










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.62 no.1 Medellín Jan./June 2009
ANTAGONISMO IN VITRO DE Trichoderma harzianum Rifai SOBRE Fusarium oxysporum Schlecht f. sp passiflorae EN MARACUYÁ (Passiflora edulis Sims var. Flavicarpa) DEL MUNICIPIO ZONA BANANERA COLOMBIANA
ANTAGONISM IN VITRO OF Trichoderma harzianum Rifai AGAINST Fusarium oxysporum Schlecht f. sp passiflorae IN PASSION FRUIT (Passiflora edulis Sims var. Flavicarpa) FROM COLOMBIAN BANANERA ZONE MUNICIPALITY
Reinel José Fernández Barbosa1 y Carol Libeth Suárez Meza2
1 Microbiólogo. Universidad Popular del Cesar (UPC). Salida a Patillal, Balneario Hurtado, Valledupar. Colombia. <reifer6@gmail.com>
2 Microbióloga. Universidad Popular del Cesar (UPC). Salida a Patillal, Balneario Hurtado, Valledupar. Colombia. <suarezaser@gmail.com>
Recibido: Enero 21 de 2009; Aceptado: Mayo 12 de 2009.
Resumen. Fusarium oxysporum Schlecht f. sp passiflorae causa la marchitez del maracuyá (Passiflora edulis Sims var. Flavicarpa), afectando su rendimiento. En la búsqueda de alternativas para su control se realizó la presente investigación con el objetivo de determinar la capacidad antagónica de 6 aislamientos de Trichoderma harzianum sobre dicho patógeno. Se evaluaron 3 aislamientos comerciales (TCC-001, TCC-005 y TCC-006) y 3 aislamientos nativos de suelo cultivado con palma de aceite en el Centro de Investigación Caribia de Corpoica (TCN-009, TCN-010, TCN-014). Se hizo la prueba in vitro empleando la técnica de cultivo dual en platos Petri con Agar Sabouraud. Se evaluó competencia por nutrientes y espacio, micoparasitismo y porcentaje de inhibición del crecimiento radial (PICR), por 10 días a 28 ºC. Se estableció un diseño completamente aleatorio, con 13 tratamientos y 3 repeticiones. Todos los aislamientos de T. harzianum superaron en crecimiento a F. oxysporum con radios de de 7,42 cm en cultivo dual. Mientras que el patógeno mostró un radio de 1,99 cm. TCN-009 y TCC-006 expresaron los mejores radios al crecer 4 veces mas rápido que F. oxysporum y reducir 3 veces menos el RCP con respecto al testigo, sin diferencias significativas entre estos tratamientos (P=0,0001); además, produjeron el mayor PICR a los 10 días con valores de 64,61 y 65,91%, respectivamente. No hubo diferencias significativas al comparar los aislamientos comerciales y nativos; sin embargo, por la naturaleza autóctona TCN-009, resulta ser a nivel in vitro, el aislamiento mas promisorio en el biocontrol de F. oxysporum por hallarse en condiciones agroclimáticas similares en la Zona Bananera Colombiana.
Palabras claves: Control biológico, marchitez, inhibición, competencia, micoparasitismo.
Abstract. Fusarium oxysporum Schlecht f. sp passiflorae cause withering of the passion fruit (Passiflora edulis Sims var. Flavicarpa), affecting their performance. In the search of alternatives for its control was carried out the present investigation with the objective of determining the antagonistic capacity of 6 strains of Trichoderma harzianum against the pathogen. 3 commercial strains were evaluated (TCC-001, TCC-005 and TCC-006) and 3 isolated from soil cultivated with palm oil in the Research Center Caribia of Corpoica (TCN-009, TCN-010, TCN-014). These were evaluated under conditions in vitro using the technique of dual cultivation in Petri plates with Agar Sabouraud. Competition for nutritious and space, micoparasitism and Percentage of Radial Growth Inhibition (PRGI) was evaluated, incubating them for 10 days at 28 ºC. A totally randomized design settled down, with 13 treatments and 3 repetitions. All the isolates of T. harzianum exceeded in growth to F. oxysporum with a pathogen growth radiu averages (PGR) of 7.42 cm in dual culture. While the pathogen showed average radiu of 1.99 cm. TCN-009 and TCC-006 expressed the best results in competition for nutrients and space to grow 4 times faster than F. oxysporum and to reduce 3 times less the PRGI compared with the control, without significant differences between treatments (P=0.0001); also, produced the largest PRGI to 10 days with values of 64.61 and 65.91% respectively. There was no significant difference when comparing commercial and native isolates, but for the indigenous nature of the isolation, TCN-009 is, an in vitro level, more promising in the search for an antagonist in the control F. oxysporum in similar agroclimatic regions to the Bananera Zone Colombian.
Key words: Biological control, withering, inhibition, competition, micoparasitism.
El departamento del Magdalena tiene características agroecológicas apropiadas para el cultivo de maracuyá (Passiflora edulis Sims var. Flavicarpa); sin embargo, en el 2006 tuvo una participación en la producción de solo 2,27%, ubicada como una de las zonas con menor aporte, además, con un 0,7% menos que en el 2005 (Corporación Colombia Internacional, 2006). Esta relativa escasez es originada por el desestímulo en el manejo agronómico del cultivo y abandono por problemas fitosanitarios provocados principalmente por alteraciones en frutos y planta por patógenos como Fusarium sp., especialmente en el municipio Zona Bananera, donde el Centro de Investigación Caribia de CORPOICA estableció 20 has de cultivo, de las cuales en el 2006 el 10% fueron totalmente arrasadas por problemas de marchitez, amarillamiento foliar, necrosis foliar, pudrición radical, secamiento total, y posterior desfoliación. Esta enfermedad es conocida como pudrición seca, marchitez, fusariosis o secadera (Espinoza y Mendoza, 2001). La enfermedad tiene como agente etiológico a Fusarium oxysporum Schlecht f. sp passiflorae y se caracteriza por ubicarse en el tejido xilemático de las plantas (Rico et al., 2001). En Colombia existen registros de que esta enfermedad ocasionó, en el norte del Valle del Cauca, entre el 90 y el 100% de muerte a plantas en el periodo de 1997-1999 (Torres et al., 2000).
El riesgo que representa F. oxysporum Schlecht f. sp passiflorae para cultivares susceptibles del maracuyá, ha obligado a utilizar mecanismos de control físicos y químicos, los cuales han alterado negativamente el ecosistema y la calidad del fruto. Esta situación ha conllevado a la búsqueda de nuevas alternativas de control como es el uso de Trichoderma harzianum como biocontrolador (Hermosa et al., 2001). Se han evaluado diferentes concentraciones de Trichoderma sp. contra F. oxysporum, obteniendo que concentraciones de 106 esporas/ml resultan efectivas a nivel de campo (González et al., 2005). Bernal et al. (2000) utilizaron especies de Trichoderma sp. como alternativa ecológica para el control de F. oxysporum Schlecht f. sp cubense, obteniendo antagonismo de parte de los aislamientos con más de un 70%. En Colombia, Trichoderma sp. se ha trabajado en la evaluación de la capacidad antagónica contra hongos fitopatógenos agentes causales de pudriciones radicalesy marchitamientos como Rhizoctonia solani, Sarocladium sp. y Sclerotinia sp. en arroz, flores, papa, hortalizas, frutales y fríjol, F. oxysporum en clavel, Botrytis cinerea en flores, Ceratocystis fimbriata en cafeto, Rosellinia bunodes en cacao, Phytophthora cactorum en manzano, Colletotrichum gloeosporioides en tomate de árbol. En la región caribe colombiana se han establecido estrategias que incorporan a Trichoderma sp. en los procesos de manejo integrado de enfermedades. En este sentido, Páez y Baquero (2004) recomiendan el uso de Trichoderma koningii para el manejo de Phythophthora parasitica Dastur en cítricos.
El objetivo de la investigación fue determinar y comparar la capacidad antagónica in vitro de aislamientos nativos y comerciales de T. harzianum Rifai contra F. oxysporum Schlecht f. sp passiflorae causante de la marchitez de plántulas de maracuyá en el municipio Zona Bananera.
MATERIALES Y MÉTODOS
El estudio se realizó en el Laboratorio de Microbiología y Fitopatología del Centro de Investigación Caribia de CORPOICA, ubicado en la Zona Bananera, distrito Sevilla (Magdalena), con una altitud de 20 msnm, localizado en un ecosistema de bosque seco tropical, con temperatura promedio de 30,4 ºC, precipitación anual media de 1,300 mm y humedad relativa del 83% (Rodríguez, 1998).
Selección de los aislamientos antagonistas. Como antagonistas, se utilizaron aislamientos comerciales de T. harzianum Rifai TCC-001, TCC-005 y TCC-006 provenientes del Centro de Investigación El Roble S.A. y aislamientos nativos Trichoderma harzianum Rifai TCN-009, TCN-010, TCN-014 obtenidos en el C.I. Caribia, caracterizados, evaluados y estudiados por Suárez et al. (2008) como antagonistas para Fusarium solana.
Selección del patógeno. Fusarium oxysporum (FSM-003) fue aislado de plántulas enfermas del maracuyá existentes en el C.I. Caribia por el protocolo del Fusarium Research Center de la Universidad de Pennsylvania, utilizado por Burges et al. (1994) e identificado por el método usado por Gilchrist et al. (2005) y las claves taxonómicas de Seifert (1996), Arikan et al. (2001). Ambos, patógeno y antagonistas fueron mantenidos por separado en cajas Petri con Agar Papa Dextrosa (PDA).
Control biológico in vitro (Cultivos duales). Las pruebas de enfrentamiento se realizaron empleando la técnica de cultivo dual en platos Petri de 5 cm de radio, conteniendo 20 ml de Agar Sabouraud Dextrosa (ASD) de OXOID® pH 5,5. Se colocó en un extremo de la caja de Petri un disco de agar de 4 mm de diámetro con micelio del patógeno (F. oxysporum) y en el extremo opuesto otro disco de 4 mm con micelio del antagonista (T. harzianum) a 5 cm aproximadamente entre ellos (Howell, 2003); se sembró en cajas separadas, un disco de agar (inóculo) con los hongos de cada antagonista y del patógeno, los cuales correspondieron a los controles; posteriormente, los cultivos se incubaron a 25±1 ºC, 12 h continuas en luz, seguido de 12 h de oscuridad durante 10 días, haciéndose mediciones cada 24 h del crecimiento radial del micelio de la colonia de los hongos.
El antagonismo de T. harzianum se comprobó estudiando las variables: Radio de Crecimiento Antagonista (RCA), Radio de Crecimiento Patógeno (RCP), Micoparasitismo (MICMO) y Porcentaje de Inhibición del Crecimiento Radial (PICR).
La competencia por nutrientes y espacio se valoró comparando la velocidad del crecimiento, para esto, se midieron los radios de crecimiento del patógeno (RCP) y de los antagonistas (RCA) con la ayuda de un calibrador "Pie de rey" según recomienda Suárez et al. (2008). Conjuntamente, se evaluó el porcentaje de inhibición de crecimiento radial (PICR), empleando la fórmula de Ezziyyani et al. (2004), PICR = (R1 - R2)/R1 x 100, donde R1 es el radio del patógeno testigo y R2 es el radio del patógeno en enfrentamiento.
Para indicar el micoparasitismo (MICMO) como posible mecanismo de acción de T. harzianum nativos y comerciales, se realizaron observaciones macroscópicas de los cultivos duales, tomándose como índice de micoparasitismo, la invasión del antagonista sobre la superficie del micelio patógeno, teniéndose en cuenta la escala de la Tabla 1 creada por Elías y Arcos (1984) citada en Ezziyyani et al. (2004). Fue necesario comprobar el micoparasitismo, por medio de microscopía, con un microscopio binocular ZEIZZ con aumento de 100x, donde se debieron analizar las interacciones de las hifas antagonistas con las del patógeno, ya sea por enrollamiento o por penetración.
Tabla 1. Escala creada por Elías y Arcos (1984) citada en Ezziyyani et al. (2004) para evaluación de la capacidad antagónica (micoparasitismo), de acuerdo a la medida de la invasión de la superficie, colonización y esporulación de Trichoderma harzianum sobre Fusarium oxysporum.

Diseño y análisis estadístico. Se evaluaron por triplicado 13 tratamientos (T), de los cuales seis pertenecieron a cultivos duales, 1 al testigo del patógeno, 3 a testigos del antagonista nativo y tres a testigos del antagonista comercial. Establecidos como sigue:
T1 (TCN-009 + FSM-003)
T2 (TCN-010 + FSM-003)
T3 (TCN-014 + FSM-003)
T4 (TCC-001 + FSM-003)
T5 (TCC-005 + FSM-003)
T6 (TCC-006 + FSM-003)
T7 (Testigo FSM-003)
T8 (Testigo TCN-009)
T9 (Testigo TCN-010)
T10 (Testigo TCN-014)
T11 (Testigo TCC-001)
T12 (Testigo TCC-005)
T13 (Testigo TCC006)
Se realizó un análisis de varianza bajo un diseño experimental completamente al azar y para identificar las diferencias significativas se uso la prueba de comparación de promedios Duncan con el programa estadístico Mstat SAS versión 6.12.
RESULTADOS Y DISCUSIÓN
Estudio de la competencia por nutrientes y espacio según la comparación del RCA y el RCP. Al hacer la comparación diaria del radio del crecimiento antagonista (RCA), con el radio de crecimiento patógeno (RCP) de cada cultivo dual, se determinó la competencia por nutrientes y espacio, donde se encontró, que los aislamientos de T. harzianum nativos y comerciales se desarrollaron a una velocidad superior a la de F. oxysporum. Debido a que los aislamientos del antagonista tuvieron un crecimiento promedio de 7,42 cm al décimo día en enfrentamiento, muy similar al promedio de sus testigos (7,55 cm), (Figura 1); mientras que el patógeno mostró un crecimiento limitado en cultivo dual, con un promedio de 1,99 cm al décimo día, significativamente diferente al testigo T7 que creció hasta 4,93 cm de radio (Figura 1).
Figura 1. Crecimiento radial de Fusarium oxysporum frente aislamientos de Trichoderma harzianum comerciales y nativos en cultivos duales, a los 10 días de evaluación.
La eficiencia, en cuanto a capacidad antagónica por competencia de nutrientes y espacio, es similar, ya que no presentaron diferencias significativas entre los RCP y RCA en los tratamientos T1, T3, T4, T5 y T6. No obstante al analizar el comportamiento de la cepa nativa TCN-010 en T2, se aprecia que el antagonista crece en menor proporción y el patógeno se desarrolla en mayor distancia, significativamente diferente a los demás tratamientos, sin embargo, el crecimiento del patógeno tiene diferencia altamente significativo (P=0,0001), comparado con el testigo, lo que sugiere que existe represión del crecimiento por parte de T. harzianum TCN-010, pero en dicho proceso se ve afectado el crecimiento de este antagonista ya que varía significativamente al ser comparado con T9 (Figura 1).
Adicionalmente, al comparar los aislamientos nativos y comerciales, se observo que solo hubo diferencias significativas en los cultivos duales de los tratamientos T2 y T6 (P=0,0001), correspondiente al antagonista nativo TCN-010 y al antagonista comercial TCC-006 respectivamente, (Figura 1). Sin embargo, el tratamiento que mostró el mejor radio de crecimiento fue T1 (TCN-009) con un radio de 7,66 cm. Se recomendaría utilizar a nivel de campo un antagonista nativo como éste, porque es probable que su capacidad de adaptabilidad a las condiciones medioambientales sea mayor que la del microorganismo foráneo.
Lo anterior permite plantear que con los aislamientos nativos de T. harzianum es probable llegar a un eficaz control de F. oxysporum de forma preventiva a nivel de campo, aplicando el antagonista antes y en el momento de la siembra de la planta, estimulando la colonización de la rizosfera por la rapidez de crecimiento del hongo, impidiendo el desarrollo y la llegada del patógeno a la planta, siendo favorable cuando el antagonista se encuentra adaptado a las condiciones ambientales del medio (Bernal et al., 2000), sin embargo, es necesario realizar estudios a nivel de invernadero, no solo para determinar su efectividad in vivo, sino para hallar la concentración de esporas/ml apropiada para la aplicación.
Por otra parte, no se presentó micoparasitismo. Es decir, que los aislamientos nativos y comerciales del antagonista, no lograron crecer sobre la superficie del micelio de F. oxysporum y tampoco se evidenció en las observaciones microscópicas, penetración o enrollamiento de las hifas de T. harzianum sobre las de F. oxysporum, lo que sugiere, que los aislamientos nativos y comerciales no poseen capacidad de micoparasitar a este patógeno. Este comportamiento probablemente ocurrió porque, la quitina de la pared celular de F. oxysporum, esta cubierta por una capa proteica que previene su degradación por quitinasas y β-1-3-glucanasas que produce T. harzianum (Inbar y Chet, 1997), lo cual dificulta el proceso de control; a pesar que F. oxysporum no fue micoparasitado, existen reportes, de que este hongo puede ser atacado por Trichoderma sp. (Zago, 2006). Además, Suárez et al. (2008) registran micoparasitismo de estos mismos aislamientos sobre Fusarium solani.
Inhibición del crecimiento radial de Fusarium oxysporum. La inhibición del crecimiento se observó en todos los tratamientos al cuarto día (Figura 2), aún cuando el antagonista se encontraba distante del patógeno, posiblemente a través de antibiosis, mecanismo característico para estas especies de biocontroladores. La forma en que T. harzianum probablemente inhibe el crecimiento radial, aun estando a distancia del patógeno, esta mediada por diversos mecanismos, destacándose la producción de compuestos inhibitorios al medio; antibiosis por producción de metabolitos volátiles y no volátiles entre los cuales se encuentran, pirones, isocianatos, pépticos, y trichocinas (Howell, 2003); además, la producción de enzimas extracelulares difundibles tales como peptinasas, cutinasas, glucanasas, y quitinasas e inactivación de las misma hacia el patógeno, tal como lo sugieren Durán et al. (2003).
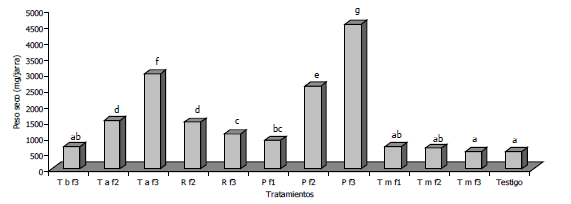
Figura 2. Inhibición del crecimiento radial (PICR) en cultivos duales de Fusarium oxysporum con Trichoderma harzianum aislamientos TCN-009, TCN-010, TCN-014, TCC-001, TCC-005, y TCC-006 durante 10 días de evaluación. Las letras diferentes entre columnas del mismo día indican diferencias significativas para Duncan (P≤0,05)
La Figura 2 describe el comportamiento de los cultivos duales con respecto a la reducción del crecimiento de F. oxysporum, observándose que para el décimo día, la cepa comercial TCC-006 en el tratamiento T6, fue quien exhibió el mayor PICR con un 65,91%, seguido por T1, donde se evaluó la cepa nativa TCN-009, con un 64,61% sin diferencias significativas entré los tratamientos. De acuerdo a los niveles de significación (P=0,0001) entre los resultados obtenidos en la prueba de Duncan, se pudieron categorizar los antagonistas según el grado de inhibición en tres grupos, TCN-009 y TCC-006 en grado alto (60-65%), TCN-014 y TCC-005 en grado medio (57-59%) y TCN-010 con TCC-001 en grado bajo (54 y 56%), todos presentaron un PICR superior al 50%.
Se podría decir que en este tipo de enfermedades el tratamiento es preventivo y no curativo. Por esta razón, resulta más efectivo utilizar un antagonista que crezca primero que el patógeno y que además comience la inhibición de forma temprana, como lo hizo el aislamiento TCN-009.
CONCLUSIÓN
Los aislamientos de T. harzianum utilizados presentaron un modo de acción favorable por competencia de nutrientes y espacio, ya que crecen rápidamente, superando el crecimiento de F. oxysporum, impidiendo el desarrollo normal e inhibiendo en mas del 50% el desarrollo de este, sin diferencias significativas al comparar los aislamientos comerciales y nativos; sin embargo, por la naturaleza autóctona del aislamiento TCN-009 resulta, a nivel in vitro, mas promisorio para la búsqueda de un antagonista en el control de F. oxysporum en regiones agroclimáticas similares a la Zona Bananera del Magdalena.
AGRADECIMIENTOS
Los autores agradecen al C.I. Caribia de CORPOICA y la Compañía Envasadora del Atlántico (CEA) por la financiación de la investigación, a la Universidad Popular del Cesar (UPC) por todo el grupo técnico que desarrollo el análisis de la investigación, y a la C.I. El Roble S.A. por la donación de aislamientos comerciales de Trichoderma harzianum.
BIBLIOGRAFÍA
Arikan, S., M. Lozano, V. Paetznickv and J. Rex. 2001. In vitro susceptibility testing methods for caspofungin against Aspergillus and Fusarium isolates. Antimicrobial Agents and Chemotherapy 45(1): 327-30. [ Links ]
Bernal, A., C.M. Andréu, M.M. Moya, M. González y O. Fernández. 2000. Utilización de Trichoderma spp como alternativa ecológica para el control de Fusarium oxysporum Schlecht f.sp cubense (e.f. Smith) Snyd & Hans. [ Links ]
Burges, B., S. Ummerell, Bullock K., Gott. y Backhouse P. 1994. Laboratory manual for Fusarium research. University of Sydney and Botanic Gardens, Department of Crop Science, Fusarium Research Laboratory, Sydney, Australia. 191 p. [ Links ]
Corporación Colombia Internacional. 2006. Información de monitoreo internacional. Maracuyá. SIM. Inteligencia de mercados [ Links ]
Durán, E., F. Robles, J. Martínez y M. Brito. 2003. Trichoderma Un hongo combatiente de patógenos. Revista Técnico Ambiental Teorema Ambiental 42: 23-26. [ Links ]
Espinoza, L.L. y C. Mendoza. 2001. Etiología de la pudrición de raíz y cuello del chile (Capsicum annuum L.) ocasionado por el hongo Phytophthora capsici en la región de Valseguillo, Puebla, México. Fitopatología 30: 47-55. [ Links ]
Ezziyyani, M., S.C. Pérez, M.E. Requena, L. Rubio y M.E. Candela 2004. Biocontrol por Streptomyces rochei -Ziyani-, de la podredumbre del pimiento (Capsicum annuum L.) causada por Phytophthora capsici. Anales de Biología 26: 69-78. [ Links ]
Gilchrist, L.G., G. Fuentes, C. Martínez, R. López, R.P. Duveiller, M. Singh, I. Henry y A. García. 2005. Guía práctica para la identificación de algunas enfermedades de trigo y cebada. Segunda edición. CIMMYT. México, D. F. [ Links ]
González, J.C., J.M. Maruri y A. González. 2005. Evaluación de diferentes concentraciones de Trichoderma spp., contra Fusarium oxysporum agente causal de la pudrición de plántulas en papaya (Carica papaya L.) en Tuxpan, Veracruz, México. Revista Científica UDO Agrícola. 5 (1): 45-47. [ Links ]
Hermosa, M.R., I. Grondona, J.M. Díaz, E.A. Iturriaga and E. Monte. 2001. Development of a strain-specif SCAR marker for the detection of Trichoderma atroviride 11, a biological control agent against soilborne fungal plant pathogens. Current Genetics 38(6): 343-350 [ Links ]
Howell, C.R. 2003. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Disease 87(1): 4-10. [ Links ]
Inbar, J. and I Chet. 1997. The role of recognition in the induction of specific chitinases of during mycoparasitism by Trichoderma harzianum. Microbiology 141: 2823-2829. [ Links ]
Páez, A.R. y C. Baquero. 2004. Uso de Trichoderma koningii y coberturas muertas para el manejo de la pudrición del pié de los cítricos (Phytophthora parasitica Dastur) en la región Caribe Colombiana En: Memorias. XXV Congreso Nacional ASCOLFI. Cali, Colombia. [ Links ]
Rico, L., B.Z. Guerrero, A. López, C.I. Muñoz, L. Guevara, R.G. Guevara, I. Torres y M.M. González. 2001. Búsqueda de resistencia natural en plantas de chile (Capsicum spp.) contra aislados del complejo fúngico que causa pudrición de raíz. En: Memorias. XXVIII Congreso Nacional de Fitopatología. Querétaro. [ Links ]
Rodríguez, P. 1998. Frutos de la investigación. Corpoica Cinco años. Compendio de productos y procesos de investigación y desarrollo tecnológico. Corporacion Colombiana de Investigacion Agropecuaria-Corpoica, Santa Fe de Bogota, Colombia. 18 p. [ Links ]
Seifert, K. 1996. Fuskey. Fusarium interactive key. Agriculture and agi-food Canada 1-65 [ Links ]
Suárez, C.L., R.J. Fernández, N.O. Valero, R.M. Gomez, A.R. Paez. 2008. Antagonismo in vitro de Trichoderma harzianum Rifai sobre Fusarium solani (Mart.) Sacc., asociado a la marchitez en maracuyá. Revista Colombiana de Biotecnología 10(2): 35-43. [ Links ]
Torres, C., M. Sánchez y L. Gomez 2000. Enfermedades fungosas y bacterianas en el cultivo del maracuyá Passiflora edulis Sims var. Flavicarpa Degener en dos agroecosistemas. Revista de la Asociación Colombiana de la Fitopatología y Ciencias Afines ASCOLFI 24(2): 47-53. [ Links ]
Zago, L. 2006. Dinâmica populacional e ação de Trichoderma sp. no controle de fusariose em mudas de tomateiro e pepineiro. Tese de doutorado. Centro de Ciências Rurais. Universidade Federal de Santa Maria. Santa Maria, RS, Brasil. [ Links ]

