las especies asociadas, principalmente en el rendimiento. Solo unas pocas investigaciones abordan el estudio de aspectos morfológicos, de componentes del rendimiento y dinámica del crecimiento y desarrollo de ambas especies, que pueden contribuir a identificar y explicar las causas de los efectos del asocio sobre el comportamiento de ambas especies, como en el caso del rendimiento. Las investigaciones se han concentrado en variables condicionantes como genotipos y densidad de población. En la asociación, el fríjol es la especie más afectada; sin embargo, su alta plasticidad morfofiosiológica posibilita las ventajas de la asociación frente a los unicultivos de maíz y fríjol. La comprensión del funcionamiento de la asociación permitirá avanzar en el mejoramiento de la producción de maíz y fríjol en asocio y de la relaciones de competencia en los agroecosistemas.
Palabras claves: Cultivos asociados, maíz, fríjol, sistemas de producción, ecofisiología, agroecosistemas.
ABSTRACT
This paper reviews the state of knowledge on maize – climbing bean intercropping from an ecophysiological perspective. Starting from the revision of evaluated variables in the consulted investigations and classifing them within three categories: descriptive, explanative and conditional variables. This classification provides elements to argue that researches have been more descriptive than explanative. Many studies describe both association effects and conditionals variables, such as genotypes and population densities, while few studies focus on the mechanisms that explain the effects. The few investigations on mechanisms studies performance of morphological and yield components of both species within the association, and they give few importance to the growing and development. Within the association, bean is the specie most affected; however, it has high morphophysiological plasticity that gives it advantages in the association. The knowledge of these mechanisms may add to understanding of association performance and the selection of genotypes and population densities, and in this way, it may to contribute of the improvement of the intercropping maize-bean.
Key words: Intercropping, maize, climbing common bean, production systems, ecophysiology, agroecosystems.
 INTRODUCCIÓN
INTRODUCCIÓN
EFECTOS Y MECANISMOS
CONCLUSIONES
BIBLIOGRAFIA
Una de las formas en que se cultiva maíz y fríjol voluble trepador, es en asociación (MxFv), y consiste en la siembra simultánea de las dos especies en el mismo sitio, en proporciones de dos a seis plantas de maíz y una a cuatro de fríjol, en donde el maíz sirve de soporte para el crecimiento del fríjol, con arreglos espaciales en cuadro, surcos o triángulo, con distancias entre 1,2 y 0,8 m. La principal característica de la asociación MxFv es que, si bien, los rendimientos de cada especie son menores a los logrados en el correspondiente unicultivo, la combinación de ambos rendimientos es mayor por unidad de área, siendo más eficiente en el uso de recursos económicos, mano de obra, tierra y ambientales (agua, nutrientes y luz). En la asociación los rendimientos del fríjol se reducen en más del 40% y en el maíz entre 10 y 21% (Tabla 1).
Tabla 1. Variables evaluadas para el análisis de las interacciones de competencia del cultivo asociado maíz – fríjol (MxFv) desde la perspectiva ecofisiológica.

El indicador más utilizado para evaluar rendimientos en asociación es el Uso Eficiente o Equivalente de la Tierra (UET) o Rendimiento Relativo Total (RRT). Un UET mayor de 1; por ejemplo, de 1,2, indica que la asociación produce 20% más que el maíz o el fríjol en unicultivo por unidad de área y que entre las especies se presenta algún tipo de interacción que beneficia su cultivo asociado o que hacen demandas diferentes sobre los mismos recursos, disminuyendo la intensidad de competencia, en lo cual la plasticidad del fríjol es fundamental (Gebeyehu, Simane y Kirkby, 2006; Tsubo y Walker, 2004; Abera, Tanaand y Pant, 2005; Mead y Willey, 1980; Sánchez, 1981; Harper, 1977; Davis y García, 1983a). En la asociación MxFv es común UET mayores de 1,2. Ligarreto, Bastidas y Castillo (1992), obtuvieron UET de 1,61 y 1,54 para proporciones de plantas de maíz y fríjol de 4:2 y 4:1, que corresponden a poblaciones de 47 mil plantas de maíz/ha y 23.648 y 11.814 plantas de fríjol/ha, respectivamente. Las variaciones en el valor del índice pueden obedecer a las diferentes poblaciones y genotipos utilizados.
El propósito de esta revisión de literatura sobre el cultivo MxFv, es analizar, desde una perspectiva eco-fisiológica, las relaciones de competencia entre el maíz y el fríjol y los efectos del asocio sobre el microclima. A su vez, los cambios en el microclima tendrán efectos sobre características morfológicas, fisiológicas y en la ontogenia de las especies asociadas. Esta perspectiva de análisis puede contribuir a explicar el comportamiento del maíz y el fríjol en la asociación.
La revisión de literatura se realizó en las bibliotecas de la Universidad Nacional de Colombia - Sedes Bogotá y Medellín, Centro Internacional de Agricultura Tropical (CIAT), Centro de Investigación de Café (CENICAFE), los Centros de Investigación de la Corporación Colombiana de Investigación Agropecuaria (CORPOICA) La Selva, Tulio Ospina y Tibaitatá, y la base de datos CAB International. También se revisó la documentación del Programa de Fríjol del CIAT. La literatura referenciada es aquella relacionada con el cultivo de MxFv, de manera explicita, ya que algunos autores utilizan el término intercultivo sin diferenciar entre cultivos asociados, intercalados o en relevo, y tampoco especifican si el fríjol es voluble o arbustivo.
Se encontraron 21 referencias de investigaciones realizadas en Colombia que evalúan algún aspecto fisiológico o ecofisiológico del cultivo MxFv. En la Tabla 1 se presentan las variables evaluadas en estas investigaciones. Estas variables fueron clasificadas en términos de efectos, mecanismos y variables condicionantes.
Variables evaluadas como efectos, con poca capacidad explicativa. Estas investigaciones son más descriptivas y comprenden la evaluación de los rendimientos, (84,6% de las investigaciones), la eficiencia en el uso de la tierra (UET) (19%), componentes del rendimiento (38,1%), características fenológicas (33,3%) y morfológicas y arquitectura de la planta (28,6%). Las variables componentes del rendimiento y arquitectura de la planta, pueden ser explicativas en la medida que se correlacionen con los rendimientos.
Variables evaluadas como mecanismos o explicativas. Características fisiológicas como materia seca (MS) acumulada (9,5%) en la planta y su distribución entre diferentes órganos captadores de recursos (nutrientes, agua y luz) y de acumulación o receptores de la semilla.
Variables que condicionan la magnitud de los efectos y mecanismos. Las investigaciones para mejorar la producción y el UET, se han enfocado a buscar genotipos (28,6%) y densidades de población adecuadas de cada especie en la asociación (19%).
Esta clasificación coincide con la conclusión de Edje y Laing (1982), acerca de la poca información publicada sobre factores fisiológicos (variables explicativas) que afectan los rendimientos de las leguminosas cuando se asocian con otros cultivos. Keating y Carberry (1993) y Harper (1997), argumentan que las investigaciones sobre esta temática han tratado más con las manifestaciones de la competencia que con la comprensión de las relaciones de causa y procesos que determinan el comportamiento de la asociación. Las variables explicativas constituyen el centro de interés de este artículo.
EFECTOS Y MECANISMOS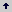
Con base en datos obtenidos por Díaz, Rivera y Quirós (1995), en las Tablas 2 y 3 se presentan los efectos de la asociación sobre rendimientos, duración de las diferentes etapas fenológicas y acumulación y distribución de materia seca (ms) durante la ontogenia de MxFv. A continuación se presenta un análisis de los principales aspectos de estos efectos.
Tabla 2. Efectos del asocio sobre el fríjol (elaborada a partir de Díaz et al., 1995).
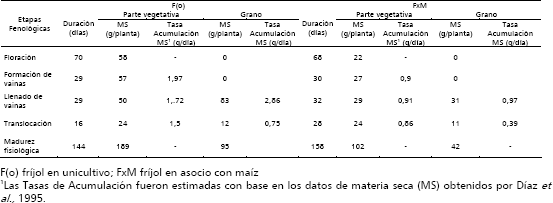
Tabla 3. Efectos del asocio sobre el maíz. (Elaborada a partir de Díaz et al., 1995).
- La duración de la formación de vainas fue similar pero Fx acumuló 30 g menos en la parte vegetativa, con una tasa de crecimiento menor en 1,07 g/día. Esta etapa coincide con la mayor tasa de crecimiento vegetativo del maíz (Tabla 3). La competencia por recursos del suelo y luz puede ser intensa. Se forman las vainas, que es el componente del rendimiento más afectado por la competencia (Tsubo y Walker, 2004; Santa Cecilia y Vieira, 1978; Francis, Prager y Laing, 1978b; Camarena y Cerrate, 1980; Amaya 1980; Davis y García, 1983a; Díaz, Rivera y Ríos 1988; Gebeyehu, Simane y Kirkby, 2006). Al final de esta etapa el tiempo acumulado total es similar, 99 y 98 días para Fo y Fx respectivamente; sin embargo, la ms acumulada total por Fx es 49 g, 66 g menos (57,39%) que Fo que acumuló 115 g.
- El llenado de vainas en Fx duró 3 días más y acumuló 21 g y 52 g menos en la parte vegetativa y en grano, respectivamente, y sus tasas de crecimiento también fueron menores, 0,81 y 1,89 g/día, respectivamente. El llenado de vainas coincide con el periodo de reposo en maíz, cuyas tasas de crecimiento en la parte vegetativa y en grano son las más bajas. La competencia por luz puede ser intensa y baja por recursos del suelo. Al final de esta etapa Fx acumula 78 g en la parte vegetativa y 31 g en grano, 87 g (52,72%) y 52 g (62,65%) menos, respectivamente, que en Fo en tiempos similares, 130 días para Fx y 128 días para Fo.
- La translocación de fotoasimilados duró 12 días más en Fx. La acumulación de MS en la parte vegetativa y grano en Fo y Fx fueron similares y las más bajas de todas las etapas. En Fx, esta etapa coincide con el llenado efectivo y parte de la translocación en maíz. La competencia por luz puede ser intensa.
- La madurez fisiológica se prolonga 14 días más en Fx y su materia seca acumulada total fue 49,3% ( 140 g) menos que en Fo. La reducción de materia seca en la parte vegetativa y grano fueron 87 g (46%) y 53 g (55,79%), respectivamente. No hay una correlación positiva entre duración del periodo y acumulación de materia seca.
- En síntesis, la parte vegetativa en el fríjol en asocio, redujo en todas las etapas fenológicas, la ms acumulada en más del 40%, presentándose la mayor reducción durante la floración (62,1%), y sus tasas de acumulación siempre estuvieron por debajo de 1 g/día. Las tasas de acumulación de Fo en todas las etapas fenológicas fueron superiores a 1 g/día. Esto muestra que los efectos del asocio sobre la parte vegetativa el fríjol se presenta desde las etapas iniciales del crecimiento y desarrollo, posiblemente debidas a la competencia por recursos del suelo. En cuanto a la acumulación de ms en grano del fríjol, la mayor reducción se presentó durante el llenado de la vaina (62,65%) y durante la madurez fisiológica (55,79%).
Efectos del asocio sobre el maíz. La duración de la etapa de floración es similar en maíz en asocio (Mx) y en unicultivo (Mo), esto también fue reportado por Casadiego y García (1982), Davis y García (1983a) y Guzmán (1985); sin embargo, Mx acumuló 12,78% menos de materia seca en la parte vegetativa, posiblemente por competencia por recursos del suelo.
- En Mx el reposo dura 4 días más y acumula 17,39% ( 12 g) de materia seca menos en la parte vegetativa, con tasas de crecimiento menor de 1,96 g, posiblemente por competencia por recursos del suelo. La etapa de reposo coincide con la de mayor acumulación de MS del fríjol. Al final de esta etapa, 126 días después de siembra, Mx acumuló 371 g, 13,52% ( 58 g) menos que Mo que duró 5 días menos.
- El llenado efectivo grano en Mx dura 15 días menos y acumula 33 g y 45 g menos en la parte vegetativa y grano respectivamente; sin embargo, sus tasas de crecimiento fueron mayores en 0,59 y 0,5 g/día, respectivamente. Este periodo coincide con el de translocación en fríjol. No se presenta competencia del fríjol hacia el maíz. El Mx, al final del periodo [153 días después de siembra (dds)], acumuló 430 g en la parte vegetativa y 69 g en grano, 17,47% ( 91 g) y 39,47% ( 45 g) menos, respectivamente, que el Mo que duró 10 días más.
- La translocación en Mx dura 18 días más y acumula 37 g y 25 g más en la parte vegetativa y grano, respectivamente y sus tasas de crecimiento fueron similares. Este periodo coincide, principalmente, con la madurez fisiológica en fríjol. No hay competencia del fríjol hacia el maíz.
- La madurez fisiológica en Mx dura 8 días más y acumuló 9,69% ( 74 g) menos que el Mo. No hay una correlación positiva entre duración del periodo y acumulación de MS. Casadiego y García (1982), Davis y García (1983a) y Guzmán, 1985, encontraron, en otros genotipos de maíz, que la asociación no afectó la duración del ciclo. La reducción en la parte vegetativa y grano fueron, respectivamente, 54 g (9,29%) y 20 g (10,93%) menos que el Mo.
- En síntesis, en el caso del maíz, es el maíz en unicultivo el que presenta la mayor reducción en la acumulación de materia seca en la parte vegetativa durante una etapa fenológica, en la translocación, 61,67%. Sin embargo, en las otras etapas fenológicas la materia seca acumulada en el maíz en asocio, siempre fue menor a la del maíz en unicultivo, presentando la mayor reducción durante el llenado efectivo del grano (35,87%). En el balance total, la materia seca total acumulada por el maíz en asocio en la parte vegetativa, fue menor en 9% con respecto a la del maíz en unicultivo. En cuanto a la acumulación de ms en grano de maíz, el maíz en asocio presenta la mayor reducción durante el llenado efectivo del grano (45%), aunque en la etapa siguiente, translocación, supera al maíz en unicultivo en 25%. En el balance total, la materia seca total acumulada por el maíz en asocio en grano, fue menor en 20% con respecto a la del maíz en unicultivo.
Los mecanismos que expliquen estos efectos pueden ser abordados a partir de las relaciones de competencia entre maíz y el fríjol y de las interacciones del cultivo MxFv con el microclima.
Efectos de la competencia. La reducción de los rendimientos del maíz y del fríjol en el asocio (Mx y Fx respectivamente) han sido atribuidos a los efectos de la competencia por luz y recursos del suelo sobre los componentes del rendimiento y la estructura fotosintética. En Fx, que es el componente de la asociación más afectado, la competencia reduce la altura y el número de nudos y ramas, principalmente después de floración, 24% (27 cm) a los 80 días, y 44% (109 cm) a los 160 días (Angulo, 1986), afectando negativamente el potencial de formación de flores y de vainas (Kohashi-Shiabata, 1990), el área foliar y su distribución en el dosel de la asociación, la habilidad competitiva y los rendimientos (Davis y García, 1983a).Ligarreto, Bastidas y Castillo (1992), encontraron una correlación positiva entre altura del fríjol y densidad de población del maíz. A mayor población de maíz la reducción de la altura del fríjol fue mayor, 40 cm cuando la proporción de maíz: fríjol fue 4:2 y 5 cm cuando la proporción fue 1:1. Angulo (1986) y Murcia y González (1988) encontraron que el número semillas/vaina fue 25% menor en el asocio; en maíz, solo fue afectado el peso de granos. El componente de rendimiento más afectado por variaciones de población es número de vainas/planta. Francis (1976), encontró disminuciones en rendimientos de 78% en la asociación, donde el único componente afectado de manera significativa fue vainas/planta (-58%). Los resultados de Denis (1971), mostraron que de 22 factores del rendimiento estudiados, los tres principales, responsables del 83 y 73% de la variación fueron: peso de las vainas que caracteriza a las plantas con altos rendimientos, número de vainas que caracteriza a las plantas con numerosos entrenudos cortos, y un factor de crecimiento que caracteriza a las plantas con mayor crecimiento vegetativo y rendimientos deficientes.
Dinámica de la competencia por nutrientes, agua y luz. A partir de lo descrito en las Tablas 2 y 3, se puede inferir la dinámica de la competencia y sus efectos diferenciales durante la ontogenia del MxFv. En las etapas iniciales de crecimiento en ambas especies predomina la competencia por recursos del suelo, especialmente por nutrientes. Los siguientes resultados apoyan esta inferencia. En estas etapas los dos cultivos presentan los máximos niveles de extracción de nutrientes (Rajcan y Swanton, 2001; Sanabria, 1975); la competencia por malezas, en maíz y fríjol en unicultivo y asocio, entre los 20 y 60 dds, reduce sus rendimientos en más del 50% (Rajcan y Swanton, 2001; Henao y Mejía, 1981; Sanabria, 1975); la competencia interespecifica maíz-fríjol empieza 20 dds. (Pessanha, 1980) cuando la competencia por luz no es significativa; en relevo (el fríjol se siembra cuando el maíz está en choclo) Davis y García (1983a), no encontraron efecto del sombreado ejercido por el maíz sobre el desarrollo y rendimientos del fríjol; y la competencia por agua es más intensa durante la floración del fríjol, cuando el consumo de agua en la asociación es 30-40% mayor que en los unicultivos (Arango y Vásquez, 1979; Medina, 1977). El estrés por agua durante el crecimiento vegetativo puede limitar la altura, biomasa y tasa de aparición de hojas, lo cual es consistente con los resultados y análisis de las Tablas 2 y 3, pero no necesariamente los rendimientos, estos pueden ser reducidos si el estrés se produce durante la polinización o llenado de la semilla (Rajcan y Swanton, 2001).Con respecto a la competencia por luz, los resultados obtenidos por Tsubo y Walker, 2004, muestran como el sombreado disminuyó el crecimiento del fríjol, la distribución de los foto-asimilados hacía las vainas (40% menos), los rendimientos y la radiación interceptada aunque la eficiencia en su uso fue mayor. Estos autores, encontraron que las hojas de fríjol y maíz en el tercio inferior del dosel del asocio utilizaron aproximadamente 60 y 20%, respectivamente, de la radiación foto-sintéticamente activa y concluyen que el factor agronómico más importante para disminuir la reducción de en la biomasa y en los rendimientos en la asociación es la época de siembra y la distribución espacial de las plantas de ambas especies en el asocio, de tal manera que retrase el inicio de competencia por luz.
Allen et al. (1976), demuestran, para un unicultivo de maíz, que las tasas más bajas de intercambio de CO2, vapor de agua y calor sensible, ocurren en el tercio inferior del dosel del cultivo, en donde el recurso limitante para las fotosíntesis es la radiación fotosintéticamente activa. Esto contribuiría a explicar la significativa reducción en la ms acumulada en Fx, por su menor altura y la disposición de su dosel con respecto al maíz al concentrarlo en los tercios medio e inferior (Figura 1).
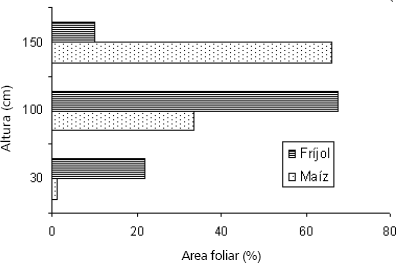
Figura 1. Porcentaje de área foliar ocupando diferentes estratos de maíz y fríjol en intercultivo (adaptado de Vandermmeer, 1989).
Ballesteros (1997a) y (1997b); White e Izquierdo (1989); Kaplan y Koller (1974); Wien et al. (1976); Fukai y Trenbath (1993), relacionan los rendimiento en fríjol más con la duración del llenado de grano que con la tasa de llenado. Las Tablas 2 y 3, muestran como a pesar de una mayor duración de las etapas de llenado de grano en Fx, los rendimientos fueron significativamente menores. Esto permite argumentar que en asociación la principal causa de reducción de biomasa acumulada y rendimientos es la menor radiación fotosintéticamente activa (RFA) que recibe el Fx, a lo cual éste responde con cambios morfológicos-anatómicos (respuestas plásticas o de adaptación modificativas) (Marín, Olivar y Cavanario, 1998; Ballesteros, 1997a y 1997b; Díaz, Rivera y Quirós, 1995 y Díaz, Rivera y Ríos, 1988; Francis, Prager y Laing, 1978b) y bioquímicos (respuestas elásticas o de adaptación modulativa) a nivel de hoja, que reducen la tasa fotosintética y producción de fotoasimilados por hoja. Las investigaciones sobre este tipo de respuestas son escasas, a continuación se presentan los resultados más relevantes en hojas desarrolladas bajo sombra:
- El área foliar aumenta con la menor densidad de flujo radiante, produciendo hojas más delgadas. Esto permite aumentar la eficiencia en captación de luz y en su conversión en fotoasimilados (Lópes, 1988).
- Las hojas son más pequeñas y delgadas, reducen el número de estomas por unidad de área foliar, incrementando la resistencia estomática a la difusión de CO2. Crookstom et al. (1975) estimaron el aumento de esta resistencia en 48%, si el número de estomas disminuía 36%. El tamaño de estomas, células de guarda y apertura estomatal no fue modificado.
- La resistencia del mesófilo a la difusión de CO2 aumentó en 98% y la resistencia total de la hoja para la toma CO2 se incrementó, en promedio, 76% (Crookston et al., 1975).
- La actividad carboxilasa de la Ribulosa Difosfato (RuDP), expresada sobre la base del área foliar, disminuyó 70% y la reducción de la fotosíntesis neta fue del 38% en promedio (Crookston et al., 1975).
- El tamaño de cloroplastos fue menor debido a la menor cantidad de almidón, aunque el número de grana (centros cosechadores de luz) fue mayor, dando una proporción grana/volumen y grana/almidón mayor (Louwerse y Zweerde V.D., 1977).
- Las diferencias en la intensidad de la radiación solar no produjeron cambios importantes en el contenido de clorofila y de enzimas de carboxilación por unidad de área foliar, ni en tamaño de células del mesófilo.
- Estos resultados indican que la fotosíntesis no estuvo limitada por clorofila ni por enzimas de carboxilación, y que es dependiente del número de cloroplastos que se incrementan proporcionalmente en hojas más gruesas (Louwerse y Zweerde V.D., 1977; Sinclair et al., 1977).
Los resultados de Sanabria (1975), sobre producción de biomasa, nutrición mineral y absorción de agua en la asociación maíz – fríjol cultivada en solución nutritiva en invernadero, muestran que la asociación favorece al fríjol, aumentando su producción en biomasa, peso seco de raíces y rendimiento, en comparación con plantas sin asociar. Sin embargo, cuando este experimento se realizó en campo, los efectos sobre estas características fueron similares a los de las otras investigaciones. Estos resultados indicarían que los efectos depresivos de la competencia por nutrientes y agua sobre el fríjol están mediados por el sustrato suelo.
Cambios en el microclima y sus efectos en las especies asociadas. Las Tablas 2 y 3, muestran el efecto de la asociación sobre la duración de las etapas fenológicas en maíz y fríjol. En la asociación la duración del periodo de llenado de grano en maíz, que coincide con el de translocación en fríjol, es reducida en 15 días en tanto que el periodo de translocación en fríjol dura 12 días más. Es decir, durante el mismo momento cronológico, el maíz disminuye el número de días para alcanzar la siguiente etapa, mientras que en fríjol aumentan. En maíz, en la etapa siguiente, translocación, la duración fue mayor. Es decir, para el mismo cultivo, en una etapa se reduce la duración y en la inmediatamente siguiente se alarga. Con base en Allen et al., 1976, es posible plantear, como hipótesis, el siguiente análisis sobre los efectos de la asociación en el microclima, y, a su vez, los efectos del microclima en la ontogenia del fríjol y del maíz.Por su arquitectura y mayor altura, la planta de maíz sombrea al fríjol que concentra su dosel en el tercio medio del maíz (Figura 1). La mayor duración de esta etapa en el fríjol podría atribuirse a la disminución de la incidencia directa de la radiación solar que atenúa la temperatura con relación al unicultivo, reduciendo la acumulación de grados día necesarios para pasar a la etapa siguiente. En las capas bajas de cultivos densos, como en el cultivo MxFv, la concentración de CO2 y de vapor de agua puede ser mayor, en tanto que la temperatura del aire puede disminuir hasta 5ºC (Lemon, 1965, 1967; Allen y Lemos, 1975; Begg et. al., 1964; Lemon et. al., 1967). En un intercultivo sorgo /maní, Keating y Carberry, 1993 encontraron que el sombreado causado por el sorgo redujo la temperatura del dosel del maní.
En cuanto al maíz, su tercio superior está expuesto directamente a la radiación solar, en tanto que su tercio medio experimentaría un incremento en la temperatura, con respecto al maíz en unicultivo, debido al efecto sofocante provocado por la transpiración del follaje del fríjol (Figura 1). De acuerdo con Hallen et al. (1976), el flujo de vapor de agua (calor latente) y de calor sensible es mayor en el tercio medio donde la densidad del área foliar es mayor. Además, la velocidad del viento en el tercio superior del maíz asociado debe ser menor. Estas modificaciones en el microclima permitirían a la planta de maíz acumular más rápidamente los grados día necesarios para pasar a la siguiente etapa.
Así, en fríjol predominaría el efecto refrescante del sombreamiento sobre el sofocante, mientras que en maíz predominaría el efecto sofocante sumado a las condiciones menos refrescantes en su tercio superior. La mayor duración del maíz asociado en la etapa siguiente, translocación, 18 días más, posiblemente se debe a que el fríjol ha llegado a la MF y se encuentra en secado de hojas, su efecto sofocante ya no es tal y la temperatura del microclima disminuye.
Tipos de interacciones. Con base en Harper (1977), es posible describir las interacciones MxFv, en los siguientes términos. El efecto del maíz sobre el fríjol (M/F) es mayor que el efecto del fríjol sobre el fríjol (F/F) y el efecto de fríjol sobre el maíz (F/M) es menor que el efecto del maíz sobre el maíz (M/M), (M/F>F/F y F/M<M/M). Las dos especies hacen demandas similares de los mismos recursos, sin embargo, sus diferencias morfológicas y fisiológicas, les permiten adaptarse, principalmente al fríjol. Este análisis se documenta con los resultados de Ligarreto Bastidas y Castillo (1992) (Figura 2).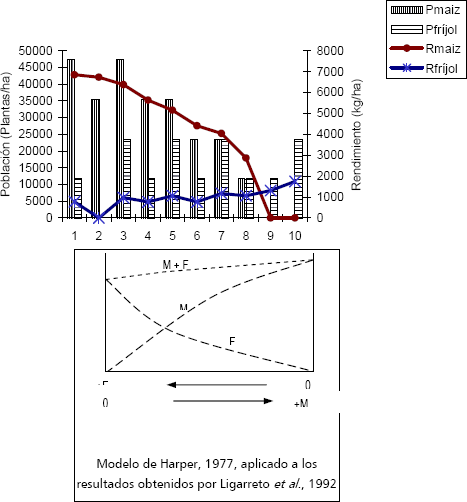
Figura 2. Rendimiento de maíz y fríjol en asociación (elaborado a partir de Ligarreto et al., 1992) (Pmaíz: población maíz; Pfríjol: población fríjol; Rmaíz: rendimiento maíz; Rfríjol: rendimiento fríjol; M maíz; F fríjol).
El efecto del M/F disminuye los rendimientos del F hasta en un 75%, y el efecto del F/M reduce los rendimientos del M hasta en 24%. Con base en los resultados de Ospina (1999) y Chica (2000), es interesante anotar como el incremento en la población de maíz, especialmente por encima de 65 mil plantas/ha, influye significativamente características del maíz, también afectadas en experimentos aditivos por la competencia del fríjol pero no de manera significativa. Estas características son el incremento en el número de días de siembra a floración, la reducción de la prolificidad, del peso y diámetro de las mazorcas, longitud y número de hileras de mazorca (Murcia y González, 1988; Hernández, 1984, Francis et al., 1983 y Ramírez 1980). Oljaca et al. (2000), al evaluar el comportamiento de los componentes del rendimiento de fríjol y maíz, concluyen que en intercalamiento predominó la competencia intraespecífica en maíz (M/M>F/M) y la interespecífica en fríjol (M/F>F/F).
El efecto del maíz sobre el fríjol es mayor que el efecto del fríjol sobre el fríjol y el efecto de fríjol sobre el maíz es mayor que el efecto del maíz sobre el maíz (M/F>F/F y F/M>M/M). En este caso, hay antagonismo y ninguna de las especies contribuye a los rendimientos del asocio en la proporción esperada, cada especie daña el ambiente del otro. Esto puede ocurrir en diseños de reemplazo con una proporción maíz:fríjol de 1:1, o diseños aditivos aumentando la población de fríjol en mayor proporción que la de maíz. En ambos casos hay alto riesgo de volcamiento (Figura 3).
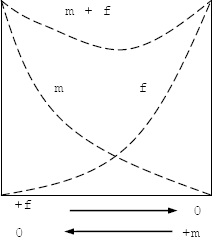
Figura 3. Expresión del comportamiento del maíz y el fríjol en asociación. Los efectos del maíz sobre el fríjol (M/F) son mayores que los efectos del fríjol sobre el fríjol (F/F) y los efectos del fríjol sobre el maíz (F/M) son mayores que los efectos del maíz sobre el maíz (M/M) (Harper, 1977) (m: maíz; f: fríjol).
- Las investigaciones se han orientado a evaluar los efectos de la asociación sobre rendimientos y a la búsqueda de genotipos y densidades de población que optimicen el rendimiento del asocio. Estas investigaciones son más descriptivas que explicativas y correspondes a una escala de trabajo agronómica.
- Es importante a notar que si bien el análisis realizado no consideró la incidencia de los genotipos y/o ambientes en que se realizaron las investigaciones, lo cual no constituía un objetivo de esta revisión, los resultados muestran que los efectos de la asociación son los mismos, sin importar el genotipo y/o el ambiente y que estos inciden en la magnitud de los resultados más no en su tendencia.
- Pocas investigaciones se han ocupado de procesos a escalas morfológicas y fisiológicas como altura de planta, tamaño de hojas, acumulación y partición de fotasimilados. Solo tres investigaciones reportan resultados sobre procesos de apertura y cierre de estomas, intercambio y conductancia de gases a nivel de hoja, cambios en la disposición de cloroplastos y en la concentración y actividad de la clorofila y Rubisco. No se encontraron investigaciones en esta escala para fríjoles volubles comerciales en Colombia y la Región Andina.
- Las investigaciones han tratado más con las manifestaciones de la competencia que con procesos determinantes del comportamiento de la asociación, dado que no se orientan a identificar y explicar los mecanismos responsables; por ejemplo, de la dinámica de la competencia durante la ontogenia de la asociación. Al respecto, es importante establecer aspectos como los siguientes:
¿Es la competencia por los diferentes recursos simultánea o diferencial en el tiempo?
¿Se presentan relaciones de retroacción entre los recursos? Por ejemplo, si la competencia por recursos del suelo afecta la eficiencia fotosintética y con ello la disponibilidad de fotoasimilados para la raíz, afectando la captación de nutrientes.
¿Se presentan procesos de retroacción entre la ontogenia del asocio y su ambiente? Por ejemplo, la competencia inicial por recursos en el suelo, puede ocasionar cambios morfofisiológicos (número de ramas, tamaño y grosor de las hojas de fríjol) que pueden modificar la cantidad y el espectro de luz que penetra en el dosel de la asociación, el déficit de presión de vapor y la temperatura, produciendo cambios morfogénicos y/o morfofisiológicos.
- La competencia por recursos del suelo afecta principalmente la acumulación de ms y no la duración de las etapas. En las primeras etapas de crecimiento, la competencia afecta las estructuras captación de luz (perfil del dosel y anatomía de la hoja), y los componentes del rendimiento (número de vainas). A partir de la formación de vainas, el efecto de competencia es mayor sobre la fotosíntesis. El sombreamiento que ejerce el maíz y la posible competencia por nitrógeno, reduce el tamaño y actividad del aparato fotosintético, disminuyendo tamaño y grosor de hoja, número de estomas, número de cloroplastos y actividad de enzimas, afectando los procesos de intercambio gaseoso y la fijación de CO2.
- El fríjol es la especie más afectada por el asocio. Las respuestas del fríjol se dan principalmente mediante mecanismos de elasticidad y de plasticidad, a través de cambios bioquímicos, moleculares, anatómicos y morfológicos.
- El bajo número de publicaciones encontradas y de temáticas tratadas dan cuenta del estado incipiente del conocimiento sobre la ecofisiología del cultivo MxFv. Además, en los aspectos considerados falta validar los resultados encontrados mediante un mayor número de experimentos, ya que con pocas investigaciones en pocos ambientes (en el espacio y en el tiempo) no es posible proponer como definitivos los resultados y conclusiones obtenidos. Esta situación muestra el enorme potencial por explorar en este cultivo que constituye unos de los renglones productivos de mayor importancia en Colombia, la Región Andina y Centroamérica, dada su contribución a la sostenibilidad económica y ecológica de los campesinos, a la conservación de la agrobiodiversidad y a la soberanía alimentaría de estas regiones. Además, la investigación en estos aspectos toma mayor relevancia debido a la expansión que el cultivo MxFv está teniendo en África (Broughton et al., 2003; Jolliffe y Wanjau, 1999; Vélez, 1999; Fukai, 1993; Trenbath, 1993; Tobón, 1990; Beets, 1982; Sánchez, 1981; Mead y Willey, 1980; Camarena y Cerrate, 1980; Francis, 1978; Bazan et al., 1975; Smale y Jarvis, 2002; Smale, 2002; van Dusen, 2002; Brush, 1999; Morales y Manrique. 2000; Thrupp, 1998; Checa y Blair, 2003, CIAT, 1993a;).
Abera, T., T. Tanaand y L.M. Pant. 2005. Grain yield and LER maize-climbing beam intercropping affected by inorganic, organic fertilizers and population density in western Oromiya, Etiopia. Asian J. Plant Sci. 4(5):458-465. [ Links ]
Allen, L.H., T.R. Sinclair and E.R. Lemon.1976. Radiation and microclimate relationship in multiple cropping systems. p. 171-200. In: Matthias, Stelly (ed.). Multiple cropping. American Society of Agronomy, Wisconsin, United States . 278 p. [ Links ]
Amaya, C. 1980. Efecto de diferentes poblaciones de maíz y de fríjol en el sistema de asociación de maíz x fríjol de enredadera. Tesis Maestría, Facultad de Agronomía, Universidad Nacional de Colombia, Sede Bogotá. 97 p. [ Links ]
Angulo, N. 1986. Comportamiento agronómico de selecciones avanzadas de fríjol voluble con maíces en asocio y en unicultivo. Tesis de Maestría. Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 89 p. [ Links ]
Arango, O. y F. Vásquez. 1979. Evaluación de la competencia agronómica ejercida por el maíz ICA H-210 sobre cuatro materiales de fríjol en asociación. Trabajo de grado, Facultad de Agronomía, Universidad de Caldas, Manizales. 125 p. [ Links ]
Ballesteros, G. 1997a. Análisis del crecimiento en el fríjol común (Phaseolus vulgaris L). Colegio de Posgraduados, Programa de Botánica, Montecillos, México. 71 p. [ Links ]
Ballesteros, G. 1997b. Relaciones fuente demanda en el fríjol común (Phaseolus vulgaris L). Colegio de Posgraduados, Programa de Botánica, Montecillos, México. 71 p. [ Links ]
Bazan, R. A. Pinchinat; G. Paez; N. Mateo; R. Moreno; J. Fargas y W. Forsythe. 1975. Investigación sobre sistemas de producción agrícola para el pequeño agricultor del trópico. Turrialba 25(3):283-293. [ Links ]
Begg, J.E., J.F. Bierhuizen, E.R. Lemon, D.K. Misra, R.O. Slatyer and W.R. Stern. 1964. Dirunal energy budget and water exchanges in bulrush millet in an area of high solar radiation. Agr. Meteorol. 1:294-312. [ Links ]
Beets, W. 1982. Multiple cropping and tropical farming systems. Westview Press, USA . 156 p. [ Links ]
Birol, E. 2002. Applying environmental valuation methods to support the conservation of agricultural biodiversity. In: Smale, M., I. Már and D.I. Jarvis (eds.). 2002. The economics of conserving agricultural biodiversity on farm. pp. 19-29. En: Proc. Wkshp. hosted by the Inst. for Agrobotany (IA), and the International Plant Genetic Resources Institute (IPGRI), Italy 13-16, May 2002. Gödöllo, Hungary. [ Links ]
Broughton, J., G. Hernández, M. Blair, S. Beebe, P. Gepts and J. Vanderleyden. 2003. Beans (Phaseolus spp.) – model food legumes. Plant and Soil 252:55-128. [ Links ]
Brush, S.B. 1999. Genes in the field: on farm conservation of crop diversity. Lewis, Boca Ratón, Fla. 288 p. [ Links ]
Camarena, P. y A. Cerrate. 1980. Comparación de sistemas en monocultivo y asociado y época oportuna de siembra de fríjol en asociación con maíz en el callejón de Haylas, Perú. An. Cient. UNA, Lima, 18(1-4):191-197. [ Links ]
Casadiego, E. y J. García, 1982. Evaluación agronómica de veinticinco variedades de maíz (Zea mays L.) de clima frío bajo el sistema de asociación con fríjol voluble. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 164 p. [ Links ]
CIAT, 1993a. Adaptation of andean climbing beans to warm temperatures. Bean Program, Annu. Rpt. 1993. Working Document 161:85-87. [ Links ]
CIAT, 1993b. Cultivar differences in leaf thickness. Bean Program, Annu. Rpt. 1993. Working Document 161:79-96. [ Links ]
Crookston, R., K. Treharne, P. Ludford and J. Ozbun. 1975. Response of bean to shading. Crop Sci. 15:412-416. [ Links ]
Checa, O. and M.W. Blair. 2003. Trait correlations in climbing beans. Annu. Rpt. Bean Imp. Coop. (BIC) (46):15-16 [ Links ]
Chica, F. 2000. Respuesta del maíz (Zea mays L.). ICA VI57 a cuatro densidades de siembra. Trabajo de grado, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín. 58 p. [ Links ]
Davis, J. and S. Garcia, 1983b. Competitive ability and growth habit of indeterminate beans and maize for intercropping. Field Crops Res. 6:59-75. [ Links ]
Davis, J; Garcia, S. 1983a. Interacción de genotipos por sistema de cultivo en fríjol - maíz. CIAT, Programa de Fríjol. Curso de Fríjol, Cali, Colombia. 8 p. [ Links ]
Denis, L. 1971. Análisis factorial de las variables del tipo de planta, relacionados con el rendimiento del fríjol (Phaseolus vulgaris L.). Tesis de Doctorado, Departament of Crop and Soil Science, Michigan State University. 92 p. [ Links ]
Díaz, C., J. Ríos y J. Rivera, 1983. Evaluación de diferentes arreglos en el sistema de siembra de maíz-fríjol en el oriente antioqueño. Rev. ICA 18(4): 251-259. [ Links ]
Díaz, C., J. Rivera y J. Quirós, 1995. Crecimiento y desarrollo del maíz y fríjol trepador en monocultivo y asociación. Actualidades Corpoica Medellín, Regional 4. (9):5-10. [ Links ]
Díaz, C., J. Rivera y J. Ríos. 1988. Efecto de la competencia en el sistema de siembra maíz/fríjol. En: XIII Reunión de Maiceros de la Zona Andina, Chiclayo, Perú. pp.110-122. [ Links ]
Edje, O.T. and D.R. Laing. 1982. Physiological aspects of maize and bean in monoculture and in association summary intercropping. pp. 69-70. In: Proc. 2nd. Symp. Intercropping in Semi Arid Areas 4-7 Aug. 1980. Morogoro, Tanzania , [ Links ]
Francis, C. 1976. Cultivos asociados. Resumen N. 8c. CIAT, Cali. [ Links ]
Francis, C. 1978. Multiple cropping potentials of beans and maize. Hort. Sci. 13(1):12-17. [ Links ]
Francis, C., M. Prager, G. Tajeda and D. Laing. 1983. Maize genotype by cropping pattern interactions: monoculture vs. intercropping. Crop Sci. 23(2):302-306. [ Links ]
Francis, C., C. Flor and M. Prager. 1978a. Effects of bean on yields and yield components of maize. Crop Sci. 18:760-764. [ Links ]
Francis, C., M. Prager, D. Laing and C.A. Flor, 1978b. Genotype x environment interactions in climbing bean cultivars in monoculture and associated with maize. Crop Sci. 18:242-246. [ Links ]
Fukai, S. 1993. Intercropping, basis of productivity. Fields Crop Res. 34:239-245. [ Links ]
Fukai, S. and B. Trenbath. 1993. Processes determining intercrop productivity and yields of components crops. Fields Crops Res. 34(3-4):247-271. [ Links ]
Gebeyehu, S., B. Simane and R. Kirkby. 2006. Genotype x cropping system interaction in climbing beans (Phaseolus vulgaris L.) grown as sole crop and in association with maize (Zea mays L.). Eur. J. Agron. 24:396–403. [ Links ]
Guzmán, 1985. Efecto de cinco épocas de siembra y dos sistemas de producción en el rendimiento del sistemina maíz x fríjol y en la ocurrencia del disturbio del fríjol. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 40 p. [ Links ]
Harper, J.L. 1977. Population biology of plants. Academic Press, New York. 892 p. [ Links ]
Henao, G. Mejía, F. 1981. Épocas críticas de competencia entre las malezas y el cultivo de fríjol (Phaseolus vulgaris L.) en la zona de Santagueda, Caldas. Trabajo de grado, Facultad de Agronomía, Universidad de Caldas, Manizales. 127 p. [ Links ]
Hernández, C. 1984. Efecto de la densidad de población sobre los rendimientos en la asociación maíz por fríjol. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 74 p. [ Links ]
Jolliffe, P.A. and F.M. Wanjau. 1999. Competition and productivity in crop mixtures: some properties of productive intercrops. J. Agr. Sci. Cambridge 132:425-435. [ Links ]
Kaplan, S.L. and H.R. Koller. 1974. Variation among cultivars in seed growth rate during the linear phase os seed growth. Crop Sci.Madison 14(5):613-614. [ Links ]
Keating, B.A and P.S. Carberry. 1993. Resource capture and use intercropping: solar radiation. Field Crop Res. 34:273-301. [ Links ]
Kohashi-Shiabata, J. 1990. Aspectos de la morfología y fisiología del fríjol (Phaseolus vulgaris L.) y su relación con el rendimiento. Centro de Botánica, Colegio de Postgraduados, Chapingo, Montecillos, México. 44 p. [ Links ]
Lemon, E.R. 1965. Micrometeorology and the physiology of plant in their natural environment. Plant physiology IVA, Academic Press, New York. pp. 203-227. [ Links ]
Ligarreto, G., G. Bastidas y M. Castillo. 1992. Efecto de las densidades de siembra en el rendimiento de la asociación maíz-fríjol voluble. Rev. ICA 27:25-33. [ Links ]
Lopes, Nei Fernandes. 1988. Adaptabilidade fisiológica ao consorcio. p. 375-395. En: Zimmermann, M.J. de O., M. Rocha e T. Yamada (eds.). Cultura do feijoeiro: fatores que afetam a productividade. Associçao Brasileira para Pesquisa da Potasa e do Fosfato, Piracicaba, SP. 589 p. [ Links ]
Louwerse, W. and W. Zweerde V.D. 1977. Photosynthesis, transpiration and leaf morphology of Phaseolus vulgaris and Zea mays grown at different irradiances in artificial and sunlight. Photosyn. 11(1):11-21. [ Links ]
Marin, D., Y. Olivar y R. Cavanario. 1998. Crecimiento y rendimiento en granos en una asociación de maíz (Zea mays L.) y caraota (Phaseolus vulgaris L.), con siembra simultánea. Rev. Fac. Agron. (LUZ) 15:297-311. [ Links ]
Mead, R. and R. Willey. 1980. The concept of a Land Equivalent Ratio and advantages in yield from intercropping. Exp. Agric. 16: 217-228. [ Links ]
Medina, E. 1977. Introducción a la Ecofisiología vegetal. Instituto Venezolano de Investigaciones Científicas, Caracas, Venezuela. 48 p. [ Links ]
Morales, C.F. y A. Manrique. 2000. Agrobiodiversidad en la región andina y amazónica. Tarea Asociación Gráficas Educativas, Lima, Perú. 416 p. [ Links ]
Murcia, S. y J. González, 1988. Evaluación en Semijaca, Cundinamarca, de 6 genotipos de maíz x fríjol voluble en diferentes densidades de población. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 155 p. [ Links ]
Oljaca, S., R. Cvetkovic, D. Kovacevic, G. Vasic and N. Momirovic. 2000. Effect of plant arrangement pattern and irrigation on effciency of maize (Zea mays) and bean (Phaseolus vulgaris) in intercropping system. J. Agr. Sci. 135:261-270. [ Links ]
Ospina, J.G. 1999. Tecnología del cultivo de maíz. Produmedios, Santafé de Bogotá. 335 p. [ Links ]
Pessanha, 1980. Estudios sobre mixturas de cultivares de Feijao (Phaseolus vulgaris L.). Tese Ph.D. Universidad Federal de Viçosa, Minas Gerais. 96 p. [ Links ]
Rajcan, I., and C.J. Swanton. 2001. Understanding maize–weed competition: resource competition, light quality and the whole plant. Field Crops Res. 71(2):139-150. [ Links ]
Rámirez, C. 1980. Efectos de la densidad de siembra sobre la madurez fisiológica y periodo de llenado del grano de poblaciones precoces de maíz. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 104 p. [ Links ]
Rivera, A. 1990. Mejoramiento para los sistemas de cultivo en maíz-fríjol. En: Curso Nacional de Fríjol. 3-7 Dic. 1990. ICA, Rionegro Antioquia, Colombia. 17 p. [ Links ]
Sanabria de M., E. 1975. Producción de biomasa, nutrición mineral y absorción de agua en la asociación maíz-fríjol cultivada en solución nutritiva. Tesis de Maestría, Instituto Interamericano de Ciencias Agrícolas de la OEA, Centro Tropical de Enseñanza e Investigación, Dpto. de Cultivos y Suelos Tropicales, Turrialba, Costa Rica. 80 p. [ Links ]
Sánchez, P. 1981. Suelos del trópico: características y manejo. IICA, San José, Costa Rica, 660 p. [ Links ]
Santa-Cecilia, F. and C. Vieira. 1978. Associated cropping of beans and maize. I. Effects of bean cultivars with different growth habitats. Turrialba 28 (1): 19-23. [ Links ]
Santalla, M., P.A. Casquero and A.M. Ron. 1999. Yield and yield components from intercropping improved bush bean cultivars with maize. J. Agron. Crop Sci. 183(4):263-269. [ Links ]
Serna, G., J. 1997. Efecto de diferentes niveles y épocas de aplicación de nitrógeno en el cultivo asociado maíz-fríjol en el oriente de Cundinamarca. Tesis de Maestría, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 127 p. [ Links ]Sinclair, T.R., J. Goudriaan and De Wit, C.T. 1977. Mesophyll resistence and CO2 compensation concentration in leaf photosynthesis models. Photosynthetica 2(1):56-65.
[ Links ]Skinner, E. 1991. Relaciones alométricas de diez genotipos de maíz asociados y no asociados con fríjol voluble. Trabajo de grado, Facultad de Agronomía, Universidad Nacional de Colombia, Bogotá. 106 p. [ Links ]
Smale, M. 2002. The conceptual framework for economics research in IPGRI´s Global in situ Conservation On-farm Project. p. 4-11. In: Smale, M. I. Már and D.I. Jarvis (eds.). The economics of conserving agricultural biodiversity on farm. En: Proc. Wkshp. hosted by the Inst. for Agrobotany (IA), and the International Plant Genetic Resources Institute (IPGRI), Italy 13-16, May 2002. Gödöllo, Hungary . [ Links ]
Smale, M. and Jarvis, D. 2002. The economics of conserving agricultural biodiversity on farm. Definitions and context. p. 1-3. In: Smale, M. I. Már and D.I. Jarvis (eds.). The economics of conserving agricultural biodiversity on farm. En: Proc. Wkshp. hosted by the Inst. for Agrobotany (IA), and the International Plant Genetic Resources Institute (IPGRI), Italy 13-16, May 2002. Gödöllo, Hungary . [ Links ]
Thrupp, L.A. 1998. Cultivating diversity. Agrobiodiversity and food security. World Resources Institute, Washington, D.C. 80 p. [ Links ]
Tobón, J.I. 1990. La asociación de leguminosas en Colombia. En: Curso Nacional de Fríjol, diciembre 3-7 dic. 1990. ICA, Rionegro, Antioquia. 12 p. [ Links ]
Torregroza, M., C. Ramirez y J. Moreno. 1983. Investigaciones agronómicas en el sistema maíz-fríjol. Part I: Efecto de la productividad y la prolificidad de asociar dialélicamente siete genotipos de maíz con cinco de fríjol voluble. ICA. Subgerencia de Investigación. Tibaitatá. Enero de 1983 (separata). [ Links ]
Trenbath, B.R. 1993. Intercropping for the management of pests and diseases. Field Crop Res. 34:381-405. [ Links ]
Tsubo, M. and S. Walker. 2004. Shade effects on Phaseolus vulgaris L. intercropped with Zea mays L. under well-watered conditions. J. Agron. Crop Sci. 190(3):168-176. [ Links ]
Universidad de Nariño - Facultad de Ciencias Agrícolas. 2003. Comportamiento de líneas mejoradas de fríjol voluble y maíz tipo morocho en asocio directo, en la zona cerealista de Nariño. San Juan de Pasto, Nariño. 28 p. (Mimeografiado). [ Links ]
Van Dusen, M.E. 2002. Identifying the determinants of crops biodiversity on farm with ecomometric applications of the household model. p. 11-19. In: Smale, M.I. Már and D.I. Jarvis (eds.). The economics of conserving agricultural biodiversity on farm. En: Proc. Wkshp. hosted by the Inst. for Agrobotany (IA), and the International Plant Genetic Resources Institute (IPGRI), Italy 13-16, May 2002. Gödöllo, Hungary. [ Links ]
Vélez, L.D. 1999. Dinámica y funcionalidad del cultivo de fríjol (Phaseolus vulagaris L.) a escala predial. pp. 1-6. En: Memorias del Seminario Desarrollo Tecnológicos para el Cultivo de Fríjol. Dpto. de Ciencias Agronómicas, Universidad Nacional de Colombia, – Medellín. EL Peñol, Antioquia. [ Links ]
White, J. y J. Izquierdo. 1989. Fríjol: fisiología del potencial del rendimiento y la tolerancia del estrés. CIAT - FAO. Santiago, Chile . 91 p. [ Links ]
Wien, H.C., S.L. Attschuler, J.L Zbun and D.H. Wallace. 1976. Assimilate distribution in Phaseolus vulgaris L. during the reproductive period. J. Amer. Soc. Hort. Sci. 101(5):510-513. [ Links ]

