










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.62 no.1 Medellín Jan./June 2009
COMPOSICIÓN Y FACTORES ANTINUTRICIONALES DE LAS SEMILLAS DEL GÉNERO Mucuna
NUTRITIONAL AND ANTINUTRIENTS COMPOSITION OF Mucuna BEANS
Sandra Patricia Chaparro Acuña1; Iván Darío Aristizábal Torres2 y Jesús Humberto Gil González3
1 Químico de Alimentos. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <spchaparroa@unal.edu.co>
2 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <idaristi@unal.edu.co>
3 Profesor Asistente. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <jhgilg@unal.edu.co>
Recibido: Julio 18 de 2008; Aceptado: Abril 1 de 2009
Resumen. Las especies del género Mucuna, leguminosas originarias de la India, han sido ampliamente difundidas en las regiones tropicales y subtropicales del mundo por su facilidad de adaptación. Estas semillas tienen potencial como suplemento proteínico para mejorar la nutrición animal si se disminuyen los factores antinutricionales que contienen, tales como: L-Dopa, fenoles, taninos, ácido fítico e inhibidores de proteasas. Además, estas leguminosas se caracterizan por tener baja cantidad de grasa y alto contenido de fibra, minerales y aminoácidos esenciales. El objetivo de este artículo es revisar los conocimientos actuales de las propiedades nutricionales y los antinutrientes de las especies del género Mucuna.
Palabras claves: Mucuna, antinutrientes, composición química, perfil de aminoácidos, L-Dopa.
Abstract. Mucuna seeds originated from India have been widespread in tropical and subtropical regions due to their easy adaptation. These seeds have potential as a protein supplement to improve ruminant nutrition if the adverse effect of anti-nutritive factors such as L-Dopa, tannins, phenols, phytic acid and protein inhibitors can be diminished. Moreover, these legumes have low fat quantity and high content of fiber, minerals and essential amino acids. The intent of this paper is to review the current knowledge on nutritional and antinutrients composition of Mucuna beans.
Key words: Mucuna beans, antinutrients, chemical composition, amino acid profiles, L-Dopa.
En los países tropicales y del tercer mundo, se ha enfocado un interés arrollador en explorar las proteínas subutilizadas de las leguminosas debido a una escasez aguda de alimentos ricos en proteína, ocasionada por la explosión demográfica y por los altos costos de la proteína animal. Las leguminosas del género Mucuna son de gran interés debido al elevado valor nutricional y su bajo costo. Aunque esta legumbre es subutilizada, tiene una demanda alta como producto vegetal con alto valor nutraceútico en industrias de alimentos y farmaceúticas (Bhat et al., 2007). Por su alto contenido nutricional, las semillas se utilizan en la elaboración de concentrados para la alimentación animal a pequeña escala (Carsky, 2001), y la sustancia química que contiene, la L-Dopa, se usa en medicina para reducir los síntomas de la enfermedad de Parkinson (Amarasekara y Jansz, 1980). El objetivo de este artículo es revisar las características nutricionales, los antinutrientes y los métodos de reducción de éstos en las especies del género Mucuna, para facilitar en un futuro la aplicación como materia prima en la elaboración de alimentos balanceados para animales a escala industrial.
El género Mucuna. Cerca de 100 géneros de Mucuna han sido encontrados en los trópicos y subtrópicos en ambos hemisferios, de los cuales sólo 13 se han caracterizado en Indochina, Malasia y Tailandia. Entre estas se encuentran la Mucuna pruriens, Mucuna nivea (syn. Mucuna lyonii Merr.) (Lyon velvet bean), Mucuna hassjoo (Yokohama velvet bean), Mucuna aterrima Holl. (Mauritius or Bourbon velvet bean), Mucuna utilis Wall. (Bengal velvet bean), and Mucuna deeringiana Merr. (Florida or Georgia velvet bean). (Wilmot-Dear, 1984) (Figura 1). Las semillas del género Mucuna son leguminosas ampliamente distribuidas en Asia, Centroamérica y África, entre los principales países productores se encuentra la India, Nigeria, Guinea y México. Estas variedades son ampliamente utilizadas como alimento, en la dieta de animales y en productos farmacéuticos. Las semillas de este género tienen alto contenido de proteína, carbohidratos y fibra, bajos niveles de lípidos, minerales adecuados y los aminoácidos requeridos. Estas leguminosas poco convencionales son promisorias en cuanto a la nutrición, desarrollo agronómico y en rotación de cultivos. Las plantas tienen gran capacidad para producir follaje de corte o abono verde, eliminar malezas, controlar la erosión y mejorar los suelos (Echeverri y Rodríguez, 1999). El SENA (Servicio Nacional de Aprendizaje) ha implementado esta planta en el mejoramiento de suelos y control de malezas en el Urabá Antioqueño (Colombia) obteniendo excelentes resultados para suelos pobres en nutrientes, ácidos, altos en hierro y aluminio muy pedregosos e invadidos por malezas (AUPEC, 1997). Aunque tiene usos variados, la utilización como materia prima en la elaboración de concentrados para animales puede ser una alternativa de desarrollo económico y social, principalmente en la región del bajo Cauca antioqueño, que presenta erosión debido a la explotación cocalera (Monroy, 2008) y minera (Universidad de Antioquia, 2004; Vicepresidencia de la República de Colombia, 2008).

Figura 1. Semillas del género Mucuna. (Fuente: www.tropicalforages.info mucuna)
Propiedades nutricionales. Las semillas del género Mucuna tienen potencial en la elaboración de alimentos concentrados por su alto contenido de proteína y otros elementos aprovechables (Vadivel y Janardhanan, 2000; Agbede y Aletor, 2005) que hace que se constituya en una alternativa factible para el establecimiento de cultivos comerciales con el propósito de desarrollar productos alimenticios para animales (Trejo, 1998; Echeverri y Rodríguez, 1999; Del Carmen et al., 1999; Navas y Bernal, 1999). Estas leguminosas tienen baja humedad y cenizas, pero alta cantidad de aminoácidos esenciales como isoleucina, leucina, fenilalanina, tirosina y lisina, mientras que los aminoácidos limitantes son metionina y cisteína (Adebowale et al., 2005).
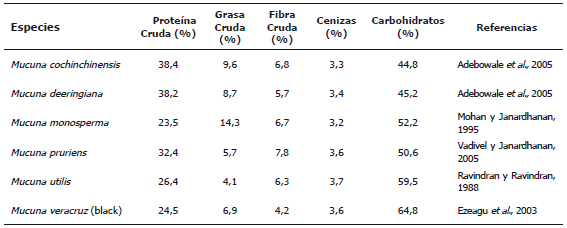
Composición proximal. El contenido de proteína de algunas variedades de Mucuna está entre 23,5-38,4% (Tabla 1). El valor más alto lo presenta la M. cochinchinensis (38,4%), mientras que el resultado más bajo lo tiene la M. monosperma (23,5%). Estos resultados son similares a los encontrados en otras leguminosas como fríjol caupí (24,1%), haba (24,9%), fríjol mungo (24%), lenteja (25,1%) (Narasinga et al., 1989), fríjol guandul (19,2%), guisante (18,2%) y maní (26,7%) (Bressani, 2002), pero más bajos que el observado en la torta de soya (44,3%) (Burcu et al., 2006). De acuerdo con el consumo promedio de proteína, recomendado por el National Research Council (1974), las variedades del género Mucuna pueden contribuir significativamente a la reducción del problema de la desnutrición proteica de los países en vía de desarrollo. Por su parte, la grasa cruda está en un rango de 4,1 a 14,3%, resultados que son similares a los encontrados en otras leguminosas como Canavalia ensiformis (4,3%), C. gladiata (3,3%), Cassia floribunda (3,1%) (Vadivel y Janardhanan, 2005). Estas semillas no se consideran fuente de grasa porque la cantidad de ésta es muy baja comparada con la soya y el maní que tiene altos contenidos de lípidos (19,5 y 25,3%, respectivamente). El contenido de fibra cruda se encuentra entre el 4,2 y el 7,8%, valores más altos que los existentes en otras leguminosas como guisantes y sésamo (Premakumari et al., 1984). La FAO/WHO (1991) ha recomendado un consumo de 22 a 23 g de fibra cruda por cada dieta de 1000 kCal (Kanwar et al., 1997). Por lo tanto, el rango alto de contenido de fibra cruda en las especies de Mucuna es una característica deseable ya que está juega un papel importante en la dieta de humanos y animales. El contenido de cenizas en la Mucuna está entre 3,2-3,7%. Éste es un índice de la calidad de materias primas para concentrados de animales (Pomeranz y Meloan, 1981). La composición de carbohidratos es relativamente alta (44,8-64,8 %) en relación con otras leguminosas como la soya y el maní que tienen alrededor de 20,9 y 26,1%, respectivamente (Narasinga et al., 1989).
Tabla 1. Composición proximal de algunas especies del género Mucuna.
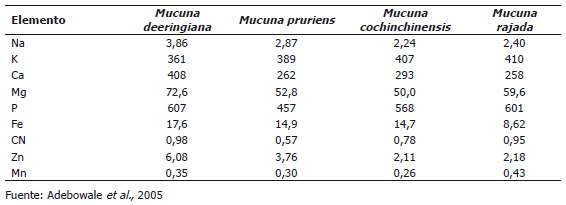
Minerales. Las leguminosas son fuente importante de minerales especialmente potasio, magnesio, hierro, zinc y calcio (Salunkhe et al., 1985). En algunas especies del género Mucuna los principales minerales son potasio (361-410 mg 100 g-1), fósforo (457-607 mg 100 g-1) y calcio (262-408 mg 100 g-1) (Tabla 2), aunque también presentan concentraciones bajas de sodio, hierro, zinc, cobre y manganeso. Estos resultados son más bajos que los encontrados por Vadivel y Janardhanan (2005) en algunas leguminosas, no obstante, estos valores son similares a los de potasio, fósforo y calcio.
Tabla 2. Composición mineral (mg 100 g-1) de algunas especies del género Mucuna.
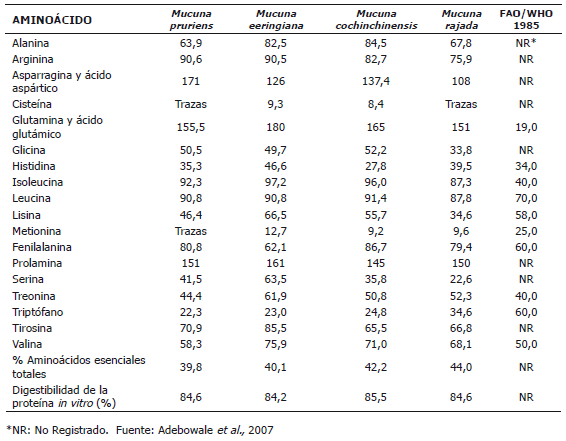
Composición de aminoácidos y digestibilidad de la proteína in vitro. La digestibilidad y la biodisponibilidad de los aminoácidos que constituyen una proteína son factores importantes que caracterizan la calidad de ésta (Hsu et al., 1977; Suman et al., 1992). En su papel de sustancias esenciales de la vida, los aminoácidos son nutrientes críticos en cualquier formulación dietética animal. De aquí radica la importancia de la calidad y cantidad de los aminoácidos presentes en las materias primas utilizadas en la elaboración de alimentos balanceados para animales. En la Tabla 3 se presenta la composición de estos importantes nutrientes en diferentes semillas de Mucuna y los valores de ingesta recomendados por la FAO/WHO (1985). Es importante resaltar que el contenido de aminoácidos de las especies de Mucuna está dentro de los valores de ingesta recomendados por la FAO, y en algunos casos, duplican su valor. Se puede observar que la metionina y la cisteína son los aminoácidos limitantes en la mayoría de las especies de Mucuna. El contenido alto de lisina en la proteína de las especies de Mucuna es un atributo nutricional destacable y probablemente más importante que el contenido de proteína, debido a que hace de esta legumbre una proteína suplementaria significativa para dietas en animales. La digestibilidad de las semillas está entre el 84,2 y 85,5%, representando valores más altos que los observados en Canavalia ensiformis (74,6%), C. gladiata (63,9%), C. obtusifolia (74,6%), Phaseoulus vulgarus, (78,5%), Lentil esculenta, (80,3%) y Cajanus cajan, (59,9%) (Hsu et al., 1977; Oshodi et al., 1995 y Vadivel y Janardhanan, 2005). Sin embargo, los datos obtenidos son más bajos que lo encontrado por Acton et al. (1982) para la caseína (97,7%). Los resultados fueron similares a los observados en los aislados proteicos de otras semillas como fríjol caupi (Sánchez-Vioque et al., 1999) y canola (Gonclaves et al., 1997). Martín-Cabrejas et al., (2008) han encontrado que la germinación de algunas leguminosas no convencionales aumenta la digestibilidad de la proteína y el contenido de aminoácidos, contribuyendo al incremento del valor nutricional de estas semillas.
Tabla 3. Composición de aminoácidos (mg•g-1 proteína cruda) de algunas especies del género Mucuna.
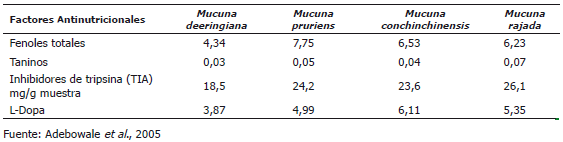
Factores antinutricionales (FAN). Los FAN son sustancias naturales no fibrosas generadas por el metabolismo secundario de las plantas, como un mecanismo de defensa ante el ataque de mohos, bacterias, insectos y pájaros, o en algunos casos, productos del metabolismo de las plantas sometidas a condiciones de estrés, que al estar contenidos en ingredientes utilizados en la alimentación de animales, ejercen efectos contrarios a su óptima nutrición, reduciendo el consumo e impidiendo la digestión, la absorción y la utilización de nutrientes por el animal. Su naturaleza, mecanismos de acción y potencia de sus efectos son muy variados y tienen una amplia distribución en el reino vegetal (Huisman et al., 1990; Liener, 1994). Los componentes antinutricionales más importantes de las especies de Mucuna son la L-Dopa, fenoles, taninos, inhibidores de proteasas, lectinas y ácido fítico (Ravindran y Ravindran, 1988; Siddhuraju et al., 1996; Janardhanan et al., 2003). Adebowale et al., (2005) observaron que la M. deeringiana tiene el contenido más bajo de L-Dopa, fenoles totales, inhibidores de tripsina y saponinas en comparación con otras especies de Mucuna analizadas (Tabla 4).
Tabla 4. Factores antinutricionales de diferentes especies de Mucuna (g•100 g-1 de materia seca)
Dopa. La L-Dopa o (L)-3,4-dihidroxifenilalanina es un aminoácido no proteico precursor del neurotransmisor dopamina. El contenido de dopamina se reduce en el cerebro debido al bloqueo que sufre en su travesía causado por barreras de sangre y que le impiden alcanzar su sitio de acción. La L-Dopa cruza esta barrera y se convierte en dopamina continuando con la neurotransmisión (Kulhalli, 1999). Este compuesto es farmacológicamente activo, por lo cual está siendo usado para tratar los síntomas de la enfermedad de Parkinson (Shaw y Bera, 1993; Prakash y Tewari, 1999). Esta es una enfermedad neurodegenerativa progresiva causada por el desbalance de la dopamina y la acetilcolina en el cerebro (Quinn, 1987). Cuando se usa para tratar esta enfermedad produce alucinaciones serias, discinesia, además de, trastornos gastrointestinales como nauseas, vómito y anorexia (Infante et al., 1990; Buckles, 1995). También ha demostrado ser tóxica para individuos con deficiencia de glucosa-6-fosfato deshidrogenasa en sus eritrocitos, dando como resultado la inducción al favismo con síntomas como dolores de cabeza severos, palpitaciones, confusión y agitación (Kosower y Kosower, 1967). La presencia de L-Dopa fue encontrada inicialmente en Vicia faba (Apaydin et al., 2000) y posteriormente en Phanera Pileostigma, Cassia, Canavalia, Dalbergia, entre otras. Sin embargo, una cantidad más alta de L-Dopa fue observada sólo en especies de fríjol terciopelo (Daxenbichler et al., 1971; Prakash y Tewari, 1999; Marg, 2003). Debido a su estructura química la L-Dopa es oxidable a pH alcalino y a altas temperaturas (70-100 ºC), y en condiciones de suficiente humedad (Siddhuraju y Becker, 2001a). Takasaki y Kawakishi (1997) establecieron que los productos de oxidación de la L-Dopa se conjugan con los enlaces sulfhidrilo (SH) de la cisteína en las proteínas, formando un enlace que conlleva a la polimerización de las proteínas, con la producción de moléculas altamente reactivas que reaccionan con otros componentes del alimento y que producen compuestos coloreados que reducen la calidad del alimento. Este deterioro en el alimento se evita reduciendo el contenido de L-Dopa mediante la combinación de métodos fisicoquímicos (Martín-Cabrejas et al., 1995). Gurumoorthi et al., (2008) aplicaron hidratación, germinación, cocción en microondas y tostado en M. pruriens para la reducción de L-Dopa con buenos resultados. Por su parte, Bhat et al. (2007) observaron que el contenido de L-Dopa en M. pruriens sometida a un tratamiento de radiación g muestra una dependencia con la dosis administrada, presentándose una disminución significativa, cuando la semilla fue sometida a 15 y 30 kGy. También, se ha estudiado el efecto de la hidratación en la reducción de L-Dopa en Mucuna pruriens (Wanjekeche et al., 2003). La hidratación en agua destilada no reduce significativamente el contenido de L-Dopa en M. pruriens, mientras que la hidratación en soluciones alcalinas si la disminuye. Gurumoorthi et al., (2008) observaron que, durante el tostado, este aminoácido se reduce en un 16 a 34%, debido posiblemente a la oxidación parcial o racemización de este compuesto. Estos autores también notaron que la germinación de las semillas durante 120 horas disminuía el contenido de L-Dopa entre 34,8 a 58,4% y sugieren que estos resultados pudieron ser ocasionados por la degradación enzimática y/o cambios estructurales de esta sustancia.
Fenoles. Los fenoles son aquellos productos biosintetizados en las plantas que poseen la cualidad biológica de ser productos secundarios de su metabolismo, y la característica química de contener al menos un grupo hidroxilo unido a un anillo aromático en su estructura molecular. Los compuestos fenólicos son producidos en la planta principalmente para protegerse de situaciones de estrés como la foto oxidación, las heridas, luz ultravioleta (UV), las enfermedades, los patógenos y las plagas (Dixon et al., 1994). Estas sustancias forman complejos con las proteínas y los carbohidratos causando interferencia con la digestibilidad de éstos. También, se ha encontrado que los compuestos fenólicos están relacionados con la dureza de leguminosas (Reyes-Moreno y Paredes-López, 1993; García et al., 1998; Liu, 1995; Martín-Cabrejas et al., 1997; Maurer et al., 2004). De otro lado, la disminución de los fenoles se ha relacionado con su oxidación a altas temperaturas afectando positivamente la permeabilidad de las semillas (Marbach y Mayer, 1974) y permitiendo la reducción de la dureza de éstas (Nassar-Abbas et al., 2008). Algunas especies del género Mucuna tienen una concentración de fenoles totales entre 4,34 y 7,75 g•100 g-1 de materia seca (Adebowale et al., 2005), valores más altos que los observados en Canavalia ensiformis (1,23 %), C. gladiata (1,94%), Cassia floribunda (0,41%) y Vigna radiate (1,45%) (Josephine y Janardhanan, 1992; Vadivel y Janardhanan, 2005). Se ha determinado que los fenoles se concentran principalmente en la cáscara de las leguminosas y por tanto al descascarillarlas se reduce significativamente el contenido de estos antinutrientes (Saharan, 1994). Como los fenoles están en la periferia de la semilla, la hidratación puede también disminuir su concentración (Jood et al., 1987). Siddhuraju et al. (2000) hallaron que un 80% de los fenoles totales pueden ser removidos por el descascarillado o por la hidratación seguida de irradiación. La cocción normal y la cocción a presión que involucran tratamiento térmico en condiciones de alta humedad, pueden destruir los polifenoles (Preet y Punia, 2000). Giami y Okwechime (1993) mencionan una relación inversamente proporcional entre la concentración de fenoles totales y el tiempo de cocción para el caso de la semilla de Vigna unguiculata. Kataria et al. (1989) concluyeron que la germinación redujo el contenido de fenoles en semillas de garbanzo verde (Vigna radiata). Esta disminución puede estar relacionada con la presencia de polifenol oxidasa y la hidrólisis enzimática (Jood et al., 1987).
Taninos. Los taninos son metabolitos secundarios no nitrogenados de las plantas, solubles en agua e insolubles en alcohol y solventes orgánicos (Hagerman y Butler, 1978). En las plantas cumplen funciones de defensa contra los microorganismos, ayudando a prevenir los ataques de hongos y bacterias patógenos. Estos compuestos naturales polifenólicos forman complejos con proteínas, carbohidratos y otros polímeros del alimento. Son sustancias capaces de precipitar alcaloides, gelatinas y otras proteínas en soluciones acuosas (Jansman, 1993). Los taninos tienen efectos nutricionales adversos, pueden inhibir las enzimas digestivas y formar complejos con las membranas mucosas, lo cual resulta en el aumento de pérdidas endógenas y en daños a las mismas (Liener, 1989). Los complejos taninos-proteína son insolubles y esto disminuye la digestibilidad de las proteínas (Carnovale et al., 1991). Por otro lado, se observa que los taninos hidrolizables podrían causar efectos tóxicos a nivel sistémico (Huisman y Tolman, 1992; Jansman, 1993; Butler y Bos, 1993). Las semillas de Mucuna tienen concentraciones de taninos entre 0,03 y 0,07 g•100 g-1 de materia seca. La variación en la concentración de taninos en otras leguminosas ha sido descrita en un rango de 3,0 a 15,6 g/kg (Barampama y Simard, 1994). Otros autores observaron concentraciones inferiores de taninos en algunas leguminosas estudiadas en India (0,04-0,60%) (Vadivel y Janardhanan, 2005). Estos valores son más bajos que los encontrados en fríjol mungo (Rani e Hira, 1998). La germinación reduce el contenido de taninos en las leguminosas lo que se debe, posiblemente, a la formación de una asociación hidrofóbica de los taninos con las proteínas y enzimas de las semillas (Sharma y Sehgal, 1992). Algunos autores indican que el contenido de taninos en las leguminosas se pueden reducir con tratamientos como hidratación, tostado y cocción en autoclave (Vijayakumari et al., 1996; Wanjekeche et al., 2003; Siddhuraju y Becker, 2005).
Inhibidores de tripsina. Los inhibidores de tripsina (TIA) son proteínas que están presentes en cantidades considerables en las leguminosas, y es conocido que afectan la digestibilidad de las proteínas porque reducen la actividad de las proteasas dado que, al formar complejos con ellas, no dejan que estas actúen causando hipertrofia/hiperplasia pancreática y la secreción incrementada de enzimas pancreáticas. Además, producen nódulos acinares y depresión del crecimiento en animales monogástricos (Sánchez et al., 1998; Belmar y Nava, 2000). En el género Mucuna, la concentración de TIA está en un rango de 18,5 a 26,1 g•100 g-1 de materia seca. La pérdida de los inhibidores de tripsina durante la hidratación puede ser causada por el cambio de gradiente de concentración que cambia la tasa de difusión. Generalmente los inhibidores de tripsina son proteínas de bajo peso molecular, por lo tanto, ellas pasan fácilmente de la semilla al medio de hidratación (Bishnoi y Khetarpaul, 1994). Tanto el tratamiento térmico como la cocción normal y la cocción a presión ocasionan una reducción significativa en el contenido de TIA (Siddhuraju y Becker, 2001b; Udedibie y Carlini, 1998); ésta reducción puede ser producida por la naturaleza termolábil de estos antinutrientes (Grewal y Jood, 2006). La germinación causa una reducción significativa en el contenido de TIA, esto puede ser ocasionado por la movilización y rompimiento de los constituyentes químicos incluyendo los inhibidores de tripsina.
CONCLUSIONES
Esta revisión ha permitido reconocer el alto valor nutricional del género Mucuna al ser comparado con otras leguminosas. La composición de aminoácidos de estas semillas es superior a los valores recomendados; y la digestibilidad de la proteína in vitro presenta un nivel mayor comparado otras especies. La presencia de factores antinutricionales no es un problema para el consumo de estas semillas, una vez sean sometidas a un proceso adecuado. En vista de su composición química y nutricional, las semillas de estas leguminosas pueden ser exploradas como una fuente de proteína alternativa que alivie la desnutrición proteico-calórica en niveles bajos de la población y como materia prima en la elaboración de alimentos balanceados para animales.
BIBLIOGRAFÍA
Acton, J.C., L. Breyer and L.D. Satterlee. 1982. Effect of dietary fiber constituents on the in vitro digestibility of casein. Journal of Food Science 47(2): 556-560. [ Links ]
Adebowale, Y.A., A. Adeyemi and A.A. Oshodi. 2005. Variability in the physicochemical, nutritional and antinutritional attributes of six Mucuna species. Food Chemistry 89(1): 37-48. [ Links ]
Adebowale, Y.A., A. Adeyemi, A.A.Oshodi and K. Niranjan. 2007. Isolation, fractionation and characterisation of proteins from Mucuna bean. Food Chemistry 104(1): 287-299. [ Links ]
Agbede, J. and V. Aletor. 2005. Studies of the chemical composition and protein quality evaluation of differently processed Canavalia ensiformis and Mucuna pruriens seed flours. Journal of Food Composition and Analysis 18(1): 89-103. [ Links ]
Amarasekara, A. and R. Jansz. 1980. Studies in Mucuna species of Ski Landa II. Determination of Tetrahydroisoquinoline content of seeds. Journal of National Science Council of Ski Landa 8: 99-103. [ Links ]
Apaydin, H., S. Ertan and S. Ozekmekçi. 2000. Broad bean (Vicia faba) - A natural source of L-DOPA prolongs 'on' periods in patients with Parkinson's disease who have 'on-off' fluctuations. Movement Disorders 15(1): 164 -166. [ Links ]
Agencia Universitaria de Periodismo Científico (AUPEC). 1997. La vitabosa, una aliada de los campos. En: Agencia AUPEC, http://aupec.univalle.edu.co/informes/diciembre97/boletin56/vitabosa.html. 1 p.; consulta: abril 2008. [ Links ]
Barampama, Z. and R.E. Simard. 1994. Oligosaccharides, antinutritional factors and protein digestibility of dry beans as affected by processing. Journal of Food Science, 59(4): 833-838. [ Links ]
Belmar, R. y R. Nava. 2000. Factores antinutricionales en la alimentación de animales monogástricos. En: http://www.sian.info.ve/porcinos/publicaciones/encuentros/viii_encuentro /roberto.htm. Consulta: abril 2008. [ Links ]
Bhat, R., K. Sridhar and K. Tomita-Yokotani. 2007. Effect of ionizing radiation on antinutritional features of velvet bean seeds (Mucuna pruriens). Food Chemistry 103(3): 860-866. [ Links ]
Bishnoi, S. and N. Khetarpaul. 1994. Protein digestability of vegetables and field peas (Pisum sativum). Varietal differences and effect of domestic processing and cooking methods. Plant Foods for Human Nutrition 46: 71-76. [ Links ]
Bressani, R. 2002. Factors influencing nutritive value in food grain legumes: Mucuna compared to other grain legumes. pp. 164-188. In: Florens, M., M. Eilitta, R. Myhrman, L. Carew and R. Carsky (eds.). Food and feed from Mucuna: Current user and the way forward. Proceedings of an International Workshop. CIDICCO, CIEPCA, and World Hunger Research Center. Tegucigalpa, Honduras. [ Links ]
Buckles, D. 1995. Velvetbean: A new plant with a history. Economic Botany 49(1): 13-25. [ Links ]
Burcu, B., A. Eren and E. Putun. 2006. Fast pyrolysis of soybean cake: Product yields and compositions. Bioresource Technology 97(4): 569-576. [ Links ]
Butler, L.G. and K.D. Bos. 1993. Analysis and characterization of tannins in faba beans, cereals and other seeds. A literature review. pp. 1-3. In: Recents advances of research in antinutritional factors in legume seeds: proceedings of de Second International Workshop on 'Antinutritional Factors (ANFs) in Legume Seeds', Wageningen, The Netherlands. [ Links ]
Carnovale, E., E. Lugaro and E. Marconi. 1991. Protein quality and antinutritional factors in wild and cultivated species of Vigna spp. Plant Foods for Human Nutrition 41(1): 11-20. [ Links ]
Carsky, R. 2001. Increasing Mucuna's potential as a food and feed crop. Mucuna News 2. Center for Cover Crops Information and Seed Exchange in Africa (CIEPCA). [ Links ]
Daxenbichler, M.E., C.H. VanEtten, E.A. Hallinan, F.R. Earle and A. Barclay. 1971. Seeds as sources of L-Dopa. Journal of Medicinal Chemistry 14(5):463-465. [ Links ]
Del Carmen, J., A.G. Gernat, R. Myhrman and L.B. Carew. 1999. Evaluation of raw and heated velvet beans (Mucuna pruriens) as feed ingredients for broilers. Poultry Science 78(6): 866-872. [ Links ]
Dixon, R.A., M.J. Harrison and C.J. Lamb. 1994. Early events in the activation of plant defense responses. Annual Reviews of Phytopathology 32: 479-501. [ Links ]
Echeverri, C. y H. Rodríguez. 1999. La vitabosa (Mucuna deeringiana). Servicio Nacional de Aprendizaje SENA. Antioquia. Centro Multisectorial de Oriente. Rionegro. [ Links ]
Ezeagu, I.E., B. Maziya-Dixon and G. Tarawali. 2003. Seed characteristics and nutrient and antinutrient composition of 12 Mucuna accessions from Nigeria. Tropical and Subtropical Agroecosystems 1: 129-140. [ Links ]
FAO/WHO. 1985. Energy and protein requirements. Technical report series No. 724, Geneva. [ Links ]
FAO/WHO. 1991. Protein Quality Evaluation. Report of the joint FAO/WHO expert consultation. FAO Food and Nutrition Paper 51, Food and Agriculture Organization of the United Nations. Rome, Italy. 66 p. [ Links ]
García, E., T.M.C.C. Filisetti, J.E.M. Udaeta and F.M. Lajolo. 1998. Hard-to-cook beans (Phaseolus vulgaris): Involvement of phenolic compounds and pectates. Journal of Agricultural and Food Chemistry 46(6): 2110-2116. [ Links ]
Giami, S.Y. and U.I. Okwechime. 1993. Physicochemical properties and cooking quality of four new cultivars of Nigerian cowpea (Vigna unguiculata L. Walp). Journal of the Science of Food and Agriculture 63(3): 281-286. [ Links ]
Gonclaves, N., J. Vioque, A. Clemente, R. Sanchez-Vioque, J. Bautista y F. Millan. 1997. Obtención y caracterización de aislados proteicos de colza grasos y aceites. Pesquisa Agropecuaria Brasileira 48(5): 282-289. [ Links ]
Grewal, A. and S. Jood. 2006. Effect of processing treatments on nutritional and antinutritional contents of green gram. Journal of Food Biochemistry 30(5): 535-546. [ Links ]
Gurumoorthi, P., K. Janardhanan and R. Myhrman. 2008. Effect of differencial processing methods on L-Dopa and protein quality in velvet bean, an underutilized pulse. LWT-Food Science and Technology 41(4): 588-596. [ Links ]
Hagerman, A.E. and L.G. Butler. 1978. Protein precipitation method for the quantitative determination of tannins. Journal of Agricultural and Food Chemistry 26(4): 809-812. [ Links ]
Hsu, H.W., D.L. Vavak, L.D. Satterlee, and G.A. Miller. 1977. A multienzyme technique for estimating protein digestibility. Journal of Food Science 42(5): 1269-1273. [ Links ]
Huisman, J. and G.H. Tolman. 1992. Antinutritional factors in the plant proteins of diets for non-ruminants. pp. 3-31. In: Garnsworthy, P.C., W. Haresing and D.J.A. Cole (Eds.). Recent advances in animal nutrition. Butterworth Heinemann, U.K. [ Links ]
Huisman, J., A.F. Van der Poel, M.W. Verstegen and E.J. Van Weerden. 1990. Antinutrional factors (ANF) in pig production. World Review of Animal Production 25: 77-82. [ Links ]
Infante, M.E., A.M. Perez, M.R. Simao, F. Manda, E.F. Baquete, A.M. Fernández and J.L. Cliff. 1990. Outbreak of acute toxic psychosis attributed to Mucuna pruriens. The Lancet 336(8723): 1129. [ Links ]
Janardhanan, K., P. Gurumoorthi and M. Pugalenthi. 2003. Nutritional potential of five accessions of a South Indian tribal pulse, Mucuna pruriens var. utilis I. The effect of processing methods on the content of L-Dopa, phytic acid and oligosaccharides. Journal of Tropical and Subtropical Agroecosystems 1: 141-152. [ Links ]
Jansman, A.J.M. 1993. Tannins in feed feedstuffs for simple-stomached animals. Nutrition Research Reviews 6: 209-236. [ Links ]
Jood, S., B.M. Chauhan and A.C. Kapoor. 1987. Polyphenols of chickpea and blackgram as affected by domestic processing and cooking methods. Journal of the Science of Food and Agriculture 39(2): 145-149. [ Links ]
Josephine, R.M. and K. Janardhanan. 1992. Studies on chemical composition and antinutritional factors in three germplasm seed materials of the tribal pulse, Mucuna pruriens (L) DC. Food Chemistry 43(1): 13-18. [ Links ]
Kanwar, K.C., U. Kanwar and S. Shah. 1997. Friendly fibers. Science Republic 34: 9-14. [ Links ]
Kataria, A., B.M. Chauhan and D. Punia. 1989. Antinutrients and protein digestibility (in vitro) of mung bean as affected by domestic processing and cooking, Food Chemistry 52: 9-17. [ Links ]
Kosower, N.S. and E.M. Kosower. 1967. Does 3,4-Dihydroxyphenylalanine play a part in favism?. Nature. 215: 285-286. [ Links ]
Kulhalli, P. 1999. Parkinson's disease therapy - an overview. Heritage Heal July: 29-30. [ Links ]
Liener, I.E. 1994. Implications of antinutritional components in soybeans foods. Critical Review of Food Science and Nutrition 34(1): 31-67. [ Links ]
Liener, I.E. 1989. Antinutritional factors in legume seeds: State of the art. pp. 6-14. In: Huisman, J., F.B. van der Poel, and I.E. Liener (Eds.). Recent advances in research in antinutritional factors in legume seeds. Netherlands, Wageningen. [ Links ]
Liu, K. 1995. Cellular biological and physicochemical basis for the hard to-cook defect in legume seeds. Critical Reviews in Food Science and Nutrition 35(4): 263-298. [ Links ]
Marbach, I. and A.M. Mayer. 1974. Permeability of seed coats to water as related to drying conditions and metabolism of phenolics. Plant Physiology 54: 817-820. [ Links ]
Marg, K. 2003. L-Dopa bearing plants. Natural Product Radiance 2: 126-133. [ Links ]
Martín-Cabrejas, M.A., R.M. Esteban. K. Waldron, G. Maina, G. Grant, S. Bardocz and A. Pusztai. 1995. Hard-to-cook phenomenon in beans: Changes in antinutrient factors and nitrogenous compounds during storage. Journal of the Science of Food and Agriculture 69(4): 429-435. [ Links ]
Martín-Cabrejas, M.A., R.M. Esteban, P. Pérez, G. Maina and K.W. Waldron. 1997. Changes in physicochemical properties of dry beans (Phaseolus vulgaris L.) during long-term storage. Journal of Agricultural and Food Chemisty 45(8): 3223-3227. [ Links ]
Martín-Cabrejas, M.A., M.F. Díaz, Y. Aguilera, V. Benítez, E. Mollá, and R.M. Esteban. 2008. Influence of germination on the soluble carbohydrates and dietary fibre fractions in non-conventional legumes. Food Chemistry 107(3): 1045-1052. [ Links ]
Maurer, G.A., B.F. Ozen, L.J. Mauer and S.S. Nielsen. 2004. Analysis of hard-to-cook red and black common beans using Fourier transform infrared spectroscopy. Journal of Agricultural and Food Chemistry 52(6): 1470-1477. [ Links ]
Mohan, V.R. and K. Janardhanan. 1995. Chemical analysis and nutritional assessment of lesser known pulses of the genus, Mucuna. Food Chemistry 52(3): 275-280. [ Links ]
Monroy, J.C. 2008. El suelo del Bajo Cauca está herido. En: Periódico El Colombiano, http://www.elcolombiano.com.co/BancoConocimiento/E/el_suelo_del_bajo_cauca_esta_herido/el_suelo_del_bajo_cauca_esta_herido.asp. 1 p.; consulta: abril 2008. [ Links ]
Narasinga, B.S., Y.G. Deosthale and K.C. Pant. 1989. Nutritive value of Indian foods. Indian Council of Medical Research, National Institute of Nutrition. Hyderabad, India. [ Links ]
Nasar-Abbas, S.M., J.A. Plummer, H.M. Siddique, P. White, D. Harris, D. and K. Dods. 2007. Cooking quality of faba bean after storage at high temperature and the role of lignins and other phenolics in bean hardening. LWT - Food Science and Technology 47(7): 1260-1267 [ Links ]
National Research Council. 1974. Recommended daily dietary allowance RDA. Nutrition Reviews 31: 373-395. [ Links ]
Navas, G. y J. Bernal. 1999. Caracterización de leguminosas como abono verde para los sistemas de producción del piedemonte llanero y altillanura colombiana. Boletín Técnico No. 16. Corpoica. Villavicencio. [ Links ]
Oshodi, A.A., K.O. Ipinmoroti, E.I. Adeyeye, and G.M. Hall. 1995. Amino and fatty acids composition of African yam bean (Sphenostylis stenocarpa) flour. Food Chemistry 53(1): 1-6. [ Links ]
Pomeranz, Y. and C.E. Meloan. 1981. Food analysis: Theory and practices. C.E. Meloan (Ed.). Publishing Company. AVI, Connecticut. [ Links ]
Prakash, D. and S.K. Tewari. 1999. Variation on L-dopa contents in Mucuna species. Journal of Medicinal and Aromatic Plant Sciences 21: 343-346. [ Links ]
Preet, K. and D. Punia. 2000. Antinutrients and digestibility (in vitro) of soaked, dehulled and germinated cowpea. Nutrition and Health 14(2): 109-117. [ Links ]
Premakumari, M.N., A. Fathima and G. Saraswati. 1984. Dietary fibre content of some food materials. Journal of Food Science and Technology 21: 95-96. [ Links ]
Quinn, N.P. 1987. Levodopa. pp. 317-337. In: W.C. Koller (ed.). Handbook of Parkinson's disease. Marcel Dekker, New York. [ Links ]
Rani, N, and C.K. Hira. 1998. Effect of different treatments on chemical constituents of mash beans (Vigna mungo). Journal of Food Science and Technology 35(6): 540-542. [ Links ]
Ravindran, V. ad G. Ravindran. 1988. Nutritional and antinutritional characteristics of mucuna (Mucuna utilis) bean seeds. Journal of the Science of Food and Agriculture 46(1): 71-79. [ Links ]
Reyes-Moreno, C. and O. Paredes-López. 1993. Hard-to-cook phenomenon in common beans- A review. Critical Reviews in Food Science and Nutrition 33(3): 227-286. [ Links ]
Saharan, K. 1994. Studies on the development of products from ricebean and fababean: Their sensory and nutritional evaluation. pp. 1-275. PhD Thesis. CCS Haryana Agricultural University. Hisar, India [ Links ]
Salunkhe, D.K., S.S. Kadam and J.K. Chavan. 1985. Chemical composition. pp. 29-52. In: Salunkhe, D.K., S.S. Kadam and J.K. Chavan (eds.). Postharvest Biotechnology of Food Legumes. CRC Press Inc., Boca Raton, Florida. 160 p. [ Links ]
Sánchez, A., J.A. Ramírez, O.G. Morales y J.G. Montejano. 1998. Detección de inhibidores de proteasas en extractos de leguminosas y su efecto sobre proteasas endógenas del músculo de pescado. Ciencia y Tecnología Alimentaria 2(1): 12-19. [ Links ]
Sánchez-Vioque, R., A. Clemente, J. Vioque, J. Bautista and F. Millan. 1999. Protein isolates of chickpea (Cicer arietinum L.): chemical composition, functional properties and protein characterisation. Food Chemistry 64(2): 237-243. [ Links ]
Sharma A. and S. Sehgal. 1992. Effect of domestic processing, cooking and germination on the trypsin inhibitor activity and tannin content of faba bean (Vicia faba). Plant Foods for Human Nutrition 42(2): 127-133. [ Links ]
Shaw, B.P. and C. Bera. 1993. A preliminary clinical study to cultivate the effect of Vogorex- SF in sexual disability patients. Indian Journal of Internal Medicine 3: 165-169. [ Links ]
Siddhuraju, P., K. Vijayakumari and K. Janardhanan. 1996. Chemical composition and protein quality of the little known legume Velvet bean (Mucuna pruriens L.DC.). Journal of Agricultural and Food Chemistry 44(9): 2636-2641. [ Links ]
Siddhuraju, P., K. Becker and H.P.S. Makkar. 2000. Studies on the nutritional composition and antinutritional factors of three different germplasm seed materials of an under-utilised tropical legume Mucuna pruriens var. utilis. Journal of Agricultural and Food Chemistry 48(12): 6048-6060. [ Links ]
Siddhuraju, P. and K. Becker. 2001a. Effect of various domestic processing methods on antinutrients and in vitro protein digestibility and starch digestibility of two indigeous varieties of Indian tribal pulse, Mucuna pruriens var utilis. Journal of Agricultural and Food Chemistry 49(6): 3058-3067. [ Links ]
Siddhuraju, P. and K. Becker. 2001b. Rapid reversed-phase high performance liquid chromatographic method for the quantification of L-Dopa (L-3,4-dihydroxyphenylalanine), non-methylated and methylated tetrahydroisoquinoline compounds from Mucuna beans. Food Chemistry 72(3): 389-394. [ Links ]
Siddhuraju, P. and K. Becker. 2005. Nutritional and antinutritional composition, in vitro amino acid availability, starch digestibility and predicted glycemic index of differentially processed mucuna beans (Mucuna pruriens var. utilis): an under-utilised legume. Food Chemistry 91(2): 275-286. [ Links ]
Suman, C.N., P.V. Monterio, G. Ramachandra and L. Sudharshana. 1992. In vitro enzymic hydrolysis of the storage proteins of japanese barnyard millet (Echinochloa frumentacea). Journal of Science of Food and Agriculture 58(4): 505-509. [ Links ]
Takasaki, S. and S. Kawakishi. 1997. Formation of protein-bound 3, 4- dihydroxyphenylanine and 5-S-cysteinyl-3, 4-dihydroxyphenylanine as new cross-linkers in gluten. Journal of Agricultural and Food Chemistry 45(9): 3472-3475. [ Links ]
Trejo, L.W. 1998. Evaluación nutricional del fríjol terciopelo (Stizolobium deeringianum) en la alimentación de pollos de engorde. Tesis M.Sc. Universidad Autónoma de Yucatán. México. 81 p. [ Links ]
Udedibie, A.B and C.R. Carlini. 1998. Brazilian Mucuna pruriens seeds (Velvet Bean) lack hemagglutinating activity. Journal of Agricultural and Food Chemistry 46(4): 1450-1452. [ Links ]
Universidad de Antioquia. 2004. Bajo Cauca. La ruta de los minerales. En: Universidad de Antioquia, http://regionalizacion.udea.edu.co/cauca/sec-region.htm. 2 p.; consulta: abril 2008. [ Links ]
Vadivel, V. and K. Janardhanan. 2000. Nutritional and anti-nutritional composition of velvet bean: an under-utilized food legume in South India. International Journal of Food Sciences and Nutrition 51(4): 279-287. [ Links ]
Vadivel, V. and K. Janardhanan. 2005. Nutritional and antinutritional characteristics of seven south Indian wild legumes. Plant Foods for human Nutrition 60(2): 69-75. [ Links ]
Vicepresidencia de la República de Colombia. 2008. Situación de derechos humanos en el Bajo Cauca Antioqueño. En: Vicepresidencia de la República, http://www.derechoshumanos.gov.co/modules.php?name=informacion&file=article&sid=695. 3 p.; consulta: abril 2008. [ Links ]
Vijayakumari, K., P. Siddhuraju and K. Janardhanan.1996. Effect of different post-harvest treatments on antinutritional factors in seeds of the tribal pulse, Mucuna pruriens (L.) DC. International Journal of Food Science and Nutrition 47(3): 263-272. [ Links ]
Wanjekeche, E., V. Wakasa and J.G. Mureithi. 2003. Effect of germination, alkaline and acid soaking and boiling on the nutritional value of mature and immature mucuna (Mucuna pruriens) beans. Tropical and Subtropical Agroecosystems 1: 183-192. [ Links ]
Wilmot-Dear, C.M. 1984. A revision of mucuna (Leguminosae-Phaseolae) in China and Japan. Kew Bulletin 39(1): 23-65. [ Links ]

