










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.62 no.2 Medellín July/Dec. 2009
IN VITRO EVALUATION OF ANTIMICROBIAL PEPTIDES AGAINST Mycosphaerella fijiensis MORELET AND THEIR INTERACTION WITH SOME CHEMICAL FUNGICIDES
EVALUACIÓN IN VITRO DE PÉPTIDOS ANTIMICROBIANOS CONTRA Mycosphaerella fijiensis MORELET Y SU INTERACCIÓN CON ALGUNOS FUNGICIDAS QUÍMICOS
Luz Estella Vásquez1; Fanny Guzmán2; Manuel Elkin Patarroyo3 and Rafael Arango4
1 Investigadora. Corporación para Investigaciones Biológicas. Unidad de Biotecnología Vegetal. Carrera 72A No. 78B-141. Medellín. Colombia. <lvasquez@cib.org.co>
2 Investigadora. Fundación Instituto de Inmunología de Colombia (FIDIC). Carrera 50 No. 26-00. ZC 020304. Bogota, Colombia. <correo@fidic.org.co>
3 Investigador. Fundación Instituto de Inmunología de Colombia (FIDIC). Carrera 50 No. 26-00. ZC 020304. Bogota, Colombia. <correo@fidic.org.co>
4 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias. Escuela de Biociencias. Calle 59 A No. 63-20. Medellín, Colombia. <rarango@cib.org.co>
Recibido: Abril 2 de 2009; Aceptado: 21 Septiembre de 2009.
Abstract. The interest in the use of antimicrobial peptides to control fungal diseases in banana plants has been rising in the last years. In this paper we analyzed the antifungal activity of different peptides on the in vitro development of Mycosphaerella fijiensis and found that they inhibited fungal mycelial growth and ascospore germination. All peptides evaluated displayed activity against the fungus although with marked differences in IC50 values ranging between 3.06 and 239.61 µg/mL. The peptides exhibiting better activity were the designated peptides Ib-AMP4 and Ib-AMP2, both derived from Impatiens balsamina. Furthermore, the in vitro activity of these peptides seemed compatible with existing chemical fungicides, having no antagonistic or synergistic effect with these agents. The effect of these peptides on M. fijiensis mycelial elongation and ascospores germination suggests that they can be used as potential alternative strategies for the control of black Sigatoka disease.
Key words: Mycosphaerella fijiensis, antimicrobial peptides, black Sigatoka.
Resumen. El interés en el uso de péptidos antimicrobianos para el control de enfermedades de hongos en plantas de banano ha aumentado en los últimos años. En este trabajo se analizó la actividad antifúngica de diferentes péptidos sobre el desarrollo in vitro de Mycosphaerella fijiensis y se encontró que estos inhiben el crecimiento micelial del hongo y la germinación de sus ascosporas. Todos los péptidos evaluados mostraron actividad contra el hongo, aunque con marcadas diferencias en los valores de IC50 los cuales variaron entre 3,06 y 239,61 µg/mL. Los péptidos que presentaron mayor actividad fueron los designados Ib-AMP4 e Ib-AMP2, ambos derivados de la planta Impatiens balsamina. De otro lado, la actividad in vitro de estos péptidos parece ser compatible con algunos fungicidas químicos ya que cuando se usaron en forma combinada, no se encontraron efectos antagónicos o sinérgicos con estos agentes. Los hallazgos sugieren que los péptidos antimicrobianos podrían ser utilizados como posibles estrategias alternativas para el control de la Sigatoka negra.
Palabras claves: Mycosphaerella fijiensis, péptidos antimicrobianos, Sigatoka negra.
An alternative approach for the control of fungal diseases of banana, such as black Sigatoka, the most important disease affecting this crop in the world, is the development of transgenic plants with resistance against Mycosphaerella fijiensis (Chakrabarti et al., 2000). In different plant species the more commonly used genes for this purpose are those coding for hydrolytic enzymes such as chitinases and glucanases (Gómez et al., 2004; Sreeramanan et al., 2006), elicitors of plant defense responses (Strittmatter et al., 1995; Keller et al., 1999) and antimicrobial peptides (AMPs) (Shah, 1997; Cary et al., 2000; Li et al., 2001; Kawata et al., 2003; Osusky et al., 2004; Xing et al., 2006; Sher et al., 2006). AMPs are attractive candidates for transgenic applications for several reasons: their diverse antimicrobial activity, its low toxicity for non-target cells and their probable low cost in terms of energy and biomass involved in their expression (Boman, 1991).
Several AMPs have been extensively studied, including maganins, melittins and plant defensins. Some studies have demonstrated that cecropins and their derivatives (Cary et al., 2000; Rajasekaran et al., 2001) as well as a cecropin-melittin hybrid (Osusky et al., 2000), are capable of inhibiting the in vitro growth of different important plant pathogenic fungi (Osusky et al., 2000).
Banana transgenic lines expressing Dm-AMP1 and Ace-AMP were reported to increase tolerance to Colletotrichum musae under laboratory conditions (Remy, 2000). Chakrabarti et al. (2003) reported the successful expression of a synthetic peptide in banana plants showing increased resistance to both Fusarium oxysporum f.sp. cubense and M. musicola (Chakrabarti et al., 2003).
In view of the great variety of existing antimicrobial peptides and their diverse spectrum of activity, in vitro evaluations of particular peptides to any given pathogen would be important to improve the probability of success and to lower costs in the transgenic approach. Additionally, the in vitro analysis could also allow determination of interactions between peptide mixtures or between peptides and other substances, such as chemical fungicides.
In this work we determined the in vitro activity of six AMPs against M. fijiensis, the causative agent of black Sigatoka, by studying their effect on the development of fungal mycelium and ascospore germination. Moreover, we also evaluated interactions of AMPs with three fungicides: Benomyl, Propiconazol, Azoxystrobin. The data obtained allowed to determine which AMPs appeared to be good candidates to develop transgenic banana plants with possible resistance to the black Sigatoka.
MATERIALS AND METHODS
Fungal isolates and AMPs. Evaluations were carried out with two monosporic strains (codes No. 205001 and 205019) of M. fijiensis previously isolated from infected banana leaves according to standard procedures (Smith et al., 1991a). Strains were kept in PDA (Becton Dickinson and Company, Le Pont de Claix, France) at 25±2 °C. The identity of each strain was confirmed by the morphological characteristics of their conidia and by PCR (Polymerase Chain Reaction), using specific primers (Johanson y Jeger, 1993).
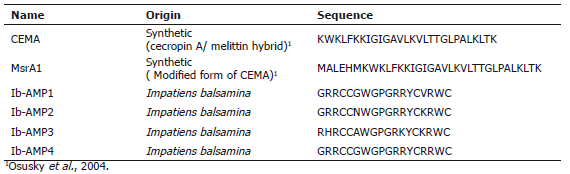
The peptides used, shown in Table 1, were synthesized by the Fundación Instituto de Inmunología de Colombia (FIDIC) (Bogotá, Colombia) by chemical synthesis based on published sequences. Ib-AMPs 1 to 4 are peptides derived from Impatiens balsamina (Tailor et al., 1997). CEMA is composed of 8 amino acid residues derived from melittin at its C-terminal end, and 16 amino acids derived from cecropin at its N-terminus. MrsA1 is the same as CEMA but with an extension of 6 amino acids at the N-terminus (Osusky et al., 2000).
Table 1. Sequence of the evaluated antimicrobial peptides
The peptides were dissolved in deionised water before their use and maintained at 4 °C. Initially, these peptides were assayed at concentrations varying from 1, to 80 µg/mL depending on test results. Concentrations were adjusted by trial and error until reaching the data presented.
Antifungal assays
Activity against mycelial growth. Antifungal tests were done according to Peláez et al. (2006). Briefly, M. fijiensis, mycelium was scraped from agar cultures and re-suspended in 5 mL of distilled water, fragmented with glass beads (Schott, Wertheim, Germany), using a vortex and passed through an etamine cloth (aprox 100 μm pores). Mycelial fragments concentration was measured in a Neubauer chamber (1/10 mm deep, bright line-Boeco, Germany) and adjusted with water to 10 x 104 fragments /mL. The effect of peptides on mycelial growth was evaluated in covered flat bottomed 96-wells microplates (Falcon Becton Dickinson 35-3075 Franklin Lakes, NJ. USA) by adding to each well 30 µL of Sabouraud dextrose broth (Beckton Dickinson and Company, Le Pont de Claix, France); 30 µL of mycelial suspension and 30 µl of the different peptide concentrations. The plate was incubated at room-temperature 25±2 ºC for 8 days and the mycelial growth was measured by the increase in absorbance determined in an ELISA Reader (Biorad model 550, Tokyo, Japan) at 595 nm. Each test was run three times separately, and each concentration was evaluated in triplicate. From these data, the percentage of inhibition for each substance was determined and the 50% Inhibiting Concentration (IC50) calculated considering as 100% the growth in the control wells not containing AMP. Statistical significance of the results was determined by ANAVA with a restrictive random block design. Individual means were compared using the Tukey test with a 95% significance level.
Evaluation of the effect on ascospore germination. A modification of the traditional ascospore germination evaluation method was used (Smith et al., 1991a). Banana leaves at advanced stages of infection with black Sigatoka were set for ascospore discharge over plastic polypropylene circles placed on Petri dishes containing water-granulated agar 2%. Sites of ascospore discharge were located and marked using a light microscope and 20 µL of each peptide solution at a concentration of 40 mg/L for peptides 24892, 24893, 24894, 24895 (over IC75 when evaluated with strain 205001) and 80 mg/l for peptides 24890 y 24891 (highest evaluated concentration) were placed over the ascospores. Plates were then incubated for 48 h at 25±2 °C in a humid chamber and ascospore germination percentage was determined by counting 60 spores per plate under the light microscope at 40X. Ascospore germination was evaluated qualitatively, considering normal germination is bipolar and has no ramifications. Comparisons were made with the growth in a control of ascospores covered only with peptide solvent.
Evaluation of the in vitro interaction between peptides and chemical fungicides. The in vitro interaction between peptides and the chemical fungicides Propiconazole, Benomyl and Azoxystrobin was evaluated using the same method described before for the evaluation of activity against mycelial growth but using combinations of the different compounds at their respective IC25, IC50 and IC75 concentrations. It was, therefore, necessary to determine the inhibitory concentrations for all the chemical fungicides used. The 205019 isolate was considered to be resistant to Benomyl as it grows at concentrations higher than 10 mg/l of this fungicide (Smith et al., 1991b). In order to determine a possible synergistic or antagonistic effect a modified Limpel’s formula Ee=(X+Y)–(XY/100) was used, where Ee is the expected effect from an additive response and X and Y are the relative inhibitory percentages of each compound when applied independently. In this tests, if the combination of the 2 agents produces any value higher than Ee, it is considered that there is a synergistic effect (Richer, 1987).
RESULTS AND DISCUSSION
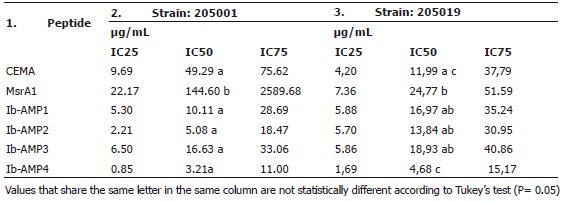
Effect of AMPs over mycelial growth. Mycelial growth inhibition tests revealed that some of the peptides displayed antifungal activity against M. fijiensis at different degrees. The IC50 values of the different peptides on strain 205001 were in a range from 3.06 to 239.61 µg/mL, and for strain 205019 from 5.06 to 19.48 µg/mL. In general, it was observed that the peptides from I. balsamina (Ib-AMP 1 -4) displayed greater activity than CEMA and MrsA1 (Osusky et al., 2004), which had IC50s between 12.60 µg/mL up to 239.61µg/mL.
Additionally, there were significant differences among the activities of the peptides from I. balsamina, the best being Ib-AMP4, with an IC50 value of 3.06 µg/mL for strain 205001 and 5.06 µg/mL for strain 205019 (Table 2), followed by Ib-AMP2(5.084 µg/mL and 13,84 µg/mL) and Ib-AMP1 (10.11 µg/mL and 16,97 µg/mL). This last peptide had was used by Remy (2000) to generate transgenic banana plants but, no activity against M. fijiensis was determined. It has been speculated that the differences in activity observed for individual Ib-AMPs are due to differences in the net charge of the peptides and that a correlation with the charge present in the wall of the target fungus exist. This speculation comes from the observation that a reduction in antifungal activity in media of increased ionic strength is a common feature of most of the small cationic peptides isolated to date, and probably reflects the weakening of electrostatic interactions with the target rather than any alteration of the structure of the peptide by the binding of ions from the medium. This is supported by the fact that the degree of reduction in antifungal activity is dependent on the test fungus (Tailor et al., 1997).
Table 2. Inhibition of mycelial growth of Mycosphaerella fijiensis: inhibitory concentrations achieved by the different peptides.
Considering the interactions related to net charges described above, an evaluation of the antifungal activity of these peptides against M. fijensis isolate 205001 was done in Tris-HCL at pHs 3.5. 4.5, 5.5, 6.5 and 7.5. No statistical differences were found between the different pHs used (data not shown), indicating that antifungal activity is not altered at these pH ranges.
Despite the fact that CEMA has been reported as a good candidate to generate transgenic plants with resistance against fungi, the data obtained in this study showed little in vitro activity against M. fijiensis. Mrsa1 was the peptide with the lower activity against M. fijiensis from all products evaluated. It is possible that other melittin /cecropin hybrids could have had better activity against this fungus since it has been reported that some hybrids can show an increase up to 100 times in their relative inhibitory activity (Osusky et al., 2000).
Effect of AMPs over ascospore germination. Due to limitations in the available amount of peptides and to technical difficulties to evaluate different peptide concentrations, the effect of peptides on ascospore germination was studied only at concentrations of 40 and 80 µg/mL as they matched or exceeded the IC75 calculated on the same data as above. At these concentrations all peptides tested caused abnormal multiple germ tube elongation, while the control which contained only solvent (water) showed the characteristic bipolar germ tube elongation pattern (Figure 1). These results confirm the inhibitory effect of AMP peptides against M. fijiensis and suggest a similar action mechanism despite of the structural differences among the Ib-AMPs and the melittin/cecropin hybrids (CEMA y MrsA1).
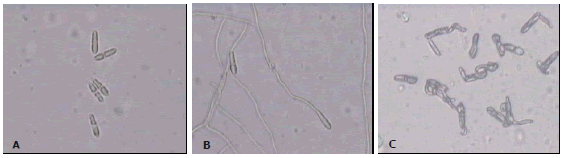
Figure 1. Effect of the peptides over ascospore germination of Mycosphaerella fijiensis. A. Non-germinating ascospores in the presence of peptide Ib-AMP4. B. Ascospores with normal germination (negative control). C. Ascospores with abnormal germination in presence of the chemical fungicide Benomyl (positive control). Ascospores were observed under light microscope at 40X after 48 hours post incubation with the respective treatment.
Cysteine-rich antimicrobial peptides have been previously demonstrated to inhibit specific cellular processes and cause abnormal germ tube formation in several fungi by binding to glucosylceramides, a specific membrane sphingolipid which act as a receptor for the peptides (Lee et al., 1999; Thevissen et al., 2004). It has also been observed that fungal susceptibility might be related to specific sphingolipid mannosyl diinositolphosphoryl ceramide biosynthesis. Yeast mutants revealed that expression of two genes involved on sphingolipids biosynthesis, IPT1 and SKN1, leaded to an increase on susceptibility to γ-thionin DmAMP1 (Thevissen et al., 2005).The disruption of the normal ascospore germination patterns suggests a similar action mechanism of these peptides in M. fijiensis.
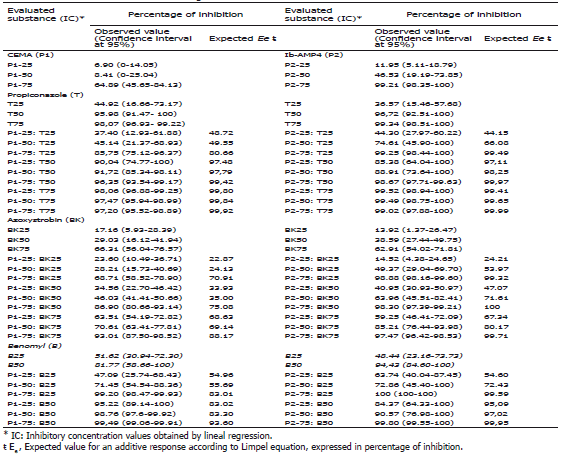
Combinations of AMPs and chemical fungicides. Several combinations of AMPs with the most commonly used chemical fungicides in the control of the disease were evaluated. As seen in Table 3, all the experiments combining AMPs and chemical fungicides showed an additive effect, as expected values (Ee) were similar to the inhibitory values recorded for each mixture. These results rule out a synergic or antagonist effect for all combinations with an exception in the results corresponding to the highest concentration of Propiconazole (IC75) where a small but not significant antagonistic effect with CEMA was observed. The results obtained in these assays suggest that the use of antimicrobial peptides could be compatible with control strategies based on chemical fungicides and that it may be possible to combine the transgenic approach with the use of fungicides.
Table 3. Antimicrobial peptides and agrochemicals: concentrations evaluated to determine the effect of various combinations in the control of black Sigatoka disease.
CONCLUSION
Recently, there has been a lot of interest in the use of antimicrobial peptides to confer resistance to diseases in banana plants (Remy et al., 1998; Gómez et al., 2002; Chakrabarti et al., 2003). In this work we determined the antifungal activity of different peptides on the in vitro development of M. fijiensis. The results presented in this work show that AMPs were capable of inhibiting fungal mycelial growth and ascospore germination. Although all peptides evaluated displayed activity against the fungus, there were differences among them. The peptide with better activity was Ib-AMP4, followed by Ib-AMP2. The effect of antimicrobial peptides on mycelial elongation and ascospore germination, support the notion that these substances may represent potential alternative strategies for the control of black Sigatoka disease.
ACKNOWLEDGEMENTS
This work was supported in part by the Servicio Nacional de Aprendizaje (SENA), the Dirección de Investigaciones Medellín (DIME) de la Universidad Nacional de Colombia, Sede Medellín, the Corporación para Investigaciones Biológicas, CIB and Postgraduate program in Biotechnology of the Universidad Nacional de Colombia, Sede Medellín.
BIBLIOGRAPHY
Boman, H.G. 1991. Antibacterial peptides: key components needed in immunity. Cell 65(2): 205-207. [ Links ]
Cary, J.W., K. Rajasekaran, J.M. Jaynes and T.E. Cleveland. 2000. Transgenic expression of a gene encoding a synthetic antimicrobial peptide results in inhibition of fungal growth in vitro and in planta. Plant Science 154(2): 171-181. [ Links ]
Chakrabarti, A., T.R. Ganapathi, P.K. Mukherjee and V.A. Bapat. 2003. MSI-99, a magainin analogue, imparts enhanced disease resistance in transgenic tobacco and banana. Planta 216: 587-596. [ Links ]
Gómez, M.A., J.A. González, J.L. Ortíz, M.E. Aguilar y J. Sandoval. 2002. Generación de banano (C.V. Gran Nain) transgénico conteniendo genes antifúngicos para conferir resistencia contra Sigatoka Negra. pp. 114-118. En: Memorias. XV Reunión. Asociación de Bananeros de Colombia AUGURA. Cartagena, Colombia. [ Links ]
Gómez, M.A., J.A. González, J.L. Ortíz, J. Sandoval and M.E. Aguilar. 2004. Genetic transformation of cv. ‘Grande naine’. In: Transactions. International Congress on Musa: Harnessing research to improve livelihoods. Penang, Malaysia.
Johanson, A. and M.J. Jeger. 1993. Use of PCR for detection of Mycosphaerella fijiensis and M. musicola, the causal agents of Sigatoka leaf spot in banana and plantain. Mycological Research 97(6): 670-674. [ Links ]
Kawata, M., T. Nakajima, T. Yamamoto, K. Mori, T. Oikawa, F. Fukomoto and S. Kuroda. 2003. Genetic engineering for disease resistance in rice (Oryza sativa L.) using antimicrobial peptides. Japan Agricultural Research Quarterly-JARQ 37(2): 71-76. [ Links ]
Keller, H., N. Pamboukdjian, M. Ponchet, A. Poupet, R. Delon, J-L. Verrier, D. Roby and P. Ricci. 1999. Pathogen-induced elicitin production in transgenic tobacco generates a hypersensitive response and nonspecific disease resistance. Plant Cell 11: 223-236. [ Links ]
Lee, D.G., S.Y. Shin, D.H. Kim, M.Y. Seo, J.H. Kang, Y. Lee, K.L. Kim and K.S. Hahm. 1999. Antifungal mechanism of a cysteine-rich antimicrobial peptide, Ib-AMP1, from Impatiens balsamina against Candida albicans. Biotechnology Letters 21: 1047-1050. [ Links ]
Li, Q., C.B. Lawrence, H.Y. Xing, R.A. Babbitt, W.T. Bass, I.B. Maiti and N.P. Everett. 2001. Enhanced disease resistance conferred by expression of an antimicrobial magainin analog in transgenic tobacco. Planta 212: 635-639. [ Links ]
Osusky, M., G. Zhou, L. Osuska, R.E. Hancock, W.W. Kay and S. Misra. 2000. Transgenic plants expressing cationic peptide chimeras exhibit broad-spectrum resistance to phytopathogens. Nature Biotechnology 18: 1162-1166. [ Links ]
Osusky, M., L. Osuska, R.E. Hancock, W.W. Kay and S. Misra. 2004. Transgenic potatoes expressing a novel cationic peptide are resistant to late blight and pink rot. Transgenic Research 13(2): 181-190. [ Links ]
Peláez, J.E., L.E. Vásquez, T.J. Díaz, D.A. Castañeda, E. Rodríguez and R.E. Arango. 2006. Use of a microtiter well dilution assay to measure activity of antifungal compounds against Mycosphaerella fijiensis Morelet. Revista Facultad Nacional de Agronomía, Medellín 59(2): 3425- 3433. [ Links ]
Rajasekaran, K., K.D. Stromberg, J.W. Cary and T.E. Cleveland. 2001. Broad-spectrum antimicrobial activity in vitro of the synthetic peptide D4E1. Journal of Agricultural and Food Chemistry 49(6): 2799-2803. [ Links ]
Remy, S., A. Buyens, B.P.A. Cammue, R. Swennen and L. Sági. 1998. Production of transgenic banana plants expressing antifungal proteins. Acta Horticulturae 490: 433-436. [ Links ]
Remy, S. 2000. Genetic transformation of banana (Musa spp.) for disease resistance by expression of antimicrobial proteins. Ph.D. thesis No. 420, Katholieke Universiteit Leuven, Belgium. 341 p. [ Links ]
Richer, D.L. 1987. Synergism: a patent view. Pesticide Science 19(4): 309-315. [ Links ]
Sher, R., M. Nishihara, S. Yamamura, I. Nakamura and M. Mii. 2006. Transgenic potatoes expressing wasabi defensin peptide confer partial resistance to gray mold (Botrytis cinerea). Plant Biotechnol 23: 179-183 [ Links ]
Smith, C.M., L.E.B Johnson and C.A Shillingford. 1991. In vitro methods for monitoring flusilazole sensitivity of Mycosphaerella fijiensis (black Sigatoka of bananas). EPPO Bulletin 21(2): 332-335. [ Links ]
Smith, C.M., A.E. Trivellas, L.E.B. Johnson and M.M. Joshi. 1991. Methods for monitoring the sensivity of a range of fungal pathogens to benzimidazole fungicides. In: FRAC methods for monitoring fungicide resistance. Bulletin EPPO 21: 336-341. [ Links ]
Sreeramanan, S., M. Maziah, N.M. Rosli, M. Sarria and R. Xavier. 2006. Particle bombardment mediated co-transformation of chitinase and β-1, 3 glucanase genes in banana. Biotechnol 5(2): 203-216.
Shah, D.M. 1997. Genetic engineering for fungal and bacterial diseases. Current Opinion in Biotechnology 8(2): 208-214. [ Links ]
Strittmatter, G., J. Janssens, C. Opsomer and J. Botterman. 1995. Inhibition of fungal disease development in plants by engineering controlled cell death. Bio/Technol. 13: 1085-1089. [ Links ]
Tailor, R.H., D.P. Acland, S. Attenborough, B. Cammue, I.J. Evans, R.W. Osborn, J.A. Ray, S.B. Rees and W.F. Broekaert. 1997. A novel family of small cysteine-rich antimicrobial peptides from seed of Impatiens balsamina is derived from a single precursor protein. Journal of Biological Chemistry 272(39): 24480-24487. [ Links ]
Thevissen, K., D. C. Warnecke, I.E.J.A. Francois, M. Leipelt, E. Heinz, C. Ott, U. Zahringer, B.P.H.J. Thomma, K.K.A. Ferket and B. Cammue. 2004. Defensins from insects and plants interact with fungal glucosylceramides. Journal of Biological Chemistry 279(6): 3900-3905. [ Links ]
Thevissen, K., J. Idkowiak-Baldys, Y.J. Im, J. Takemoto, I.E.J.A. François, K.K.A. Ferket, A.M. Aerts, E.M.K. Meert, J. Winderickx, J. Roosen and B. Cammue. 2005. SKN1, a novel plant defensin-sensitivity gene in Saccharomyces cerevisiae, is implicated in sphingolipid biosynthesis. FEBS Letters 579(9): 1973–1977.
Xing, H., C.B. Lawrence, O. Chambers, H.M. Davies, N.P. Everett and Q.Q. Li. 2006. Increased pathogen resistance and yield in transgenic plants expressing combinations of the modified antimicrobial peptides based on indolicidin and magainin. Planta 223(5): 1024-1032. [ Links ]

