










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.2 Medellín July/Dec. 2010
EFECTO DE DIFERENTES NIVELES Y ÉPOCAS DE DEFOLIACIÓN SOBRE EL RENDIMIENTO DE LA PAPA (SOLANUM TUBEROSUM CV. PARDA PASTUSA)
EFFECT OF DIFFERENT LEVELS AND TIMES OF DEFOLIATION ON THE POTATO YIELD (SOLANUM TUBEROSUM CV. PARDA PASTUSA)
Daniel Rodríguez Caicedo1; Marcela Sofía Rico Tellez2; Luis Ernesto Rodríguez Molano3 y Carlos Eduardo Ñústez López4
1 Profesor Asistente. Universidad Militar Nueva Granada. Facultad de Ciencias Básicas. Carrera 11 No. 101-80, Bogotá, Colombia. <daniel.rodriguez@unimilitar.edu.co>
2 Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Agronomía. Carrera 45 No. 26-85, Bogotá, Colombia.
3 Profesor Asociado. Universidad Nacional de Colombia. Facultad de Agronomía. Carrera 45 No. 26-85, Bogotá, Colombia. <lerodriguezmo@unal.edu.co>
4 Profesor Asociado. Universidad Nacional de Colombia. Facultad de Agronomía. Carrera 45 No. 26-85, Bogotá, Colombia. <cnustez@unal.edu.co>
Recibido: Septiembre 4 de 2009; Aceptado: Mayo 4 de 2010
Resumen. Se estudió el efecto de cuatro niveles de defoliación (20, 40, 60 y 80%) sobre el rendimiento de tubérculo en papa cv. Parda Pastusa, en tres épocas de desarrollo del cultivo (90, 120 y 150 días después de siembra (dds)), con el objetivo de analizar en forma simulada el efecto de los daños en el follaje causados por enfermedades o insectos plaga sobre el rendimiento. Se utilizaron parcelas de plantas sin defoliar como testigo. El experimento se realizó en el Centro Experimental ICA San Jorge, ubicado en el municipio de Soacha (Cundinamarca) a 3100 msnm y se evaluó el rendimiento comercial de tubérculo. Los resultados demostraron que no se presenta disminución en el rendimiento de tubérculo para ninguno de los niveles de defoliación evaluados cuando se realizó a los 90 o a los 150 dds; por el contrario, cuando se realizó a los 120 dds y en un nivel mayor o igual al 40%, se generó una importante reducción en la cantidad de tubérculos cosechados.
Palabras clave: Daño foliar, tubérculo, andigena, producción.
Abstract. The effect of four defoliation levels was studied (20, 40, 60 and 80%), in three times of crop’s development (90, 120 and 150 days after sowing (das)) on the tuber yield in Parda Pastusa cv. with the objective of analyzing in simulated form the effect of the damage in leaves caused by deseases or insects on the yield. Plants without damage were used like witness. The research was carried out in ICA San Jorge Research Center, located in Soacha (Cundinamarca) to 3200 masl and the commercial tuber yield was evaluated. The results demonstrated that decrease is not presented in the tuber yield for none of the evaluated defoliation levels when it is carried out at the 90 or the 150 das. On the contrary, when it is carried out to the 120 das and in a bigger level or similar to 40%, important decrease is generated in the tuber yield.
Key words: Foliar damage, tuber, andigena, yield.
La producción de tubérculos en el cultivo de la papa depende básicamente de tres factores: a) La cantidad de energía solar interceptada por el cultivo, b) la eficiencia de la planta para la conversión de dicha energía en materia seca y c) la eficiencia en la traslocación de la materia seca a los tubérculos (Midmore, 1981). Las plagas y enfermedades del follaje afectan al primer factor, pues al reducir el área foliar, se altera la capacidad para captar la luz, principalmente por medio de las hojas, y en menor medida por los tallos (Wellik, 1981). Esto a su vez puede tener un efecto negativo sobre los rendimientos, ya que se genera una respuesta de compensación en la que la planta redistribuye parte de la materia seca para producir hojas nuevas, con el fin de equilibrar el daño generado por la pérdida de tejidos fotosintéticamente activos, lo que implica una posible reducción en la distribución de materia seca para los tubérculos (Midmore, 1981).
En papa la defoliación puede ser dividida en: a) Completa, cuando la eliminación de los tejidos permite el paso libre de la luz solar hacia las hojas ubicadas en la parte inferior, como la generada por Leptinotarsa decimlineata, Diabrotica sp. o Spodoptera sp. b) Incompleta, cuando la luz solar no puede atravesar los tejidos dañados para llegar a las hojas inferiores; es el tipo de daño generado por Liriomyza huidobrensis, Scrobipalpula absoluta y Phthorimaea operculella (Robert, 1999).
El daño causado por Phytophthora infestans debe ser considerado con más detalle, pues es más complejo que el generado por los insectos. En las condiciones de Colombia, el hongo afecta durante todo el desarrollo vegetativo hojas, tallos y en ocasiones tubérculos, dependiendo principalmente del clima. Cuando las condiciones ambientales no favorecen al patógeno (tiempo frío y seco), la infección avanza con lentitud, las hojas atacadas se arrugan y los conidióforos casi no son conspicuos. Por el contrario, si la humedad y la temperatura son altas, las áreas necrosadas de ramas hojas y peciolos crecen rápidamente y en pocos días pueden presentarse valores de incidencia y severidad cercanos al 100% (Zapata y Trillos, 1997). Cuando la enfermedad afecta el follaje, ocasiona un daño indirecto, ya que al reducirse las funciones fotosintéticas hay una disminución en el rendimiento (Urquijo y Landaluze, 1961).
P. infestans es el principal patógeno del cultivo de la papa en Colombia, pues se siembran cv. susceptibles, tales como ICA Nevada, Tuquerreña, Parda Pastusa y Diacol Capiro, y para su control, se requieren entre 10 y 18 aspersiones de fungicidas por ciclo de cultivo, lo cual incrementa notablemente los costos de producción (Zapata y Trillos, 1997).
Conociendo el tipo de daño generado por este patógeno y en forma somera el que producen algunas plagas de insectos, conviene mencionar las limitaciones y alcances concernientes a los modelos de daño simulado (Metcalf y Luckman, 1982).
En un modelo de este tipo las hojas son eliminadas en un solo instante, mientras que el consumo de follaje por los insectos, o el daño por P. infestans ocurren por un periodo de días o incluso semanas, lo cual hace que la pérdida de los tejidos se produzca en una forma gradual, que la planta puede compensar con más facilidad. Por otro lado, el área foliar perdida por la acción de insectos y enfermedades es discontinua, y se manifiesta como un conjunto de agujeros o zonas necróticas de diferentes tamaños y formas, mientras que en estos modelos se eliminan hojas enteras o parte de las mismas. Sin embargo, como una primera aproximación para la estimación del efecto del daño por insectos, han dado buenos resultados en cultivos como la soya y la papa en Estados Unidos (Metcalf y Luckman, 1982).
En el caso de la “gota”, la aproximación del modelo a la situación real es más complicada, dado que el patógeno tiene mecanismos de producción de enzimas y toxinas que también afectan a los tejidos, y que no se toman en cuenta en el modelo. Además, el daño no se limita a las hojas, sino que puede afectar también los tallos, situación que es mucho más compleja para su evaluación pues influyen factores tales como el número de hojas presentes en el tallo, y si este es principal o secundario.
En el presente estudio solamente se evalúa el daño realizado en las hojas por medio de la eliminación de foliolos y cada planta se somete a una sola defoliación durante todo el ciclo del cultivo. Esto último podría resultar semejante a la defoliación producida por un sólo ataque de gota o de una plaga del follaje durante el ciclo del cultivo.
El efecto de la defoliación en el rendimiento total de la papa ha sido evaluado en la Estación Experimental Agrícola de Minnesota por Cranshaw y Radcliffe (1980) sobre los cv. Red Pontiac, Norchip, Russet Burbank y Anoka, pertenecientes a la subespecie tuberosum. En el estudio se consideraron como factores los tiempos de desarrollo fenológico, homogeneidad o heterogeneidad en el patrón de defoliación, realización del daño en una o varias ocasiones durante el ciclo del cultivo y defoliación en estratos diferentes de la planta (superior, medio e inferior). En estos experimentos se pudo determinar que el efecto del daño depende en gran medida de la época. El tratamiento aplicado cuando la tasa de crecimiento vegetativo disminuye y comienza el llenado de los tubérculos, generó la mayor disminución en el rendimiento. Así mismo se observó que no hubo efectos importantes al realizar la defoliación en épocas anteriores o posteriores al llenado de tubérculos y que porcentajes de defoliación inferiores al 67% no afectaron la producción de manera significativa, incluso en el periodo de llenado de tubérculos, si bien hubo diferencias según los cv.
En estudios acerca del impacto de la defoliación por el L. decimlineata se estableció que la producción de tubérculos se redujo en un 67% cuando se realizó defoliación del 100% en la etapa media de desarrollo del cultivo, mientras que la reducción en producción fue mínima si el daño ocurre antes o después (Hare, 1980). Este resultado se atribuyó a que la planta pudo recuperarse en forma virtualmente completa al ocurrir la defoliación en etapas tempranas de desarrollo y, al presentarse en los estados tardíos, el llenado de tubérculos había finalizado, por lo que no hubo efecto adverso sobre el rendimiento al remover el follaje. En dicho estudio se trabajó con el cv. Katahdin, perteneciente a la subespecie tuberosum.
Wellik, Slosser y Kirby (1981) encontraron en estudios de simulación de daño foliar por insectos, trabajando con plantas del cv. Red La Soda (subespecie tuberosum) que durante la estación media de desarrollo, éstas presentaron su mayor susceptibilidad a la defoliación.
La respuesta al daño también puede variar según el estrato que resulte afectado. Cranshaw y Radcliff (1980) encontraron que la remoción de las hojas del estrato superior generó un mayor descenso en la producción que cuando el daño ocurrió en los estratos medio o inferior.
También se estableció que la defoliación en el estrato medio siempre produjo rendimientos iguales o superiores al testigo, mientras que aplicada al estrato inferior produjo rendimientos menores, pero solo cuando las plantas se encontraban en la mitad de su desarrollo fenológico. Los mismos autores plantean que las hojas del estrato superior son las más eficientes en términos de su capacidad fotosintética, y por lo tanto su eliminación generó una reducción importante en el rendimiento.
El efecto que se da por la eliminación de las hojas del estrato inferior en la etapa intermedia de desarrollo del cultivo, no se relaciona tanto con su capacidad fotosintética sino con su contenido de hormonas, pues ellas son responsables en buena medida de la iniciación de los tubérculos. Tales hormonas se almacenan principalmente en las hojas viejas (Cranshaw y Radcliffe, 1980); en consecuencia, su eliminación puede inhibir dicha iniciación. En cuanto a los incrementos en rendimiento que se presentan al eliminar hojas del estrato medio, los mismos autores plantean que esta respuesta puede estar asociada a una optimización del Indice de Área Foliar (IAF), pues muchas hojas potencialmente eficientes desde el punto de vista fotosintético se encuentran en sectores de poca iluminación antes de aplicar la defoliación; luego de producirse ésta, la luz solar puede penetrar en mayor cantidad al follaje generándose un incremento en la fotosíntesis que conllevaría un mayor rendimiento.
Bagautdinova et al. (1976) citados por Dwelle (1985) encontraron que luego de una defoliación parcial la planta intensifica su tasa fotosintética en las hojas restantes. Rowe y Secor (1993) y Gudmestad (2008) dividieron el ciclo del cultivo de la papa en cinco estados de desarrollo principales, los cuales se presentan a continuación, pues se toman como base para la interpretación de resultados en esta investigación:
-
Siembra a emergencia: Previamente a la emergencia, las yemas y raíces se desarrollan a partir del tubérculo, el cual constituye el único suministro de energía para la planta.
-
Crecimiento vegetativo: Las yemas salen a la superficie y se desarrollan las hojas y tallos. Las raíces comienzan la toma de nutrientes desde el suelo. A partir de esta etapa, la planta es independiente de la semilla y la energía es suministrada por la fotosíntesis.
-
Iniciación de los tubérculos: Durante este periodo de tiempo se forman los pequeños tubérculos en la punta de los estolones, pero presentan un tamaño reducido. Coincide con el inicio de la floración.
-
Engrosamiento de los tubérculos: Poco tiempo después de la iniciación de los tubérculos comienza su engrosamiento. Durante esta etapa, ocurre la mayor extracción de nutrientes por parte de la planta y por esta razón se recomienda la aplicación de nitrógeno. El suministro adecuado de agua es esencial y las actividades de control de plagas y enfermedades deben realizarse cuidadosamente, pues es el momento de mayor susceptibilidad del cultivo. Esta etapa coincide con la floración plena. La producción de follaje finaliza y las hojas realizan su mayor actividad fotosintética con el fin de producir carbohidratos destinados al llenado de tubérculos y frutos.
-
Maduración de los tubérculos: en esta etapa el follaje envejece lentamente tornándose amarillo y secándose. La tasa fotosintética disminuye y los tubérculos crecen poco. Por último las plantas mueren y la epidermis del tubérculo se endurece.
La generación de información relacionada con los tópicos antes expuestos constituye un elemento importante para la determinación de las épocas críticas en las cuales se pueden estudiar los umbrales de daño económico. De esta forma se pueden generar recomendaciones de manejo del cultivo, que optimicen el potencial uso de los diferentes insumos.
El objetivo de este trabajo fue evaluar el efecto sobre el rendimiento de tubérculo, de diferentes niveles de defoliación en tres estados de desarrollo del cultivo en el cv. Parda Pastusa, bajo condiciones de páramo.
MATERIALES Y MÉTODOS
La fase experimental se realizó en el Centro Experimental ICA San Jorge en el municipio de Soacha (Cundinamarca, Colombia) a 3100 msnm, en un suelo FAr clasificado como humitropept ándico isomésico con características agroecológicas adecuadas para el cultivo. En el experimento se realizaron los controles fitosanitarios requeridos para garantizar la sanidad del cultivo.
Se utilizó un diseño completamente aleatorizado en condiciones de campo con estructura factorial 5x3. El primer factor correspondió a los porcentajes de defoliación (0, 20, 40, 60 y 80%), el segundo a las tres épocas de defoliación (90, 120 y 150 días después de la siembra (dds)). La unidad experimental consistió en un surco de 10 m de longitud, evaluando la totalidad de las plantas presentes en el mismo (aproximadamente 25). Se emplearon 5 repeticiones por tratamiento. Posteriormente, se calculó el rendimiento equivalente a 25 plantas, para cada uno de los valores obtenidos en campo y se procedió a realizar el análisis estadístico.
El número total de hojas por planta es altamente variable para un mismo estado de desarrollo del cultivo (Coeficiente de variación = 46,48%), por lo cual no resulta recomendable eliminar un número establecido de hojas en todas las plantas para asumir un porcentaje de defoliación determinado (Tabla 1). Adicionalmente, la eliminación de hojas completas implicaría asumir un área foliar constante para todas las hojas, lo cual no resulta válido. En consecuencia, se desarrolló un método en el cual se elimina el mismo porcentaje de área en cada una de las hojas de la planta. Para ello se requirió realizar un análisis previo de la distribución del área foliar teniendo en cuenta diferentes tamaños de hoja.
Tabla 1. Parámetros estadísticos para el número de hojas por planta de papa cv. Parda Pastusa a los 120 días después de la siembra.

Se tomaron 60 hojas de cada una de las categorías, provenientes de diez plantas (cinco de cada cv.) considerando este número de hojas como representativo. Se establecieron tres rangos de tamaño que permitieron agrupar las hojas en las categorías pequeña (longitud < 15 cm), mediana (longitud entre 15-20 cm) y grande (longitud > 20 cm). Se determinó el área foliar de la hoja completa y posteriormente se cortaron la totalidad de los foliolos presentes en cada hoja, registrando el área en centímetros cuadrados de cada uno de ellos. Se procedió a determinar el porcentaje de área que cada foliolo representa respecto al área foliar de la hoja completa y se estimaron los mismos parámetros estadísticos antes mencionados. Los foliolos se numeraron desde el ápice hasta la base (Figura 1).
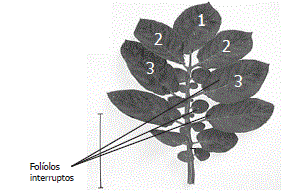
Figura 1. Numeración de los foliolos para la metodología de defoliación en plantas de papa cv. Parda Pastusa. Tabla 1. Parámetros estadísticos para el número de hojas por planta de papa cv. Parda Pastusa a los 120 días después de la siembra.
En cada uno de los tamaños se determinó el porcentaje de área que representaba cada foliolo respecto al área total de la hoja y se obtuvo un promedio e intervalo de confianza para cada tamaño (Tabla 2).
Tabla 2. Parámetros estadísticos para el número de hojas por planta de papa cv. Parda Pastusa a los 120 días después de la siembra.
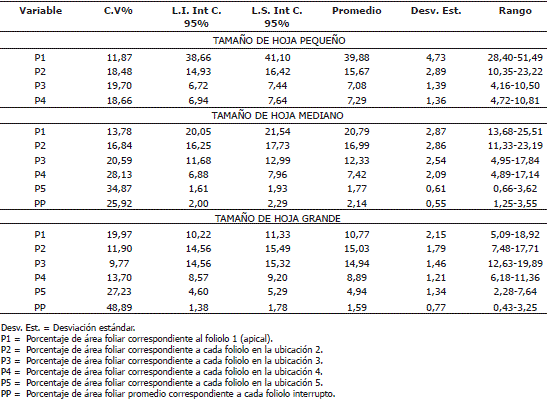
Con base en esta información se puede afirmar que la variabilidad en términos del área, de cada foliolo es mucho menor que la presente a nivel de número de hojas por planta. Adicionalmente, la estimación del área promedio por foliolo permite diseñar un patrón de defoliación que permita obtener los porcentajes de daño simulado deseados, de acuerdo al tamaño de la hoja, lo cual no es posible al eliminar hojas completas.
A partir de los resultados del procedimiento recién descrito se diseñó la forma de defoliación tomando en cuenta los tres tamaños de hoja antes mencionados. Esto se realizó con el fin de facilitar la aplicación de la metodología en el cultivo, pues se requiere que el operario que realiza la labor pueda rápidamente definir una categoría para la hoja que se dispone a cortar y porque de acuerdo al tamaño de hoja, el foliolo que ocupa una posición determinada corresponde a un porcentaje de área diferente.
El método desarrollado y empleado para lograr los diferentes niveles de defoliación en los distintos tamaños de hoja se relaciona en la Tabla 3.
Tabla 3. Procedimiento para la obtención de diferentes niveles de defoliación en distintos tamaños de hoja en la planta de papa cv. Parda Pastusa.
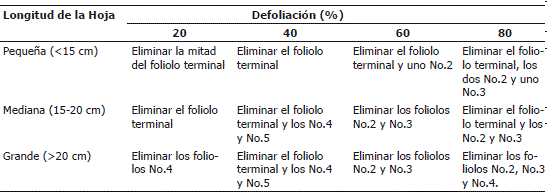
A manera de ejemplo en las Figura 2 se ilustra visualmente la metodología. La línea ubicada a la izquierda de las fotografías representa una longitud de 10 cm.
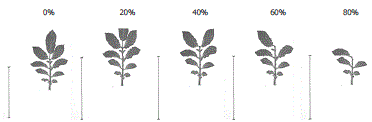
Figura 2. Metodología de defoliación en plantas de papa cv. Parda Pastusa para el tamaño de hoja pequeña.
En el momento de la cosecha los tubérculos fueron clasificados en categorías comercial y no comercial y se registró el peso correspondiente. El análisis de la información se efectuó mediante regresión múltiple, buscando obtener un modelo que permita estimar el rendimiento, por medio de una ecuación de la forma Y (rendimiento) = f (% defoliación, días después de siembra (época).
RESULTADOS
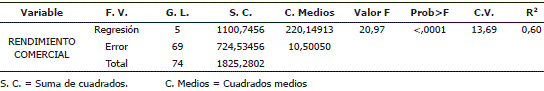
Se presentaron diferencias altamente significativas (P<0,01) en el rendimiento de tubérculo comercial para los dos factores evaluados (Porcentaje y época de defoliación) y para la interacción entre ellos (Tabla 4), demostrando que ambos afectan la producción del cv. Parda Pastusa. La existencia de interacción implica que el rendimiento de tubérculo comercial, en los diferentes porcentajes de defoliación, cambia dependiendo de la época en que ocurra el daño, de modo que un mismo porcentaje de defoliación puede no afectar el rendimiento en alguna época de desarrollo del cultivo, pero resultar altamente perjudicial en otro momento del desarrollo del mismo.
Tabla 4. Cuadrados medios del análisis de varianza para las variables evaluadas, en plantas de papa cv. Parda Pastusa sometidas a diferentes niveles de defoliación.

El análisis de varianza para el modelo indica que la regresión es altamente significativa (P<0,01), lo que permite afirmar que es idóneo para explicar el comportamiento de la variable dependiente. El coeficiente de determinación para la variable rendimiento comercial se puede considerar alto y adecuado (Tabla 5).
Tabla 5. Análisis de varianza para el modelo de regresión desarrollados para plantas de papa cv. Parda Pastusa sometidas a diferentes niveles de defoliación.
Rendimiento comercial. La regresión para esta variable corresponde al siguiente modelo:
Y = a + b1E + b2E2 + b3F + b4EF +ß4EF+ß5E2F Puesto que el modelo emplea la suma de cuadrados secuencial para la estimación de parámetros, debido al empleo de los polinomios ortogonales, se requiere incluir algunos pese a no ser estadísticamente significativos (Tabla 6). Por otra parte, se busca un modelo adecuado desde el punto de vista biológico, con el mayor sustento estadístico posible. Los parámetros de interacción (b4 y b5) entre días después de siembra (E) y porcentaje de defoliación (F) resaltan el hecho de que la respuesta al daño foliar en términos de rendimiento va a estar fuertemente condicionada por la época en que tal daño se presente. Esto concuerda con lo encontrado por varios autores; al respecto, Mailloux y Bostanian (1989), evaluando en Quebec el efecto de la defoliación producida por L. decimlineata en S. tuberosum cv. Kennebec que estaba próxima a la madurez, la defoliación solo reducía el rendimiento cuando alcanzaba niveles cercanos al 100% y se presentaba daño en los tallos, mientras que en la etapa de floración se produjeron pérdidas desde niveles de daño mucho menores, entre el 12 y 25%. Irigoyen, Domeño y Muro, (2010), evaluaron en España tratamientos de defoliación baja, media y alta en diferentes estados fenológicos de la planta, sobre los cv. Red Pontiac y Baraka. Estos autores encontraron que el daño producido durante la floración y la formación de tubérculos mermó considerablemente los rendimientos, pero no se presentaron reducciones importantes cuando los tubérculos estaban maduros.
Tabla 6. Parámetros estimados para la variable peso de tubérculos de clasificación comercial, en plantas de papa cv. Parda Pastusa sometidas a diferentes niveles de defoliación.
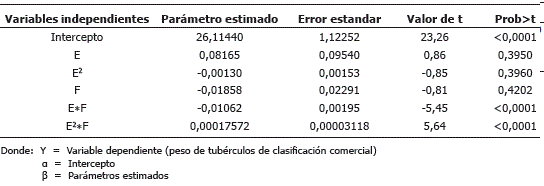
La interacción entre época de defoliación por porcentaje de defoliación (E*F) es negativa (Tabla 6), señalando que el rendimiento se reduce en forma lineal a medida que aumenta el porcentaje de defoliación, pero la pendiente de la recta varía por efecto de la época en la cual ocurre el daño, pues a los 120 días, coincidiendo con la etapa de floración y llenado de tubérculos o etapa IV según la escala de Gudmestad, (2008), esta pendiente es muy pronunciada, indicando que a esta edad la defoliación tiene un gran impacto en el rendimiento.
La interacción época al cuadrado por porcentaje de defoliación (E2*F) es positiva (Tabla 6) y se explica porque desde los 120 dds en adelante, el rendimiento se incrementa en forma cuadrática, a medida que transcurren los días alejándose de la época crítica. Por cada día que transcurre desde la época de defoliación de los 120 dds hasta la época de los 150 dds, se observa una disminución de la reducción del rendimiento (Figura 3). Esto puede explicarse porque a medida que avanza el desarrollo de la planta desde 120 días en adelante, los tubérculos van madurando y el efecto de la defoliación sobre la producción se reduce.
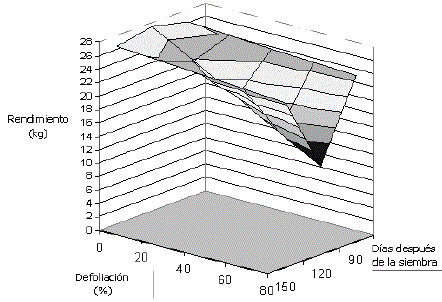
Figura 3. Superficie de respuesta para el rendimiento comercial en 25 plantas de papa cv. Parda Pastusa sometidas a diferentes niveles de defoliación.
DISCUSIÓN
El comportamiento observado en la variable evaluada (rendimiento comercial de tubérculo) se puede explicar con base en la fisiología y el desarrollo fenológico del cultivo.
A los 90 dds las plantas se encontraban en la fase de desarrollo III de la escala propuesta por Gudmestad (2008) y Rowe y Secor (1993), que corresponde al crecimiento vegetativo, junto con la inducción de tubérculos en la punta de los estolones, sin que aún se diera la presencia de flores. La defoliación producida en esta etapa tiene como efecto la estimulación de yemas aéreas que rápidamente se desarrollan al punto de compensar el daño en una forma aparentemente completa, con la generación de nuevas hojas. Adicionalmente, el estímulo de la defoliación puede generar un incremento en la tasa fotosintética en el área foliar restante, permitiendo una rápida recuperación. Puesto que el estado III (iniciación de tubérculos) no requiere una cantidad de fotoasimilados tan alta y la tasa fotosintética en esta etapa es baja (Rowe y Secor, 1993), es posible que la planta pueda superar el daño sin que haya una disminución significativa en el número de tubérculos inducidos. Por el contrario, en el estado IV la demanda de fotosintetizados es máxima, pero al llegar a esta etapa las plantas defoliadas a 90 dds ya se han recuperado del daño pues tienen la capacidad de generar nuevo follaje cuando el daño ocurre en estados tempranos (Radclifee, 2010) y por lo tanto están en capacidad de suplir la demanda para el llenado de los órganos subterráneos.
Dado que en sus fases iniciales de desarrollo las hojas se comportan más como un vertedero (Salisbury y Ross, 1992), es importante considerar que la eliminación de las hojas correspondientes al tamaño pequeño, no necesariamente implica la reducción de área fotosintéticamente activa, pues dependiendo de su estado de desarrollo, éstas pueden constituirse como fuentes, vertederos, o bien presentar un estado de transición.
La defoliación a los 120 dds coincide completamente con la floración y con la etapa de engrosamiento de los tubérculos. En esta etapa, la formación de follaje cesa y los fotoasimilados se destinan a los tubérculos. Hay dos posibles consecuencias del daño aplicado en este estado. En principio, al eliminarse las hojas la planta no dispone del área fotosintéticamente activa que le permita producir fotosintetizados en cantidad suficiente para un llenado adecuado de la totalidad de los tubérculos, por lo tanto, el peso total de tubérculos disminuye, así como el peso de tubérculos procesables. En relación con este planteamiento, Ziems et al., (2006) encontraron que la reducción en rendimiento en el cultivo de la papa como resultado de la defoliación se debe a que la capacidad de la planta para adquirir la cantidad de radiación necesaria se ve afectada cuando el porcentaje de intercepción lumínica cae por debajo del 90% durante la etapa crítica del llenado de los tubérculos. También existe la posibilidad de que la planta en respuesta al daño induzca la formación de nuevas hojas como un efecto de compensación lo cual implicaría que una parte de los productos de la fotosíntesis destinados inicialmente al engrosamiento de los tubérculos serían desviados para la formación del nuevo follaje. Ambos mecanismos podrían intervenir en la reducción del rendimiento que tiene lugar cuando se realiza el daño a los 120 dds.
La reducción del rendimiento de tubérculo comercial, no se da significativamente con el nivel de defoliación del 20%, aún en este periodo crítico (120 dds), lo cual posiblemente se debe a que la planta previamente a la defoliación presenta un valor alto de índice de área foliar (IAF), de manera que al realizar el daño en niveles bajos, la intercepción de luz no se reduce de manera importante. También Mailloux y Bostanian (1989) encontraron que durante la época de mayor susceptibilidad al daño, (la cual coincide con la floración y llenado de los tubérculos) en los cv. Superior y Kennebec hay un margen de tolerancia a la defoliación, que es de 12,5% en el primero y 25% en el segundo.
Debe además considerarse que al eliminar parte del follaje se puede producir un incremento en la tasa fotosintética de las hojas restantes y por lo tanto compensarse la pérdida de una parte del área fotosintéticamente activa, como lo reporta Pariera (2006) para S. tuberosum cv. Cal Red. La defoliación
a los 120 dds generó disminución significativa en el rendimiento comercial a partir del nivel de defoliación de 40%, reducción que alcanza su máximo en el nivel de 80% (Figura 3).
La defoliación aplicada a los 150 dds coincide con la etapa V de la escala de Rowe y Secor (1993) y Gudmestad (2008), la cual se caracteriza por la maduración de los tubérculos. También ocurre el envejecimiento del follaje y la muerte de la planta. En este estado, la translocación de fotoasimilados a los tubérculos es muy baja o ha concluido y la biomasa aérea por su estado de senescencia no está en capacidad de producir cantidades importantes de fotoasimilados, por lo tanto, el daño al follaje difícilmente podría tener consecuencias negativas en términos del rendimiento del cultivo. Por tales razones, la defoliación aplicada a los 150 dds no generó reducción importante en la producción. Esta información permite identificar la fase IV (Gudmestad, 2008), o de llenado de tubérculos y floración como la de mayor susceptibilidad por parte del cultivo al ataque por gota y demás plagas que afectan el follaje. En el cultivar evaluado bajo las condiciones de este ensayo, esta etapa se presenta aproximadamente a los 120 dds, por lo tanto, la protección del cultivo debe intensificarse en esta época para evitar pérdidas importantes en el rendimiento de tubérculo comercial.
En los estados iniciales de desarrollo, hasta 90 dds, la planta tiene capacidad de recuperarse de los daños producidos en el follaje por medio de la producción de nuevas hojas, por lo que no es muy probable que se afecten los rendimientos finales. De igual forma, el daño a los 150 dds o posteriormente, no tiene un efecto importante en el rendimiento, sin importar la gravedad del daño a que se someta el follaje, y por esto no conviene hacer énfasis en la protección de la parte aérea del cultivo en esta época, pues sería incurrir en gastos innecesarios, aunque debe tenerse presente que la probabilidad de infección en los tubérculos podría aumentar.
Para la realización de estudios posteriores, dependiendo de los objetivos del trabajo, puede resultar conveniente llevar a cabo mediciones de área foliar y de IAF en las plantas sometidas a la defoliación. Esta información permitiría establecer la capacidad de recuperación de la planta y la respuesta en términos de la redistribución de fotoasimilados que ocurre posteriormente a la aplicación del daño.
AGRADECIMIENTOS
Al Centro Experimental ICA San Jorge por las facilidades de infraestructura para desarrollar la investigación y, a los hoy colegas Ingenieros Agrónomos: Herlinda Álvarez, Rodrigo Muñoz, Alexander Perilla, Oscar Darío Ramírez, Mary Luz Ruiz y Helman Sandoval, quienes colaboraron con el trabajo de defoliaciones.
BIBLIOGRAFÍA
Cranshaw, W.S. and E.B. Radcliffe. 1980. Effect of defoliation on yield of potatoes. Journal of Economic Entomology 73(1): 131-134. [ Links ]
Dwelle, R. 1985. Photosynthesis and photoassimilate partitioning. p. 35-58. In: Li, P.H. (ed.). Potato Phisiology. Academic Press, Florida. Gudmestad, N. 2008. Potato health from sprouting to harvest. p. 67-77. In: Johnson, D.A. (ed.). Potato health management. Second Edition. American Phytopathological Society, E. U. A. 272 p. [ Links ]
Hare, J.D. 1980. Impact of defoliation by the Colorado potato beetle on potato yields. Journal of Economic Entomology 73(3): 369-373. [ Links ]
Irigoyen, I., I. Domeño and J. Muro. 2010. Effect of defoliation by simulate hail damage on yield of potato cultivars with different maturity performed in Spain. American Journal of Potato Research 69: 705-714. [ Links ]
Mailloux, G. and N.J. Bostanian. 1989. Effect of manual defoliation on potato yield at maximum abundance of different stages of Colorado potato beetle Leptinotarsa decimlineata (Say) in the field. Journal of Agricultural Entomology 6(4): 217-226. [ Links ]
Metcalf, R. and W. Luckmann, 1982. Introduction to insect pest management. Second Edition. Wiley Intescience, New York. 650 p. [ Links ]
Midmore, D.J. 1981. Respuesta de la planta de papa (Solanum spp.) al daño de insectos: algunos efectos de compensación. p. 176-200. En: Memorias. Curso sobre Control Integrado de Plagas de Papa. CIP-ICA, Bogotá. [ Links ]
Pareira, C. 2006. Photosynthetic and glycoalkaloid responses of potato (Solanum tuberosum L.) to Colorado potato beetle (Leptinotarsa decimlineata Say) defoliation. Thesis Magister Scientiae in Entomology, Montana State University, 113 p. [ Links ]
Robert, Y. 1999. Insectos de la patata. En: Rousselle P., Y. Robert and J.C. Crosnier (eds.). La patata. Ediciones Mundi-Prensa, Madrid, España. 603 p. [ Links ]
Rowe, R. and G. Secor. 1993. Managing potato health from emergence to harvest. p. 35-40. In: Rowe, R.C. (ed.). Potato health management. Randall Rowe, New York. 272 p. [ Links ]
Salisbury, F.B. and C.W. Ross 1992. Plant phisiology. Fourth Edition. Wadsworth Publishing Co., Belmont, CA. 682 p. [ Links ]
Radcliffe, E.B. 2010. Insect control. p. 64-66. In: Bohl, W. and Johnson, S. (eds.). Commercial potato production in North America. The Potato Association of America, New York. 90 p. [ Links ]
Urquijo, P. y P. Landaluze. 1961. Patología vegetal agrícola. Segunda Edición. Editorial Salvat, Barcelona. 755 p. [ Links ]
Wellik, M.J., J.E. Slosser, and R.D. Kirby. 1981. Effects of simulated insect defoliation on potatoes. American Potato Journal 58(12): 627-632. [ Links ]
Zapata, J.L. y O. Trillos. 1997. Manejo integrado de la gota de la papa P. infestans (Mont) de Bary. p.87-92. En: Memorias. IV Foro de Sanidad Vegetal y I Seminario Nacional sobre Gota de la Papa. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Medellín. [ Links ]
Ziems, J., W. Zechmann, W. Hoback, J. Wallace, R. Madsen, T. Hunt and L. Higley. 2006. Yield response of indeterminate potato (Solanum tuberosum L.) to simulated insecto defoliation. Agronomy Journal 98(6): 1435-1441. [ Links ]

