










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.1 Medellín Jan./June 2010
DISTRIBUCIÓN DEL AGUA LLUVIA EN TRES BOSQUES ALTOANDINOS DE LA CORDILLERA CENTRAL DE ANTIOQUIA, COLOMBIA
RAINFALL DISTRIBUTION IN THREE HIGH ANDEAN FORESTS IN THE CENTRAL CORDILLERA OF COLOMBIA
Juan Diego León Peláez1; Maria Isabel González Hernández2 y Juan Fernando Gallardo Lancho3
1 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. Departamento de Ciencias Forestales. A.A. 1779, Medellín, Colombia. <jdleon@unalmed.edu.co>
2 Profesora Titular. Universidad de Salamanca, España. <mimg@usal.es>
3 Investigador. IRNA/CSIC. Salamanca, España. <jgallard@usal.es>
Recibido: Marzo 11 de 2009; Aceptado: Mayo 10 de 2010
Resumen. Se estudió la distribución del agua lluvia durante tres años en un bosque climácico de roble (Quercus humboldtii) y dos plantaciones forestales, de pino (Pinus patula) y ciprés (Cupressus lusitanica) ubicados en los Andes colombianos. Los principales ingresos de agua al bosque ocurrieron por la lluvia, cuyo valor medio anual fue 1.700 mm año-1. La precipitación interna tuvo la mayor participación de los flujos hídricos del bosque, siendo máxima en el cipresal (1.486 mm a-1, 90 %) y mínima en el pinar (1.389 mm a-1, 81 %), mientras que el escurrimiento cortical fue muy bajo en los tres bosques (1-3 mm a-1, 0,1-0,2 %). La máxima intercepción se presentó en el pinar (329 mm a-1), equivalente al 19 % de la lluvia. A diferencia de estos flujos hídricos del dosel los flujos hídricos del suelo sí mostraron diferencias significativas entre coberturas (P <0,05). En el robledal se encontró el patrón característico de disminución de la lámina de agua con la profundidad y las pérdidas por drenaje profundo fueron mínimas (2 mm a-1). Los flujos hídricos del dosel mostraron una fuerte correlación con la cantidad e intensidad de lluvia. En conclusión, el robledal mostró mayor potencial regulador del agua lluvia, seguido por el pinar. El cipresal mostró un muy bajo potencial regulador, con altas y rápidas pérdidas de agua por drenaje profundo.
Palabras clave: Precipitación interna, escurrimiento cortical, precipitación neta, drenaje profundo, balance hidrológico, bosques montanos, Quercus humboldtii.
Abstract. Rainfall distribution was assessed for three years in a mature oak forest (Quercus humboldtii), pine (Pinus patula) and cypress (Cupressus lusitanica) plantations in the northern part of the colombian Andean mountains. An annual average of 1700 mm was the main water input in the forests, which came from the bulk precipitation. The higher water flows were represented by throughfall, which showed a maximum value in the cypress (1486 mm y1, 90%) and a minimum one in the pine (1389 mm y1, 81%) plantations, respectively. Stemflow was very low in the three forest types (1-3 mm y1, 0.1-0.2%). The maximum amount of water intercepted by forest crowns was found in the pine plantation (329 mm y1), which accounted for 19% out of the total rainfall. In opposition to the patterns found for throughfall and stemflow, soil hydric flows did show significant differences among forest types (P<0.05). In the oak forest water infiltration inversely decreased with soil depth; this forest type showed the lowest water losses by deep drainage (2 mm y1). Finally, throughfall and stemflow were significantly correlated with the amount and intensity of rainfall. In conclusion, oak forests showed the highest rainfall regulation capacity, followed by the pine plantation. In contrast, the cypress plantation had a very low rainfall regulation potential once that this forest type was characterized by high and fast water losses by deep drainage.
Key words: throughfall, stemflow, net precipitation, deep drainage, hydrological balance, montane forests, Quercus humboldtii.
La principal entrada de agua al bosque ocurre a través de la lluvia o precipitación incidente (Pp). La diferencia entre ésta (que alcanza el follaje) y la que llega al piso forestal se conoce como agua de intercepción del dosel arbóreo (I), y constituye una pérdida en la medida que desde allí se evapora (Cavelier y Vargas, 2002). Otra proporción de la lluvia que atraviesa el dosel (o cae desde las copas de los árboles) y alcanza el piso del bosque se considera precipitación interna (Pi), mientras que la que resbala por los troncos de los árboles o arbustos hacia abajo, depositándose en las bases de éstos, se denomina escurrimiento cortical (EC) (Parker, 1983; Koichiro et al., 2001; Wang et al., 2004). La precipítación neta (Pn), es el resultado de la sumatoria entre la Pi y el Ec (Pn= Pi + Ec), se ha utilizado para evaluar la capacidad del dosel para almacenar agua (Cavelier et al., 1997). Cada uno estos flujos hídricos interactúa de un modo particular con la vegetación, generando en ellos cambios tanto cuantitativos como cualitativos (Cavelier et al., 1997; Wang et al., 2005). Estos cambios dependen de factores relacionados con características propias del bosque (composición, estructrua, edad, etc) y ambientales (intensidad y duración de la precipitación, vientos, etc.) (Carlyle, 2004).
En los bosques templados, el agua aportada al piso por la Pn representa entre 70 y 90 % de la Pp (Moreno et al., 1996a; Levia y Frost, 2003). En los bosques tropicales esta variación es mayor dada la alta variabilidad de factores que allí regulan estos flujos. A diferencia de la Pi, que representa la mayor proporción de la Pn, el Ec es muy bajo, entre 0,1 y 2,1 % (Bruijnzeel, 1990; Veneklaas y Van Ek, 1990; Cavelier y Vargas, 2002), habiéndose estudiado poco (Levia y Frost, 2003). Sin embargo, en los bosques de niebla el Ec puede ser el 10% de la Pp, como resultado de la precipitación horizontal (Weaver, 1972; Bruijnzeel y Proctor, 1995). Además, se ha destacado su importancia relativa (Bruijnzeel, 1989; Levia y Frost, 2003; Carlyle, 2004) si se compara con la cantidad de lluvia interceptada. No toda la Pn se incorpora al suelo mineral a través de la infiltración, ya que parte de ella puede convertirse en escorrentía superficial (Es) y otra parte quedar retenida en el mantillo. No obstante, la Es ha sido usualmente señalada como de muy bajas proporciones en bosques donde el mantillo es apreciable (McGregor, 1980; Maene y Sulaiman, 1980; Alegre et al., 1990; Ruiz et al., 2005a, 2005b). En el mantillo de los bosques tropicales suelen presentarse grandes cantidades de raíces finas como estrategia competitiva de las especies vegetales en procura de nutrientes de escasa disponibilidad en el suelo (Herrera et al., 1978; Cuevas y Medina, 1988; Barreto y León, 2005), con lo cual se consume parte del agua que atraviesa el horizonte O.
Una vez en el suelo mineral el agua circula rápidamente por los macroporos del suelo originados por raíces muertas y por actividad animal, y muy lentamente por los poros capilares (Voigt, 1980; Seven y Germann, 1981; Moreno et al., 1996b). En los bosques lluviosos tropicales, pueden ocurrir pérdidas significativas de agua y nutrientes vía drenaje profundo (Dp), como sucede en los bosques templados que reciben grandes cantidades de lluvia (Reynolds et al., 1990; Menéndez et al., 1995). A pesar de la reconocida importancia de ambos flujos (aéreos y subsuperficiales) en el balance hidrológico del bosque, en los trópicos su estudio ha sido relativamente modesto al compararlo con el de bosques templados (Muoghalu, 2003). Tales estudios son escasos en los bosques altoandinos a pesar de su importancia en la regulación de caudales de fuentes abastecedoras de agua de áreas densamente pobladas, dada su localización en las partes altas de las cuencas hidrográficas.
La deforestación de estas áreas ha ocurrido a gran escala, poniendo en riesgo la regulación de caudales, así como la alta diversidad y endemismo que caracterizan a los bosques allí presentes (Gentry y Dodson, 1987; Henderson et al., 1991). De hecho en Colombia los bosques altoandinos, se encuentran entre los cinco ecosistemas forestales más vulnerables a la conversión de acuerdo con las predicciones de área transformada (Etter et al., 2006). De aquí la necesidad de obtener un conocimiento adecuado del funcionamiento hidrológico de los remanentes de estos bosques, así como del que ocurre en las plantaciones forestales que usualmente se realizan en estos pisos altitudinales para la regulación hidrológica en áreas de influencia de proyectos hidroeléctricos.
En este trabajo se analiza la distribución del agua lluvia a través de diferentes rutas como una aproximación al balance hidrológico de tres bosques de los Andes Colombianos, localizados en la cuenca hidrográfica de Piedras Blancas, antigua abastecedora de agua para la ciudad de Medellín.
El estudio se realizó durante tres años consecutivos en un robledal climácico (bosque dominado por Q. humboldtii) y en dos plantaciones forestales maduras de pino (Pinus patula) y ciprés (Cupressus lusitanica). Se propusieron como objetivos: i. Comparar la distribución del agua lluvia (Pp) entre coberturas mediante los procesos de intercepción (I), precipitación interna (Pi) y escurrimiento cortical (Ec); ii. Determinar la magnitud de los cambios que experimentan las aguas que alcanzan el piso del bosque vía Pn, a medida que atraviesan el mantillo, se infiltran en el suelo (Is) y se constituyen en pérdidas en el balance hidrológico (Dp); y iii. Modelar estos flujos a partir de diferentes variables de la lluvia y de la vegetación.
MATERIALES Y MÉTODOS
Sitios de estudio. El estudio se realizó desde Marzo de 2001 hasta Marzo de 2004 en la cuenca hidrográfica de Piedras Blancas (Antioquia, Colombia), a una distancia de 25 km de la ciudad de Medellín y a una elevación de 2.490 msnm (06º 18” N y 75º 30” O). Las características físicas y climáticas, y las de los bosques estudiados fueron ampliamente descritas por Londoño et al., (2005, 2007) y Zapata et al. (2007).
Procedimientos de campo. En cada bosque se estableció una parcela de 5.000 m2 en la cual se instalaron sistemáticamente 10 pluviómetros para captar la precipitación interna (Pi) más 2 pluviómetros en los alrededores de cada fragmento para registrar la precipitación a campo abierto (Pp). Cada pluviómetro se construyó con un tubo de PVC de 15,2 cm de diámetro. Dentro del tubo se instaló un embudo de igual diámetro, recubierto de una malla fina plástica, acoplado a una manguera para conducir las aguas recogidas hasta un recipiente plástico. La altura de instalación de cada pluviómetro fue de 80 cm sobre la superficie del terreno. Para el monitoreo de las aguas que escurren por los troncos (escurrimiento cortical, Ec) se instalaron 12 collarines en cada bosque, procurando que quedaran representados con estos, proporcionalmente, las diferentes tallas diamétricas de los árboles presentes. Los collarines consistieron en mangueras plásticas de 7,6 cm de diámetro seccionadas por la mitad en su longitud. Cada collarín se dispuso helicoidalmente en cada árbol, garantizando el total cubrimiento de su diámetro. Se fijaron con grapas y clavos galvanizados, previo pulido de la superficie de la corteza y, para lograr un adecuado movimiento del agua desde el tronco del árbol hasta el collarín, el área de contacto entre ambas superficies se recubrió con silicona. Las aguas captadas se condujeron hasta recipientes plásticos, cuya capacidad, según el diámetro del árbol (DAP), osciló entre 10 y 60 L.
Para el estudio de los flujos hídricos del suelo, se conformaron dos calicatas (2 m x 1m) en cada bosque (Londoño et al., 2005; 2007). En sus perfiles se instalaron 6 lisímetros plásticos, 2 por nivel de monitoreo. La primera capa la constituyó el mantillo (horizonte O), denominándose como Ho el agua que lo trascola. El segundo nivel se localizó -30 cm por debajo del anterior para captar las aguas de infiltración superficial (Is) correspondientes al horizonte A. El último nivel se localizó a una profundidad de -80 cm desde la superficie del suelo para monitorear las pérdidas por drenaje profundo (Dp). Los lisímetros aquí empleados fueron de succión cero y consistieron en bandejas de 20 x 30 cm2 seccionadas e introducidas en el suelo siguiendo la dirección de la pendiente topográfica, conectadas mediante mangueras plásticas a recipientes plásticos. Estos lisímetros permiten recoger tanto el flujo vertical como el horizontal y corresponden con la descripción de un lisímetro ideal (Keller y Vedy, 1991) consistente en un dispositivo de tensión cero (abierto) de pequeña talla, inerte y con escasa demanda de mantenimiento. La necesidad de definir el tipo de agua recolectada es un aspecto que determina la elección del tipo de lisímetro a utilizar en un estudio (Londoño et al., 2007). Los lisímetros con tensión recogen agua retenida a potenciales difíciles de conocer y con una pauta un tanto aleatoria, mientras que los lisímetros sin tensión sólo recogen agua que percola a través del perfil, es decir, agua insaturada (Haines et al., 1982), siendo esta última la que constituye la principal forma de pérdida de agua del suelo. Para disminuir la evaporación del perfil del suelo, éste se recubrió con sábanas de polipropileno blanco.
En cada punto de muestreo de los flujos Is y Dp del perfil se instalaron bloques de yeso con electrodos en acero inoxidable para registrar semanalmente el contenido de humedad (lector Delmhorst KS-D10). Durante el estudio se midieron con una frecuencia semanal (mediante probetas plásticas graduadas) la totalidad de los volúmenes recogidos por los diferentes dispositivos (pluviómetros, collarines y lisímetros). La información para las variables de intensidad de la precipitación fue tomada de la Estación Chorrillos de EPM (842.280 E, 1.188.220 N, 2.370 msnm).
Tratamiento de datos y análisis estadísticos. Los volúmenes semanales de los flujos de Pp, Pi, Ho, Is y Dp, fueron convertidos de las unidades de registro (L) a lámina de agua (mm), considerando para esto el área individual de recepción de los dispositivos que funcionaron correctamente durante cada período semanal. Para la Ec el volumen de agua recolectado en cada bosque fue asociado al área basal de los árboles cuyos collarines funcionaron adecuadamente en cada semana de muestreo. A partir del área basal real de cada rodal estos valores se llevaron a lámina de agua en el flujo de Ec. Con estos valores, se determinó la participación de cada flujo dentro del bosque y se calculó la intercepción (Moreno et al., 1996a):
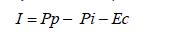
Las láminas de los flujos de los tres bosques se compararon mediante ANAVAS con medidas repetidas en el tiempo, empleando el programa SAS 8.0. Para ello, cada dispositivo fue considerado como una seudo-repetición, teniéndose sobre cada uno de estos una medida repetida en el tiempo. Cuando los datos presentaron heterocedasticidad, los valores fueron transformados mediante logaritmo natural. Cuando se presentaron diferencias en los flujos, éstas se determinaron con la prueba de Tukey (P =0,05). Mediante análisis de regresión y correlación se determinó la existencia de relaciones entre los flujos en cada bosque y diferentes presentaciones de la lluvia, tanto de cantidad como de intensidad. Por su potencial incidencia en la generación de la Pn y sobre la circulación del agua en el suelo se determinaron, entre otras, las siguientes variables: a) Intensidad máxima de la lluvia semanal (Imax); b) Intensidad promedio de los tres eventos de lluvia de máxima intensidad semanal (Iprom3max); c) Lámina de agua asociada a los eventos de lluvia semanal de intensidad promedio (CIprom); y d) Número de eventos de lluvia semanal con intensidad mayor a 10 mm h-1 (Evmayor10). Para evitar problemas de multicolinealidad entre estas variables, se realizó el proceso stepwise de eliminación de variables. Adicionalmente para el Ec se ajustaron modelos de regresión con variables características dasonómicas de los árboles individuales. Para ello se midieron en campo el área y diámetro de la copa (AC y DC, respectivamente), la altura (H), y el diámetro a la altura del pecho (DAP).
RESULTADOS
Flujos hídricos del dosel. De los tres periodos estudiados, el tercero fue el más lluvioso (Tabla 1 y Figura 1); sin embargo, las diferencias entre los valores anuales no fueron significativas (P >0,05). Los máximos de precipitación ocurrieron entre los meses de Marzo-Mayo y Septiembre-Noviembre, configurando un régimen bimodal de precipitación. En promedio, la Pi representó con respecto a la Pp, 85 % en el robledal, 81 % en el pinar y 90 % en el cipresal. La Pi mostró un comportamiento similar entre bosques y una estrecha correlación con la Pp (Tabla 2), sin presentarse diferencias significativas en sus valores (P >0,05).
Tabla 1. Lámina media anual (±error estándar) de los flujos hídricos del dosel en bosques altoandinos. Valores expresados en mm.

Pp: Precipitación incidente, Pi: precipitación interna, Ec: escurrimiento cortical, I: intercepción. Letras diferentes en cada flujo denotan diferencias significativas entre bosques (P <0,05). Los valores entre paréntesis representan su porcentaje respecto al total de lluvia.
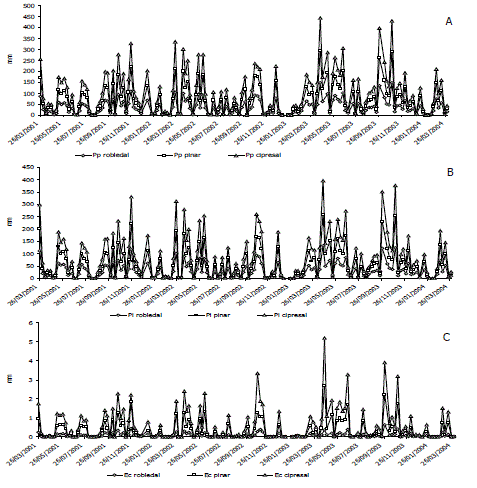
Figura 1. Evolución de los flujos hídricos del dosel (mm) en bosques altoandinos. A: Precipitación incidente (Pp); B: Precipitación interna (Pi); C: Escurrimiento cortical (Ec).
Tabla 2. Coeficientes de correlación lineal de Pearson* para la precipitación interna (Pi) y el escurrimiento cortical (Ec) en tres bosques altoandinos.
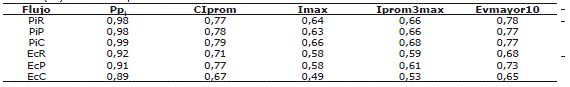
*Todas las correlaciones significativas (P <0,05), PiR: precipitación interna semanal en el robledal, PiP: Precipitación interna semanal en el pinar, PiC: Precipitación interna semanal en el cipresal, EcR: Escurrimiento cortical semanal en el robledal, EcP: Escurrimiento cortical semanal en el pinar, EcC: Escurrimiento cortical semanal en el cipresal, Ppi: precipitación promedio semanal para cada bosque, CIprom: cantidad de agua asociada a los eventos de lluvia semanal de intensidad promedio, Imax: intensidad máxima de la lluvia semanal, Iprom3max: Intensidad promedio de los tres eventos de lluvia de máxima intensidad semanal, Evmayor10: número de eventos de lluvia semanal con intensidad >10 mm h-1.
La Pi se correlacionó significativamente (Tabla 2 y Figura 2) con la CIprom. La correlación entre la Iprom3max y la Pi, aunque menor (r = 0,67 en promedio para los tres bosques) fue también significativa. La representación del Ec de la Pp fue muy baja. Los valores medios de Ec del robledal (0,9 mm a-1) fueron inferiores a los de las plantaciones (3,0 mm a-1); no obstante, el ANAVA no mostró diferencias significativas entre ellos. El Ec y la Pi estuvieron muy correlacionadas con la Pp (Tabla 2). La mayor I se encontró en el pinar (19 %) y la menor en el cipresal (10 %) sin presentarse diferencias significativas entre los tres bosques (P>0,05). En muy pocas semanas la Pn fue mayor que la Pp (Figura 3); de las cerca de 150 semanas que comprendió el estudio ello ocurrió en 16 de ellas en el robledal, sólo 9 en el pinar y 15 semanas en el cipresal.
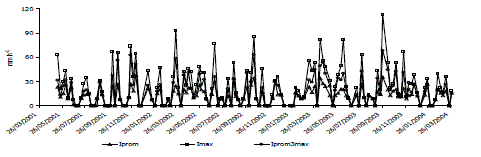
Figura 2. Presentaciones de la intensidad de las lluvias durante el periodo de estudio. Iprom: intensidad promedio de la lluvia semanal, Imax: intensidad máxima de la lluvia semanal, Iprom3max: intensidad promedio para los tres eventos de lluvia de máxima intensidad semanal. Datos procedentes de la estación meteorológica Chorrillos de EPM.
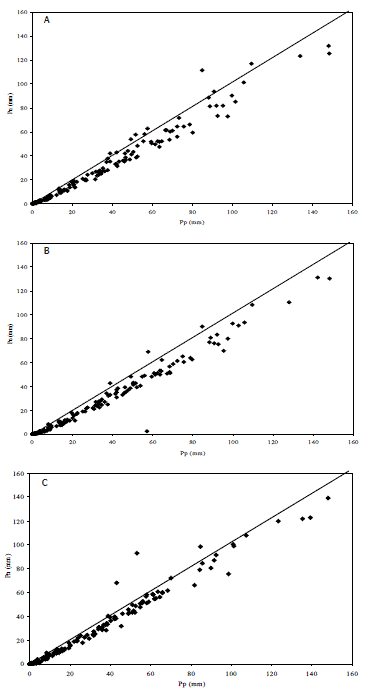
Figura 3. Relaciones entre los valores semanales de precipitación incidente (Pp) y precipitación neta (Pn) en bosques altoandinos. A: Robledal; B: Pinar; C: Cipresal.
Flujos hídricos del suelo. La Pn constituye mayoritariamente el agua que podría incorporarse a los flujos hídricos del suelo. Aunque la mayor lámina de Pn se encontró en el cipresal (Tabla 3) en este sistema no se verificó la máxima infiltración en el mantillo (Ho), sino en el robledal climácico. La participación del flujo Ho siguió la secuencia decreciente siguiente: robledal > cipresal > pinar, siendo muy superiores los valores del robledal. Esta situación se invirtió para las aguas de infiltración superficial (Is) en el robledal, donde su participación en relación con la Pn fue de 9%. El Dp fue muy superior en el cipresal, casi 30% de la Pn, en el pinar fue inferior al 2% y en el robledal prácticamente despreciable. En conjunto los flujos hídricos del suelo siguieron el patrón de lluvias, con valores picos asociados a los de la Pp (Figura 4). Para estos flujos (Ho, Is, Dp) las diferencias fueron significativas entre los tres bosques (P <0,05). En los tres bosques el flujo Ho se correlacionó significativa y directamente con la Pn y con la CIprom, con máximos absolutos en el robledal (Tabla 4). La Is en las plantaciones de coníferas se correlacionó fuertemente con estas dos variables, así como con la Imax. Únicamente en el cipresal se registraron correlaciones significativas entre el Dp y las dos variables mencionadas. No se encontraron correlaciones significativas entre el contenido de humedad (CHi) y el Dp.
Tabla 3. Lámina media anual (±error estándar) de los flujos hídricos del suelo en bosques altoandinos. Valores expresados en mm.

Ho: aguas que atraviesan el mantillo, Is: infiltración superficial, Dp: drenaje profundo. Letras diferentes en cada flujo denotan diferencias significativas entre bosques (P <0,05). Los valores entre paréntesis representan su porcentaje del agua neta del bosque (Pi + Ec).
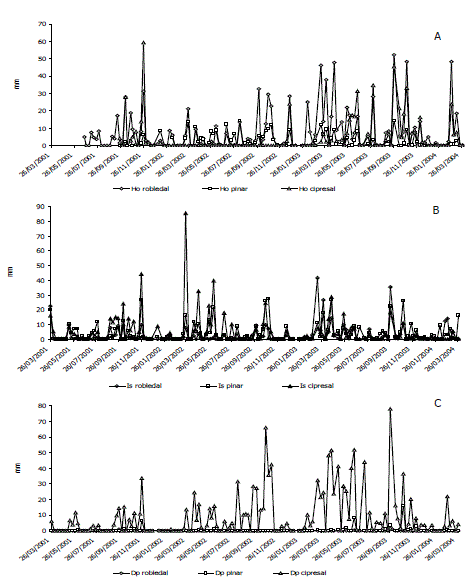
Figura 4. Evolución de los flujos hídricos del suelo (mm) en bosques altoandinos. A: Aguas que atraviesan el mantillo (Ho); B: Infiltración superficial (Is); C: Drenaje profundo (Dp).
Tabla 4. Coeficientes de correlación lineal de Pearson para la infiltración en el mantillo (Ho), la infiltración superficial (Is) y el drenaje profundo (Dp) en tres bosques altoandinos.
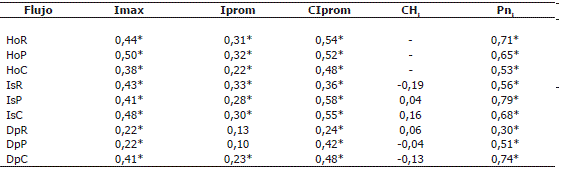
*Correlaciones significativas (P <0,05), HoR: aguas que atraviesan el mantillo del robledal, HoP: aguas que atraviesan el mantillo del pinar, HoC: aguas que atraviesan el mantillo del cipresal, IsR: infiltración superficial en el robledal, IsP: infiltración superficial en el pinar, IsC: infiltración superficial en el cipresal, DpR: drenaje profundo del robledal, DpP: drenaje profundo del pinar, DpC: drenaje profundo del cipresal, Imax: intensidad máxima de la lluvia semanal, Iprom: intensidad promedio de la lluvia, CIprom: cantidad de agua asociada a los eventos de lluvia semanal de intensidad promedio, CHi: contenido de humedad del suelo en cada profundidad y bosque, Pni: precipitación neta en cada bosque.
Modelación de los flujos hídricos. Todos los modelos de regresión ajustados para Pi y Ec fueron significativos (Tabla 5). A diferencia de las ecuaciones para Pi, las relativas a Ec (con excepción del robledal) mostraron una mayor complejidad. Dado que fue notoria la mayor dependencia del Ec (además de las características de las lluvias) de las variables dasonómicas de cada árbol individual al cual estaba fijado cada collarín, se incluyeron en los modelos ambos tipos de variables (Tabla 6). Los modelos fueron significativos (P <0,05), no mostraron problemas de autocorrelación en los residuales, y siempre quedaron en ellos como variables el DAP y la Pp. Los modelos de regresión lineal obtenidos para la I (Tabla 7) fueron asimismo todos significativos (P <0,05), reflejando una clara similitud entre bosques y quedando incluidas en todos ellos como variables la Pp y la intensidad de las lluvias (Iprom, Iprom3max). Todos los modelos ajustados para los flujos Ho, Is y Dp fueron significativos (Tabla 8). Las variables de cantidad (Pi) y de intensidad (Imax, Iprom) de la lluvia, quedaron incluidas en la mayoría de modelos.
Tabla 5. Modelos de regresión lineal ajustados para los flujos hídricos en bosques altoandinos en función de variables de la lluvia*
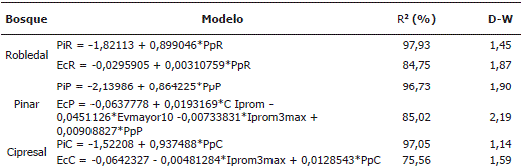
*Todos los modelos significativos (P <0,05); PiR: precipitación interna semanal en el robledal, PiP: precipitación interna semanal en el pinar, PiC: precipitación interna semanal en el cipresal, EcR: escurrimiento cortical semanal en el robledal, EcP: escurrimiento cortical semanal en el pinar, EcC: escurrimiento cortical semanal en el cipresal, PpR: precipitación semanal en el robledal, PpP: precipitación semanal en el pinar, PpC: precipitación semanal en el cipresal, C Iprom: cantidad de agua asociada a los eventos de lluvia semanal de intensidad promedio, Evmayor 10: número de eventos de lluvia con intensidad mayor de 10 mm h-1, Iprom3max: intensidad promedio para los tres eventos de lluvia de máxima intensidad semanal, R2: coeficiente de determinación ajustado, D-W: estadístico Durbin–Watson.
Tabla 6. Modelos de regresión lineal ajustados para el escurrimiento cortical (Ec) en tres bosques altoandinos en función de variables dasonómicas del árbol y la lluvia*

* Todos los modelos significativos (P <0,05), EcR: escurrimiento cortical semanal en el robledal, EcP: escurrimiento cortical semanal en el pinar, EcC: escurrimiento cortical semanal en el cipresal, PpR: precipitación promedio semanal en el robledal, PpP: precipitación promedio semanal en el pinar, PpC: precipitación promedio semanal en el cipresal, H: altura del árbol correspondiente a cada collarín, AC: área de copa del árbol correspondiente a cada collarín, DAP: diámetro a la altura del pecho del árbol correspondiente a cada collarín, DC: diámetro de copa del árbol correspondiente a cada collarín, R2: Coeficiente de determinación ajustado, D-W: Estadístico Durbin–Watson.
Tabla 7. Modelos de regresión lineal ajustados para la intercepción (I) en tres bosques altoandinos*

*Todos los modelos significativos (P <0,05); IR: intercepción del robledal, IP: intercepción del pinar, IC: intercepción del cipresal, PpR: precipitación semanal en el robledal, PpP: precipitación semanal en el pinar, PpC: precipitación semanal en el cipresal, Iprom: intensidad promedio de la lluvia semanal., Iprom 3max: intensidad promedio para los tres eventos de lluvia de máxima intensidad semanal, R2: coeficiente de determinación ajustado, D-W: estadístico Durbin–Watson.
Tabla 8. Modelos de regresión lineal ajustados para los flujos hídricos del suelo en tres bosques altoandinos.*

*Todos los modelos significativos (P <0,05), HoR: aguas que atraviesan el mantillo del robledal, IsR: infiltración superficial en el robledal, DpR: drenaje profundo del robledal, HoP: aguas que atraviesan el mantillo del pinar, IsP: infiltración superficial en el pinar, DpP: drenaje profundo del pinar, HoC: aguas que atraviesan el mantillo del cipresal, IsC: infiltración superficial en el cipresal, DpC: drenaje profundo del cipresal, PiR: precipitación interna en el robledal, PiP: precipitación interna en el pinar, PiC: precipitación interna en el cipresal, Imax: intensidad máxima de la lluvia semanal, Iprom: intensidad promedio de la lluvia semanal, Ciprom: cantidad de agua asociada a los eventos de lluvia semanal de intensidad promedio, CH: contenido de humedad semanal del suelo, Pp: precipitación incidente, R2: coeficiente de determinación, D-W: estadístico Durbin-Watson.
DISCUSIÓN
Flujos hídricos del dosel. Los valores de Pi encontrados fueron todos superiores al 80% de la Pp (Tabla 1), coincidiendo con los observados en otros estudios (76-94%) realizados en bosques montanos tropicales (Jackson, 1971; Lundgren y Lundgren, 1979; Steinhardt, 1979; Edwards, 1982; Mamanteo y Veracion, 1985; Vis, 1986; Stadtmüller y Agudelo, 1990; Veneklaas, 1990), y con los sugeridos por Bruijnzeel (1990) de 75 a 86%, para bosques lluviosos tropicales de tierras bajas del mundo. La magnitud de la Pi guarda una estrecha relación con la composición del bosque ya que la arquitectura del dosel determina su capacidad para almacenar agua. Hölscher et al. (1998) mostraron cómo, mientras en un bosque con alta diversidad la Pi era del 60-80 % de la Pp, en otro con una única especie estos valores disminuyeron hasta el 20-40 %. No obstante, para las plantaciones forestales aquí estudiadas los valores de Pi fueron muy similares a los del robledal.
La estrecha correlación entre la Pi y la Pp quedó evidenciada mediante los modelos de regresión, cuyas pendientes fueron muy cercanas a la unidad, revelando una fuerte proporcionalidad entre ambas variables (especialmente en el cipresal). No obstante, las relaciones porcentuales entre Pi y Pp pueden ser altamente variables, consecuencia de diversos factores que las regulan: estructura del rodal, estado de humedad del bosque antes de la lluvia, diferencias en intensidad y duración de la lluvia y condiciones de evaporación (tanto entre los distintos períodos muestreados, como en las condiciones particulares de los sitios) (Edwards, 1982; Hafkenscheid, 2000; Tobón et al., 2000; Tobón y Arroyave, 2008).
A diferencia de la Pi el Ec tuvo una participación muy baja en la Pn del bosque, por lo que usualmente se considera marginal (Levia y Frost, 2003; Koichiro et al., 2001; Carlyle, 2004). De hecho, los valores de Ec aquí encontrados (0,1-0,2%) fueron muy bajos, y similares a los encontrados por Veneklaas y Van Ek (1990) (0,1%) en bosques montanos de Colombia. Por otra parte, fueron inferiores a los encontrados por Cavelier y Vargas (2002) para bosques neotropicales del mundo (2,1 %), por Bruijnzeel (1990); 1-2 %, para bosques lluviosos tropicales del mundo y por Bruijnzeel (2001) para bosques nublados (1-2%). No obstante, al comparar con valores de Ec de otros estudios, debe considerarse el problema que representa la falta de normalización metodológica para calcular las láminas correspondientes. Aunque se ha sugerido (Edwards, 1982) el cálculo por área basal como el indicador más fiable, persiste como inconveniente relacionar los volúmenes recogidos con el DAP del individuo muestreado.
Algunos estudios (Weaver, 1972; Lloyd y Marques, 1988; Moreno et al., 1996a) han encontrado bajos valores de Ec para árboles con mayores DAP como consecuencia (Bruijnzeel, 1989) de los patrones de ramificación de los árboles de mayor tamaño, así como por la posibilidad de que el goteo desde las copas de los árboles que sobresalen en el dosel sea interceptado por árboles de menor tamaño, localizados por debajo de aquellos. Por el contrario, otros autores (Jordan, 1978; Jordan y Heyveldop, 1981; Lloyd y Marques, 1988; Hölscher et al., 1998) observaron una relación directa entre el tamaño del árbol y el Ec. Aunque las diferencias en el Ec entre bosques fueron muy bajas en términos absolutos éstas probablemente fueron resultado, a su vez, de diferencias en la arquitectura de los árboles presentes en cada bosque. El grado de inclinación de las ramas en su inserción con los tallos y el carácter monopódico de las coníferas puede influir más notoriamente en la generación del Ec que las copas amplias y ramificadas del robledal. No obstante, aunque el Ec está influenciado por factores del rodal como la composición de especies (Raich, 1983; Carlyle, 2004) y el estadío sucesional (Hölscher et al., 1998) la variabilidad en las cantidades de Ec parece ser mayor dentro de un mismo rodal que entre diferentes bosques (Levia y Frost, 2003). Se ha destacado al respecto la influencia que puede ejercer en las magnitudes del Ec el número de árboles por unidad de área que alcanzan el dosel superior (Ulrich, 1983; Hölscher et al., 2003). Esta característica determina los mayores valores de Ec aquí encontrados en las plantaciones, dado que su coetaneidad y mayor uniformidad estructural se reflejan en un mayor número de individuos dominantes que los del robledal. Adicionalmente, la mayor complejidad en cuanto a estratos intermedios y mayor epifitismo del robledal favorecen la ocurrencia de menores valores de Ec.
Liu et al., (2002) encontraron incrementos en el Ec hasta del 40% al eliminarse las epífitas. De hecho, los modelos de regresión para Ec (Tablas 5 y 6) revelaron el papel mitigador de las epífitas sobre la intensidad de las lluvias en el robledal, ya que esta variable no quedó allí incluida en los modelos a diferencia de los de las plantaciones de coníferas. La mayor complejidad del Ec se reflejó en los modelos de regresión en los que fue necesario incorporar además de variables de la lluvia (Pp), características dasonómicas de los árboles individuales (DAP, altura, diámetro y área de la copa). El signo positivo asociado a la Pp indicó su proporcionalidad directa con el Ec. Por el contrario, los modelos indicaron una disminución en el Ec a medida que el DAP fue mayor. No obstante, aún no se han estudiado profundamente los procesos que controlan la generación del Ec. Koichiro et al. (2001) encontraron paradójicamente que la cantidad debajo de las copas era mayor que la que se recogía en la base de los árboles. Esto muestra que la capacidad de almacenamiento de la corteza puede afectar los volúmenes recogidos y el carácter específico de este tipo de flujo que implica un contacto íntimo entre el agua y la vegetación. Algunos autores (Parker, 1983; Levia y Frost, 2003) consideraron que la rugosidad de la corteza es incluso más importante en la definición del flujo que la magnitud de la lluvia, mientras que otros (Carlyle, 2004) indican lo contrario. Aunque ambos factores son fundamentales (Crockford y Richardson, 1998) la dominancia de uno u otro está relacionada con características específicas del rodal y del clima.
La I del robledal (250 mm a-1, 15 % de la lluvia) fue intermedia entre la máxima obtenida en el pinar (329 mm a-1, 19%) y la mínima del cipresal (170 mm a-1, 10 %). Dicho valor I del robledal fue muy próximo a los obtenidos por Veneklaas (1990) para bosques de tierras altas de Colombia (262-265 mm a-1, 12,4-18,2 %), así como al promedio para un grupo de bosques tropicales (14,8%) (Cavelier y Vargas, 2002). A partir de varios estudios Bruijnzeel (1989) encontró un valor medio anual de I del 14% (4,5-22,0 %) para bosques tropicales de tierras bajas (n = 13) y de 18 % (10-24 %) para bosques montanos (n = 5); según este autor el mayor valor encontrado en los bosques montanos podría reflejar las intensidades de las lluvias usualmente menores que se producen a tales elevaciones. De los modelos de regresión (Tabla 7) se evidenció una relación inversa entre la I y características de la lluvia (Pp, Iprom) como resultado de la disminución en la capacidad de las copas de los árboles para retener agua a medida que aumenta la cantidad de lluvia y su intensidad promedio, lo cual coincide con otros hallazgos (Moreno et al., 1996b; Liu et al., 2002) en bosques lluviosos.
La máxima I del pinar probablemente fue resultado del gran número de individuos que alcanzan el dosel, con gran capacidad de almacenamiento de agua por sus copas, como puede deducirse de su mayor área basal y biomasa (pinar: 41,7 m2 ha-1 y 328 Mg ha-1; robledal: 17,3 m2 ha-1 y 166 Mg ha-1; y cipresal: 36,6 m2 ha-1 y 195 Mg ha-1, respectivamente). En los bosques donde abundan las briofitas y las epífitas el dosel posee una mayor capacidad de almacenamiento de agua, lo que podría provocar mayores pérdidas por I (Edwards, 1982). Las epífitas tienen la capacidad de incrementar la I al aumentar el área del dosel o al consumir el agua (Liu et al., 2002). De hecho, las epífitas de las ramas y troncos de los árboles pueden absorber hasta 500 veces su peso seco, con lo cual la capacidad de intercepción del dosel aumenta con el incremento en la biomasa de epífitas (Kőhler et al., 2007; Tobón y Arroyave, 2008). A pesar de la notoria presencia de epífitas en el robledal la mayor complejidad de su estratificación vertical probablemente incide en que la intercepción del agua por su dosel no sea tan alta como la del pinar.
Al comparar entre bosques la Pn con la Pp (Figura 3) se encontró una gran similitud. En el robledal el número de semanas que mostraron valores Pn mayores que los de Pp fue de 16, valor muy similar al del cipresal (15) y muy superiores ambos al del pinar (9). No obstante, este orden no se conservó para los valores absolutos, ya que en el cipresal la suma de todos los valores semanales en que se verificó Pn > Pp fue 83 mm, en el robledal 59 mm y en el pinar 21 mm. Si bien esto denota la ocurrencia del proceso de precipitación horizontal en los tres bosques, ésta no significó una fuente importante de agua, ya que con respecto a los ingresos por lluvia representó 1,1% en el robledal, 0,4% en el pinar y 1,7% en el cipresal. La relación entre la Pn y la Pp ha sido además empleada para determinar el valor de saturación del dosel (Cavelier et al., 1997). Para la resolución semanal que tuvieron las observaciones realizadas, no se encontró una clara diferencia entre bosques (Figura 3), siendo su valor cercano a 3 mm.
Flujos hídricos del suelo. En el robledal se encontró la máxima cantidad de Pn que atravesó el mantillo (Ho), siendo sus valores más de tres veces superiores a los del pinar (Tabla 1). En los bosques con potente mantillo suelen presentarse tasas altas de infiltración dado que el suelo se encuentra protegido del impacto de la lluvia, tiene una buena capacidad de conservación del agua y, con ésta, menores pérdidas por escorrentía superficial (Alegre y Rao, 1996; Hartanto et al., 2003). A pesar de que el espesor del mantillo del robledal (45 cm) y del pinar (40 cm) fueron muy similares las grandes diferencias entre los valores Ho de ambos bosques sugieren la ocurrencia de mayores pérdidas de agua por consumo por las raíces finas del mantillo; sin embargo, dicho proceso no explica satisfactoriamente la situación descrita, ya que en los sitios de estudio (León, 2007; León et al., 2008) la masa de raíces finas presentes en el mantillo del robledal (3,91 Mg ha-1) fue muy superior a la del pinar (0,96 Mg ha-1) o a la del cipresal (1,65 Mg ha-1).
Otra hipótesis sería una mayor escorrentía superficial en el pinar; dado que la Pn fue muy similar en los tres bosques y que las mayores diferencias se presentaron entre el robledal y las plantaciones de coníferas es probable que las características físicas y químicas de los detritos orgánicos del mantillo disminuyan el flujo Ho, favoreciendo o frenando la escorrentía superficial. La mayor uniformidad, forma lineal y pequeño diámetro de los órganos foliares de las especies de coníferas generan de hecho un fuerte empaquetamiento del mantillo, lo cual representa una barrera física para la infiltración del agua a través de él. Adicionalmente, el proceso de descomposición de materiales orgánicos de carácter ácido puede generar compuestos hidrofóbicos con alta repelencia al agua (DeBano, 2000). De hecho ha sido verificada la ocurrencia de repelencia al agua en plantaciones de P. patula y C. lusitanica de la zona de estudio, tanto en el mantillo como en los horizontes minerales del suelo (Jaramillo et al., 2000; Duque et al., 2004). Aunque todos los coeficientes de correlación de Pearson fueron significativos para el flujo Ho y la Pn (Tabla 4), fueron más bajos los valores para las plantaciones de coníferas. Los modelos de regresión para Ho (Tabla 8) mostraron fuerte correlación negativa con la intensidad máxima de la lluvia en el pinar, lo cual evidencia la baja capacidad del mantillo para favorecer la percolación y permitir al agua alcanzar la superficie del suelo, lo cual implica escorrentía superficial conforme la intensidad de la lluvia sea mayor.
La tendencia decreciente de las láminas de agua de infiltración con la profundidad sólo ocurrió con claridad en el robledal. El agua que se infiltró en el horizonte A (Is) representó una reducción del 62% del agua correspondiente a Ho. Una más clara reducción del flujo Ho hacia los horizontes minerales más profundos (Dp) también se encontró en el robledal, representando la casi totalidad del agua que entró en contacto con el suelo. La citada reducción en la lámina de agua que atraviesa el mantillo con la profundidad tiene entre sus principales explicaciones el almacenamiento temporal en el suelo, el consumo por la vegetación y la escorrentía superficial. Sin embargo, en las plantaciones de coníferas estudiadas dicho patrón se mostró alterado, ya que las láminas correspondientes a Is fueron superiores a las de Ho y, en el caso del Dp, la lámina de agua del cipresal fue 2,5 veces mayor.
La repelencia al agua verificada para estas plantaciones de coníferas por Duque et al. (2004) puede originar flujos preferenciales de agua por los macroporos del suelo (en el caso que la estructura de la hojarasca lo permita, al no estar empaquetada laminarmente) que permitirían la recepción de mayores cantidades de agua y solutos en niveles subsuperficiales del suelo (Jaramillo, 2003); no obstante, la aparición de vías preferenciales de flujos hídricos favorecen los frentes irregulares de humedecimiento (Wallis y Horne, 1992), ocasionando mayores y más rápidas circulaciones de agua a través de los macroporos hasta los horizontes inferiores edáficos, reduciéndose así las posibilidades para su almacenamiento como agua útil en el suelo. La presencia de horizontes hidrofóbicos empaquetados puede afectar negativamente las propiedades hidráulicas de la zona no saturada, modificando la curva característica del suelo (DiCarlo et al., 1999; Bauters et al., 2000). Aunque el contenido de humedad del suelo es, entre otros factores, determinante de la infiltración, esta variable únicamente quedó incluida en el modelo de Dp del pinar. Además, de la posible incidencia de la hidrofobicidad sobre esta situación los efectos que un mismo contenido de humedad puede producir en distintos suelos dependen de las características particulares de estos y de los antecedentes climáticos (Regalado et al., 2003).
Balance hídrico. Los principales ingresos de agua ocurrieron a través de la Pp y fueron muy similares en los tres bosques (Tabla 1). Los ingresos por precipitación horizontal fueron mínimos, por lo que se consideraron despreciables. Aunque ha sido ampliamente aceptado que la Es bajo cubierta forestal es mínima, Ruiz et al. (2005a) determinaron pérdidas anuales en estos mismos rodales con relación a la Pp, de 1,1, 1,6 y 11,1% en el robledal, en el pinar y en el cipresal, respectivamente. En este estudio los citados valores representaron (en el mismo orden) láminas medias de Es de 19,0, 27,6 y 184,0 mm a-1. Las pérdidas de agua (P) del bosque (calculadas como P = I + Dp +Es) fueron máximas en el cipresal (790 mm a-1), donde significaron cerca del 50% de la Pp. En el pinar fueron de 376 mm a-1 (22%) y en el robledal 271 mm a-1 (16%).
El agua retenida por el mantillo (calculada como 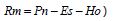 ) se puede consumir por la vegetación o almacenar temporalmente en los diferentes subhorizontes que lo conforman (Oi, Oe y Oa) (Wilcke et al., 2002). Conforme las aguas atraviesan el mantillo y entran en contacto con el horizonte A del suelo se pueden eventualmente presentar pérdidas por Es al no poder infiltrarse en él. La retención por el mantillo fue máxima en el pinar (1.262 mm a-1, 73% de la Pp), seguida por el robledal (1.128 mm a-1, 65% de la Pp) y el cipresal (1.136 mm a-1, 69% de la Pp). El consumo de agua por la vegetación se produce en mayor medida a través de las raíces finas que ocupan tanto el mantillo como los horizontes más superficiales del suelo. De hecho en los rodales aquí estudiados las raíces finas presentes en el mantillo y en el horizonte A edáfico representaron (con respecto al total) 80 % en el robledal, 78 % en el pinar y 85 % en el cipresal (León et al., 2008).
) se puede consumir por la vegetación o almacenar temporalmente en los diferentes subhorizontes que lo conforman (Oi, Oe y Oa) (Wilcke et al., 2002). Conforme las aguas atraviesan el mantillo y entran en contacto con el horizonte A del suelo se pueden eventualmente presentar pérdidas por Es al no poder infiltrarse en él. La retención por el mantillo fue máxima en el pinar (1.262 mm a-1, 73% de la Pp), seguida por el robledal (1.128 mm a-1, 65% de la Pp) y el cipresal (1.136 mm a-1, 69% de la Pp). El consumo de agua por la vegetación se produce en mayor medida a través de las raíces finas que ocupan tanto el mantillo como los horizontes más superficiales del suelo. De hecho en los rodales aquí estudiados las raíces finas presentes en el mantillo y en el horizonte A edáfico representaron (con respecto al total) 80 % en el robledal, 78 % en el pinar y 85 % en el cipresal (León et al., 2008).
Como una aproximación gruesa para estimar conjuntamente el consumo por la vegetación y el almacenamiento de agua en el suelo (Cva) se consideraron como representativas de ambos procesos el agua Rm y la que permaneció en el horizonte A (calculada como 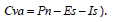 ). El máximo valor Cva se obtuvo en el robledal (1.330 mm a-1; representando el 77% de la Pp) y el mínimo correspondió al cipresal (1.073 mm a-1, 65% de la Pp). En el pinar el Cva fue 1.162 mm a-1 (68% de la Pp). No obstante, si además se computan las pérdidas por Dp (como una aproximación conjunta que involucra todo el perfil del suelo monitoreado por lisimetría) en el caso del cipresal los valores de Cva bajan considerablemente, al ser máximas las pérdidas por drenaje profundo.
). El máximo valor Cva se obtuvo en el robledal (1.330 mm a-1; representando el 77% de la Pp) y el mínimo correspondió al cipresal (1.073 mm a-1, 65% de la Pp). En el pinar el Cva fue 1.162 mm a-1 (68% de la Pp). No obstante, si además se computan las pérdidas por Dp (como una aproximación conjunta que involucra todo el perfil del suelo monitoreado por lisimetría) en el caso del cipresal los valores de Cva bajan considerablemente, al ser máximas las pérdidas por drenaje profundo.
CONCLUSIONES
El aporte principal de agua en los tres bosques (robledal, cipresal y pinar) ocurrió a través de la lluvia, la cual contabilizó en promedio, cerca de 1.700 mm a-1 y se distribuyó en un patrón bimodal a lo largo del año con mínimas diferencias interanuales.
La precipitación interna siguió una pauta muy similar a la lluvia y representó (con relación a ésta) un promedio de 85% en el robledal, 81% en el pinar y 90% en el cipresal, sin registrarse entre ellas diferencias significativas. Aunque se registraron eventos de precipitación horizontal su ocurrencia y aporte al balance hídrico fue muy escaso, sin obedecer a un patrón temporal definido.
En los tres bosques el escurrimiento cortical tuvo poca incidencia cuantitativa en el ciclo hidrológico, siendo máximo en el cipresal con cerca de 0,2% del agua de lluvia.
Las pérdidas por intercepción y drenaje profundo fueron mucho mayores en los bosques de coníferas que en el robledal.
El análisis conjunto de los resultados permite señalar una mayor eficiencia del robledal para distribuir el agua lluvia dentro del bosque, seguido del pinar y, muy distantes ambos, del cipresal.
Principalmente en el robledal (y, en segunda medida, en el pinar) las menores pérdidas de agua fuera del sistema y la alta retención de ésta por el suelo propician condiciones favorables para la regulación de caudales en estas áreas altoandinas.
Los modelos ajustados a los flujos hídricos del dosel del bosque (precipitación interna, escurrimiento cortical e intercepción) mostraron, en términos generales, una fuerte dependencia de la cantidad y la intensidad de la lluvia. En particular, para el escurrimiento cortical, se evidenció adicionalmente una correlación importante con características de cada árbol individual, especialmente en función de su grosor y tamaño de la copa. Los modelos correspondientes a los flujos hídricos del suelo revelaron una dependencia diferencial según la profundidad de la cantidad e intensidad de la lluvia, y de las láminas de agua procedentes de horizontes superiores.
AGRADECIMIENTOS
Este trabajo fue financiado por la DIME, CORANTIOQUIA y el Departamento de Ciencias Forestales de la Universidad Nacional de Colombia. Los autores agradecen al personal de la Estación Forestal Piedras Blancas (Universidad Nacional de Colombia, Sede Medellín) su ayuda, a Empresas Públicas de Medellín por permitir el acceso a los sitios de estudio y por el suministro de información meteorológica; además, se agradece de manera especial a Diana Marcela Otálvaro y Joan Alexander Correa su participación en el montaje y monitoreo del experimento.
BIBLIOGRAFÍA
Alegre, J.C. and M.R. Rao. 1996. Soil and water conservation by contour hedging in the humid tropics of Peru. Agriculture, Ecosystem and Environment 57(1): 17-25. [ Links ]
Alegre, J.C., C.F. Morales and B. La Torre. 1990. Soil erosion studies in Peru. Journal of Soil and Water Conservation 45(3): 417-420. [ Links ]
Barreto, L.H. y J.D. León. 2005. Masa total y contenido de nutrientes en raíces finas de ecosistemas forestales (Pinus patula Schltdl y Cham., Cupressus lusitanica Mill. y Quercus humboldtii Bonpl.) de Piedras Blancas, Antioquia-Colombia. Revista Facultad Nacional de Agronomía 58(2): 2907-2929. [ Links ]
Bauters, T.W.J., T.S. Steenhuis, D.A. Dicarlo, J.L. Nieber, L.W. Dekker, C.J. Ritsema, J.Y. Parlange and R. Haverkamp. 2000. Physics of water repellent soils. Journal of Hydrology 231-232: 233-243. [ Links ]
Bruijnzeel, L.A. 1989. Nutrient cycling in moist tropical forests: the hydrological framework. pp. 383-415. In: Proctor, J. (ed.). Mineral nutrients in tropical forest and savanna ecosystems. Blackwell Sci. Publ., Oxford. [ Links ]
Bruijnzeel, L.A. 1990. Hydrology of moist tropical forests and effects of conversion: a state of knowledge review. IHP-UNESCO Humid Tropical Programme, París. 224 p.
Bruijnzeel, L.A. 2001. Hydrology of tropical montane cloud forests: a reassessment. Land Use and Water Resources Reasearch 1: 1.1-1.18. [ Links ]
Bruijnzeel, L.A. and J. Proctor. 1995. Hydrology and biogeochemistry of tropical montane cloud forests: what do we really know?. pp. 38-78. In: Hamilton, L.S., J.O. Juvik and F.N. Scatena (eds.). Tropical montane cloud forests. Springer, New York. 407 p.
Carlyle, D.E. 2004. Throughfall, stemflow, and canopy interception loss fluxes in a semi-arid Sierra Madre Oriental matorral community. Journal of Arid Environments 58(2): 181-202. [ Links ]
Cavelier, J., M. Jaramillo, D. Solis and D. León. 1997. Water balance and nutrient inputs in bulk precipitation in tropical montane cloud forest in Panama. Journal of Hydrology 193(1-4): 83-96. [ Links ]
Cavelier, J. y G. Vargas. 2002. Procesos hidrológicos. pp. 145-165. En: Guariguata, M. y G. Kattan (eds.). Ecología y conservación de bosques tropicales. Libro Universitario Regional, Cartago, Costa Rica. [ Links ]
Crockford, R.H. and D.P. Richardson. 1998. Litterfall, litter and associated chemistry in a dry sclerophyll eucalypt forest and a pine plantation in south-eastern Australia: 2. Nutrient recycling by litter, throughfall and stemflow. Hydrological Processes 12(3): 385-400. [ Links ]
Cuevas, E. and E. Medina. 1988. Nutrient dynamics within Amazonian forests. II. Fine root growth, nutrient availability and leaf litter decomposition. Oecologia 76(2): 222-235. [ Links ]
DeBano, L.F. 2000. Water repellency in soils: a historical overview. Journal of Hydrology 31: 4-32. [ Links ]
DiCarlo, D.A., T.W.J. Bauters, C.J.G. Darnault, T.S. Steenhuis and J.Y. Parlante. 1999. Rapid determination of constitutive relations with fingered flow: pp. 433-440. In: Proceedings International Workshop on Characterization and Measurement of the Hydraulic Properties of Unsaturated Porous Media. Riverside, CA. [ Links ]
Duque, J.R., S.P. Arbeláez, D.F. Jaramillo y J.D. León. 2004. Estudio de hidrofobicidad de suelos en robledales (Quercus humboldtii) y plantaciones forestales (Pinus patula y Cupressus lusitanica) en la cuenca de la quebrada Piedras Blancas. Revista Facultad Nacional de Agronomía 57(1): 2423-2434. [ Links ]
Edwards, P.J. 1982. Studies of mineral cycling in a montane rain forest in New Guinea. V. Rates of cycling in throughfall and litter fall. Journal of Ecology 70(3): 807-827. [ Links ]
Etter, A., C. McAlpine, K. Wilson, S. Phinn and H. Possingham. 2006. Regional patterns of agricultural land use and deforestation in Colombia. Agriculture, Ecosystems and Environment 114(2-4): 369-386. [ Links ]
Gentry, A.H. and D. Dodson. 1987. Contribution of nontrees to species richness of a tropical rain forest. Biotropica 19(2): 149-156. [ Links ]
Hafkenscheid, R. 2000. Hydrology and biogeochemistry of montane rain forests of contrasting stature in the Blue Mountains of Jamaica. PhD thesis, Vrije Universiteit, Amsterdam. 315 p. [ Links ]
Haines, B.L., J.B. Waide and R.L. Todd. 1982. Soil solution nutrient concentrations sampled with zero-tension lysimeters: report of discrepancies. Soil Science Society of America Journal 46(3): 658-661. [ Links ]
Hartanto, H., R. Prabhu, A. Widayat and C. Asdak. 2003. Factors affecting runoff and soil erosion: plot-level soil loss monitoring for assessing sustainability of forest management. Forest Ecology and Management 180(1-3): 361-374. [ Links ]
Henderson, A. S.P. Churchill y J.L. Luteyn. 1991. Neotropical plant diversity. Nature 351: 21-22. [ Links ]
Herrera, R.A., C.F. Jordan, H. Klinge and E. Medina. 1978. Amazon ecosystems: their structure and functioning with particular emphasis on nutrients. Interciencia 3: 223-232. [ Links ]
Hölscher, D., T.D.de.A. Sá, R.F. Möller, M. Denich and H. Fölster. 1998. Rainfall partitioning and related hydrochemical fluxes in a diverse and a mono specific (Phenakospemum guyannense) secondary vegetation stand in eastern Amazonia. Oecologia 114: 251-257. [ Links ]
Hölscher, D., L. Köhler, C. Leuschner and M. Kapelle. 2003. Nutrient fluxes in stemflow and throughfall in three successional stages of an upper montane rain forest in Costa Rica. Journal of Tropical Ecology19: 557-565. [ Links ]
Jackson, I.J., 1971. Problems of throughfall and interception assessment under tropical forest. Journal of Hydrology 12(3): 234-254 [ Links ]
Jaramillo, D.F., L.W. Dekker, C.J. Ritsema and J.M.H. Hendrickx. 2000. Occurrence of soil water repellency in arid and humid climates. Journal of Hydrology 231-232: 105-111. [ Links ]
Jaramillo, D.F. 2003. Efecto de la temperatura de secado del suelo sobre la repelencia al agua en Andisoles bajo cobertura de Pinus patula. Informe de investigación. Escuela de Geociencias. Universidad Nacional de Colombia, Medellín. 36 p. [ Links ]
Jordan, C.F. 1978. Stemflow and nutrient transfer in a tropical rain forest. Oikos 31: 257-263. [ Links ]
Jordan, C.F. and J. Heyveldop. 1981. The water budget of an Amazonian rain forest. Acta Amazonica 11(1): 87-92. [ Links ]
Keller, C. and J.C. Vedy. 1991. Apport de la lisimétrie sans tensions paur l’étude des transferts de Cu et Cd dans le sols forestiers faiblement pollués. Science du Sol 29: 107-124.
Kőhler, L., C. Tobón, K.F.A. Frumau and L.A. Bruijnzeel. 2007. Biomass and water storage dynamics of epiphytes in old-growth and secondary montane cloud forest stands in Costa Rica. Plant Ecology 193(2): 171-184. [ Links ]
Koichiro, K., T. Yuri, T. Nobuaki and K. Isamu. 2001. Generation of stemflow volume and chemistry in a mature Japanese cypress forest. Hydrological Processes 15(19): 1967-1978. [ Links ]
León, J.D. 2007. Contribución al conocimiento del ciclo de nutrientes en bosques montanos naturales de Quercus humboldtii y reforestados (Pinus patula y Cupressus lusitanica) de la región de Piedras Blancas, Antioquia (Colombia). Universidad de Salamanca, Facultad de Ciencias Agrarias y Ambientales. Salamanca, España. 410 p. [ Links ]
León, J.D., M.I. González y J.F. Gallardo. 2008. Caída de hojarasca, descomposición y liberación de nutrientes en un bosque montano de roble (Quercus humboldtii Bonpl.) en los Andes colombianos. pp. 97-124. En: León, J.D. (ed.). Ecología de bosques andinos: Experiencias de investigación. La Carreta Editores, Medellín. [ Links ]
Levia, D.F. and E.E. Frost. 2003. A review and evaluation of stemflow literature in the hydrologic and biogeochemical cycles of forested and agricultural ecosystems. Journal of Hydrology 274(1-4): 1-29. [ Links ]
Liu, W., E.D. Fox and Z. Xu. 2002. Biomass and nutrient accumulation in montane evergreen broad-leaved forest (Lithocarpus xylocarpus type) in Ailao mountains, SW China. Forest Ecology and Management 158(1-3): 223-235. [ Links ]
Lloyd, C.R. and F.A. Marques. 1988. Spatial variability of throughfall and stemflow measurements in Amazonian rain forest. Agricultural and Forest Meteorology 42(1): 63-73. [ Links ]
Londoño, A., D.C. Montoya, J.D. León y M.I. González. 2005. Estudio del flujo saturado de agua en suelos forestales de Piedras Blancas, Antioquia. Boletín de Ciencias de la Tierra 17: 73-84. [ Links ]
Londoño, A., D.C. Montoya, J.D. León y M.I. González. 2007. Estudio del flujo saturado de agua en suelos forestales de Piedras Blancas, Antioquia. Revista Facultad Nacional de Agronomía 60(1): 3717-3734. [ Links ]
Lundgren, L. and B. Lundgren, 1979. Rainfall interception and evaporation in the Mazumbai forest reserve, West Usambara mountains, Tanzania and their importance in the assessment of land potential. Geografiska Annaler 61(3-4): 157-178. [ Links ]
Maene, L.M. and W. Sulaiman. 1980. Status of soil conservation research in peninsular Malaysia and its future development. pp. 202-219. In: Morgan, R.P.C. (ed.). Soil erosion and its control. Van Nostrand Reinhold, New York. 311 p.
Mamanteo, B.P. and V.P. Veracion. 1985. Measurements of fog dril, throughfall and stemflow in the mossy and Benguet pine (Pinus kesiya Royle ex Gordon) forests in the upper Agno river basin. Sylvatrop (Philippines Forest Research Journal) 10, 271-282. [ Links ]
McGregor, D.F.M. 1980. An investigation of soil erosion in the Colombian rain forest zone. Catena 7(1): 265-273. [ Links ]
Menéndez, I., G. Moreno, J.F. Gallardo and J. Saavedra. 1995. Soil solution composition in forest soils of Sierra de Gata mountains, Central-Western Spain: Relationship with soil water content. Arid Soil Research and Rehabilitation 9: 495-502. [ Links ]
Moreno, G., J.F. Gallardo, F. Ingelmo, S. Cuadrado and J. Hernández. 1996a. Soil water budget in four Quercus pyrenaica forest across a rainfall gradient. Arid Soil Research and Rehabilitation 10(1): 65-84. [ Links ]
Moreno, G., J.F. Gallardo, K. Schneider and F. Ingelmo. 1996b. Water and bioelement fluxes in four Quercus pyrenaica forest along a pluviometric gradient. Annales des Sciences Forestières 53(2-3): 625-639. [ Links ]
Muoghalu, J.I. 2003. Contributions of throughfall, stemflow and litterfall to nutrient cycling in a secondary lowland rain forest in Ile-Ife, Nigeria. Journal of Tropical Forest Science 15: 399-410. [ Links ]
Parker, G.G. 1983. Throughfall and stemflow in the forest nutrient cycle. Advances in Ecological Research 13: 57-133. [ Links ]
Raich, J.W. 1983. Throughfall and stemflow in mature and year-old wet tropical forest. Tropical Ecology 24: 234-243. [ Links ]
Regalado, C.M., G. García, J.M. Hernández, A. Pérez y A.R. Socorro. 2003. Caracterización de la zona no saturada de un bosque maduro de laurisilva en el parque nacional de Garajonay: hidrofobicidad e implicaciones hidrológicas. Estudios de la Zona no Saturada del Suelo 6: 193-200. [ Links ]
Reynolds, B., J.A. Hudson and G. Leeks. 1990. Field methods for estimating solute and sediment losses in small upland streams. pp. 103-129. In: Harrison, A.F. and P. Ineson (eds.). Nutrients cycling in terrestrial ecosystems. Field methods application and interpretation. Elsevier Applied Science, London. [ Links ]
Ruiz, O.A., J.J. Acosta y J.D. León. 2005a. Escorrentía superficial en bosques montanos naturales y plantados de Piedras Blancas, Antioquia (Colombia). Revista Facultad Nacional de Agronomía 58(1): 2635-2649. [ Links ]
Ruiz, O.A., J.J. Acosta y J.D. León. 2005 b. Flujo de nutrientes en la escorrentía superficial de bosques montanos en Piedras Blancas, Antioquia (Colombia). Meteorología Colombiana 9: 23-29. [ Links ]
Seven, K. y P. Germann, 1981. Water flow in soil macropores, 2, A combined flow model. Journal of Soil Science 32(1): 15-29. [ Links ]
Stadtmüller, T. and N. Agudelo. 1990. Amount and variability of cloud moisture input in a tropical cloud forest. International Association of Scientific Hydrology Publ. 193: 25-32. [ Links ]
Steinhardt, U. 1979. Untersuchungen über den wasser- und nährstoffhaushalt eines andinen wolkenwaldes in Venezuela. Göttinger Bodenkundliche Berichte 56: 1-185. [ Links ]
Tobón, C., W. Bouten and J. Sevink. 2000. Gross rainfall and its partitioning into throughfall, stemflow and evaporation of intercepted water in four forest ecosystems in western Amazonia. Journal of Hydrology 237(1-2): 40-57. [ Links ]
Tobón, C. y F. Arroyave. 2008. Hidrología de los bosques altoandinos. pp. 227-260. En: León J.D. (ed.). Ecología de bosques andinos: Experiencias de investigación. La Carreta Editores, Medellín. [ Links ]
Ulrich, B. 1983. Interaction of forest canopies with atmospheric constituens: SO2, alkali and earth alkali cations and chloride. pp. 33-45. In: Ulrich, B. y J. Pankrath (eds.). Effects of accumulation of air pollutants in forest ecosystems. D. Reidel Publ., Dordretch. 389 p. [ Links ]
Veneklaas, E.J. 1990. Nutrient fluxes in bulk precipitation and throughfall in two montane tropical rain forests, Colombia. Journal of Ecology 78(4): 974-992. [ Links ]
Veneklaas, E.J. y R. Van Ek, 1990. Rainfall interception in two tropical montane rain forest, Colombia. Hydrological Processes 4(4): 311-326. [ Links ]
Vis, M. 1986. Interception, drop size distributions and rainfall kinetic energy in four Colombian forest ecosystems. Earth Surface Processes and Landforms 11(6): 591-603. [ Links ]
Voigt, G.K. 1980. Acid precipitation and soil buffering capacity. pp. 53-57. In: Drabløs, S. and A. Tollan (eds.). Ecological impact of acid precipitation. Proceedings of a International Conference Ecological Impact of Acid Precipitation. Oslo, Noruega. 383 p. [ Links ]
Wallis, M.G. and D.J. Horne. 1992. Soil water repellency. Advances in Soil Science 20: 91-146. [ Links ]
Wang, M.C., C.P. Liu and C.H. Sheu. 2004. Characterization of organic matter in rainfall, throughfall, stemflow, and streamwater from three subtropical forest ecosystems. Journal of Hydrology 289(1-4): 275-285. [ Links ]
Weaver, P.L. 1972. Cloud moisture interception in the Luquillo mountains of Puerto Rico. Caribbean Journal of Science 12: 129-144. [ Links ]
Wilcke, W., S. Yasin, U. Abramowski, C. Valarezo and W. Zech. 2002. Nutrient storage and turnover in organic layers Andean tropical montane rain forest in Ecuador. European Journal of Soil Science 53(1): 15-27. [ Links ]
Zapata, C.M., J.A. Ramírez, J.D. León y M.I. González. 2007. Producción de hojarasca fina en bosques de Quercus humboldtii, Pinus patula y Cupressus lusitanica de Antioquia, Colombia. Revista Facultad Nacional de Agronomía 60(1): 3771-3784. [ Links ]

