










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.2 Medellín July/Dec. 2010
ALGUNOS FACTORES RELACIONADOS CON LA DINÁMICA FOLICULAR EN BOS INDICUS
SOME FACTORS RELATED TO FOLLICULAR DYNAMICS IN BOS INDICUS
Guillermo Henao Restrepo1
1 Profesor Titular. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779. Medellín, Colombia. <ghenao@unal.edu.co>
Recibido: Enero 22 de 2010; Aceptado: Octubre 13 de 2010
Resumen. El desarrollo folicular bovino durante un ciclo estral normal, se caracteriza por un crecimiento en forma de ondas con presencia de dos a cinco cohortes foliculares por ciclo, de las cuales solo un folículo se torna ovulatorio. El crecimiento folicular en forma de ondas también se produce durante el período prepuberal, puberal, primer tercio de la gestación y período de anestro posparto. Aunque existe mucha similitud en el patrón fundamental del desarrollo de ondas foliculares entre Bos taurus y Bos indicus, se han encontrado diferencias en la dinámica folicular que pueden afectar el comportamiento reproductivo y la aplicación de biotecnologías reproductivas. La dinámica folicular puede variar por efectos ambientales y estados fisiológicos de hembras Bos indicus que impiden establecer un patrón específico de dinámica folicular para cada raza y etapa fisiológica. El propósito de esta revisión es analizar la función reproductiva teniendo en cuenta las diferencias de la dinámica folicular entre estados fisiológicos y condiciones ambientales de hembras bovinas con énfasis en B. indicus.
Palabras clave: Cebú, ultrasonografía, reproducción animal, biotecnología reproductiva.
Abstract. The bovine follicular development during a normal estrous cycle, is characterized by a growth in wave form with presence of two to five follicular cohorts by cycle, of which only one follicle ovulate. The follicular growth in wave form also occurs during the prepubertal and pubertal periods, first quarter of the gestation, and in the postpartum anestrous period. Although there is great similarity in the fundamental pattern of the development of follicular waves between Bos taurus and Bos indicus, differences of follicular dynamics has been detected, that can affect the reproductive behavior and the application of reproductive biotechnologies. Follicular dynamics can change by environmental effects and physiological states of Bos indicus females that impede to establish a specific pattern of follicular dynamics for each breed and physiological condition. The purpose of this review is to analize the reproductive function, considering the differences in the follicular dynamics between physiological states and environmental conditions of bovine females with emphasis in B. indicus.
Key words: Zebu, ultrasounography, animal reproduction, biotechnology reproduction.
Con el apoyo de técnicas de biología molecular, ultrasonografía, radioinmunoanálisis, inmunohistoquímica y otras, ha sido posible realizar estudios importantes sobre las variaciones que puede presentar la dinámica folicular en ganado Bos taurus y Bos indicus, encontrándose diferencias representativas entre y dentro de especies, ocasionadas entre otros, por efectos genéticos y ambientales. El propósito de esta revisión es analizar la función reproductiva teniendo en cuenta las diferencias de la dinámica folicular entre estados fisiológicos y condiciones ambientales de hembras B. indicus, con el fin de favorecer la aplicación de biotecnologías reproductivas a los animales de valor genético y comercial importante para obtener la mayor cantidad de descendientes en el menor tiempo posible.
Dinámica folicular. La dinámica folicular ovárica bovina ha sido muy estudiada a través de diferentes técnicas, siendo las más aplicadas la ultrasonografía y la medición de niveles hormonales, mostrando variaciones que dependen de la genética, el estado fisiológico, el clima, la nutrición o el sistema de producción. El estudio de la dinámica folicular tiene como finalidad mejorar el conocimiento de los principios fisiológicos que rigen la producción de óvulos y de hormonas ováricas y su relación con el comportamiento reproductivo.
La teoría de crecimiento folicular en forma de ondas fue propuesta originalmente por Rajakoski (1960), quien basado en observaciones visuales e histológicas, propuso que el crecimiento de los folículos ováricos se desarrollaba en forma de ondas y que durante un ciclo estral se propagaban dos ondas foliculares (OFs). Esta teoría fue puesta en duda por más de 20 años durante los cuales se realizaron múltiples estudios con resultados contradictorios sobre la naturaleza del crecimiento folicular, variando entre ausencia de OFs y la presencia de tres o cuatro por ciclo estral (Adams et al., 2008), hasta cuando Pearson y Ginther (1984) realizaron el primer estudio ultrasonográfico de ovarios bovinos, determinando la presencia de OFs durante el ciclo estral. Una serie variada de investigaciones realizadas en las dos décadas siguientes (Spicer et al., 1986; Short, 1990; Savio et al., 1990; Murphy, Boland y Roche, 1990; Taylor y Rajamahendran, 1991; Lucy et al., 1992; Adams et al., 1992; Adams, 1994; Fortune, 1994; Ginther et al., 1996; Ginther et al., 1997; Monniaux et al., 1997; Webb et al., 1999) estudiaron el crecimiento y desarrollo de los folículos (por ultrasonografía) y midieron receptores (por inmunohistoquímica) y niveles hormonales (por radioinmunoanálisis) con el propósito de mejorar la comprensión de la función ovárica. Aunque se ha logrado un gran avance en el conocimiento de la fisiología ovárica aun se desconocen muchos eventos relacionados con el crecimiento y desarrollo de los folículos.
El desarrollo de OFs se produce durante los períodos prepuberal (Adams, Evans y Rawling, 1994; Calderón et al., 2000; Reis et al., 2005), puberal (Sunderland et al., 1996; Figueiredo et al., 1997; Calderón et al., 2000), anestro posparto (Murphy, Boland y Roche, 1990; Henao, 1998; Henao, Trujillo y Vásquez, 2002), primeros meses de gestación (Sunderland et al., 1996; Monniaux et al., 1997; Henao y Trujillo, 2003) y ciclo estral (Murphy, Boland y Roche, 1990; Savio et al., 1990; Henao, 1998).
Dinámica folicular durante el ciclo estral. Durante un ciclo estral las vacas Brahman pueden desarrollar una (6,7%), dos (90,0%) o tres (3,3%) OFs (Roa et al., 2006), mientras que las vacas del mismo grupo racial estudiadas por Zeitoun, Rodriguez y Randel (1996) presentaron dos (38%), tres (56%) o cuatro (9%) OFs por ciclo; sin embargo, en las vacas Cebú alojadas bajo condiciones de trópico colombiano se encontraron entre dos y seis OFs por ciclo (Ruiz y Olivera, 1999). La variación en el número de OFs también fue observada en hembras Gyr multíparas que desarrollaron dos (6,6%), tres (60,0%), cuatro (26,6%) o cinco (6,6%) OFs por ciclo (Moreira et al., 2000); en vacas Cebú mestizas durante el primer ciclo estral posparto que formaron una (16,5%), dos (67%) o tres (16,5%) OFs por ciclo (Perea et al., 1998); en vacas Guzerá multíparas con dos (37,5%), tres (50%) o cuatro (12,5%) OFs por ciclo (Coutinho et al., 2007) y en las vacas Nelore que presentaron dos (83,3%) o tres (16,7%) OFs por ciclo, con un índice de constancia de 0,7 (Figueiredo et al., 1997).
La primera OF de vacas Gyr emergió el día 1,2±0,5 del ciclo, la segunda el día 7,2±0.9, la tercera el día 11,2±0,9 y la cuarta el día 15±1,4 (Moreira et al., 2000). En vacas Nelore que presentaron tres OFs, la emergencia de la primera, segunda y tercera ondas ocurrió los días 1,5±0,2, 9,1±0,4 y 15,1±0,4 del ciclo (Figueiredo et al., 1997). En la raza Gyr la divergencia entre el FD y el FS de la primera, segunda, tercera y cuarta ondas, fue observada los días 5,7±1,7; 10,2±0,9; 14,7±0,9 y 18,7±1,2 del ciclo, respectivamente (Moreira et al., 2000); sin embargo, Sartorelli et al. (2005) mediante ultrasonografías ovárica de vacas Nelore no lactantes, realizada cada 12 horas, determinaron que la desviación folicular ocurrió el día 2,4±0,8 de la onda.
De acuerdo con el número de OFs observadas durante el ciclo estral en zona tropical, una (6,7%), dos (90,0%) y tres (3,3%), la duración del ciclo estral fue de 20,6±0,5; 22,3±2,2 y 27,0±1,5 días, respectivamente (Roa et al., 2006) y en zona templada la duración fue de 20,3±0,4 y 20,8±0,3 días para vacas Brahman con dos o tres ondas por ciclo, respectivamente (Zeitoun, Rodriguez y Randel, 1996). En vacas Nelore que desarrollaron dos y tres ondas por ciclo estral, su duración fue de 20,6±0,4 y 22,0±0,4 días, respectivamente (Figueiredo et al., 1997) y en Guzerá con tres OFs, de 19,8±2,4 días. El seguimiento ultrasonográfico ovárico durante el primer ciclo estral posparto de vacas Cebú mestizas determinó la duración del ciclo estral en 14, 20 y 23 días para hembras que desarrollaron una, dos o tres OFs por ciclo, respectivamente (Perea et al., 1998). Algunas vacas con dos o tres ondas de crecimiento folicular por ciclo presentan similar duración del ciclo estral (Zeitoun, Rodriguez y Randel, 1996), lo que indica que otros factores diferentes a la vida media del cuerpo lúteo pueden determinar la duración del ciclo estral en ganado B. indicus.
Dinámica folicular durante el período prepuberal. Durante el período prepuberal de B. indicus se desarrollan OFs similares a las que se presentan durante esta etapa en B. taurus. En novillas Cebú prepúberes se encontraron OFs con folículo dominante (FD) de 11,1±0,02 mm de diámetro (Ø) y una duración de 7,3±0,1 días (Calderón et al., 2000). Las novillas Gyr prepúberes con 22±2 meses de edad y 252±25 kg de peso vivo estudiadas por Reis et al., (2005), presentaron desarrollo de OFs con crecimiento y regresión de FDs y de un número variable de folículos subordinados (FSs). Se observó una duración variable de las OFs y por esta razón fueron agrupadas en OFs ≤12 días (12,2%), de 13 a 16 días (63,3%) y ≥17 días (24,4%).
Dinámica folicular durante el período puberal. Durante la pubertad de novillas Brahman se presentan dos (26,5%), tres (66,7%) o cuatro (6,8%) OFs por ciclo (Rhodes, D´Ath y Entwistle, 1995). La emergencia del FD de la segunda onda ocurrió los días 11,8±0,4; 10,1±0,2 y 9,1±0,8 y la del folículo ovulatorio, los días 11,8±0,4; 15,8±0,3 y 18,9±0,9 en novillas de dos, tres y cuatro OFs, respectivamente. Un comportamiento similar presentaron las novillas Nelore estudiadas por Figueiredo et al., (1997) y Cebú estudiadas por Calderón et al., (2000), en las cuales se observaron dos (29,4% y 32%), tres (64,7% y 64%) o cuatro (5,8% y 4%) OFs por ciclo; el índice de constancia del número de OFs entre el primero y el segundo ciclo estral puberal fue de 0,2±0,05, indicando un cambio fisiológico entre el primero y el segundo ciclo (Calderón et al., 2000), lo que impide predecir el número de OFs de los ciclos siguientes.
En novillas Holstein-Cebú se encontró una predominancia de tres (58,3%) frente a dos (33,3%), una (6%) y cuatro (3%) OFs por ciclo. El inicio de las OFs fue detectado los días 0,4 y 9,7 para los ciclos con dos ondas y en los días 0,4; 8,2 y 15,9 para los ciclos con tres ondas (Borges et al., 2001). El índice de constancia del número de OFs por ciclo en este cruce fue de 0,72.
En novillas Brahman los FDs alcanzaron un Ø máximo de 10,2±0,1 mm (Rhodes, D´Ath y Entwistle, 1995) y en novillas Nelore, 11,4 mm (Sartorelli et al., 2005), menores (13,3±1,4 mm) que los de novillas Holstein-Cebú (Borges et al., 2001). Cuando se sincronizó la ovulación de novillas Nelore, el folículo ovulatorio alcanzó un Ø de 8,1±0,2 mm (Trevizoli, 2007), talla menor que la de novillas del mismo grupo racial (11 mm) sincronizadas por Figueiredo et al., (1997).
En novillas Brahman el ciclo estral tuvo una duración de 20,9±0,1 días, presentándose diferencia entre novillas con desigual número de OFs por ciclo. En las novillas que presentaron dos, tres y cuatro OFs, el ciclo tuvo una duración de 20,2±0,3; 21±0,2 y 22±0,67 días, respectivamente (Rhodes, D´Ath y Entwistle, 1995). La duración del ciclo estral en novillas Holstein-Cebú también dependió de la presencia de dos (20,6±1,2 días) o tres (23,0±1,3 días) ondas por ciclo (Borges et al., 2001). La duración del cuerpo lúteo se incrementa conforme aumenta el número de ondas por ciclo: 18,1±0,4; 18,9±0,2 y 20,0±0,7 días en novillas de dos, tres y cuatro OFs, respectivamente (Rhodes, D´Ath y Entwistle, 1995). La presencia de una cuarta onda folicular en B. indicus constituye una diferencia con respecto a B. taurus, especie en la que es escaso su reporte. Esta diferencia fisiológica puede tener implicaciones para el éxito de tratamientos de sincronización de estros, superestimulación ovárica y aspiración transvaginal de oocitos en ganado B. indicus.
Tasa de crecimiento folicular. La tasa de crecimiento de los FDs en vacas Brahman en clima tropical seco fue de 0,9±0,2 mm/día (Henao, Olivera y Maldonado, 2000), mientras que en vacas Guzerá la tasa de crecimiento para los FDs de la primera, segunda y tercera OFs fue de 1,3±0,09; 0,8±0,08 y 1,0±0,1 mm/día, respectivamente (Coutinho et al., 2007), similar a la de vacas Nelore, 0,8±0,1; 0,7±0,08 y 1,0±0,07 mm/día (Figueiredo et al., 1997) y Gyr 1,0±0,2; 1,0±0,2 y 1,0±0,2 (Moreira et al., 2000).
Diámetro máximo de folículos. El Ø máximo de los FDs en el ganado B. indicus es menor que el que se presenta en B. taurus y esto parece obedecer a una respuesta genética. Las vacas Brahman estudiadas por Rhodes, D´Ath y Entwistle (1995) desarrollaron FDs con Ø máximo de 10,2±0,1 mm, mientras que las estudiadas por Henao(1998); Henao, Olivera y Maldonado (2000); Henao, Trujillo y Vásquez (2000); Henao y Trujillo (2003); Henao y González (2008), desarrollaron un Ø preovulatorio de 10 a 14 mm. En vacas Nelore que desarrollan tres OFs el Ø del folículo ovulatorio (11,6±0,2 mm) fue mayor que el de la primera y segunda ondas (10,4±0,2 y 9,3±0,3 mm, respectivamente); en las que desarrollan dos ondas el FD de la segunda alcanzó mayor Ø (12,0±0,2 mm) que el de la primera (11,3±0,3 mm, Figueiredo et al., 1997); una talla similar alcanzaron los FDs de vacas Gyr (Moreira et al., 2000; Moreira et al., 2004; Sartorelli et al., 2005) y Nelore (Coutinho et al,. 2007; Carvalho et al., 2008; Machado et al., 2008). En vacas Cebú anéstricas el FD obtuvo un Ø de 11,7±2,4 mm (De Ondiz et al., 2002), diferente al de vacas Cebú (16,1±1,3 mm) alojadas bajo condiciones tropicales (Gyraldo et al., 2005) y al de vacas Gyr (13,3±0,8 mm) sometidas a estrés térmico prolongado (Torres et al., 2008). El Ø máximo alcanzado en vacas Gyr por los FSs fue de 7,2±0,7 mm (Moreira et al., 2000), similar a los de vacas Nelore (Figueiredo et al., 1997; Coutinho et al., 2007) y Brahman (Henao, Olivera, Maldonado, 2000; Henao, Trujillo y Vásquez, 2002).
Número de folículos por cohorte. El número de folículos >2 mm reclutados por cohorte en novillas Nelore y Gyr fue de 33,4±3,2 (Carvalho et al., 2008), similar a 39±4 reportado por Álvarez et. al., (2000) y mayor que el de las cohortes de ganado B. taurus X B. indicus (29,6±2,5) y de B. taurus (25,4±2,5) (Carvalho et al., 2008). En las OFs presentadas durante los primeros 60 días posparto de vacas Brahman primerizas, Henao y González (2008) encontraron 15±7,8 folículos >3mm por cohorte.
Dinámica folicular durante la gestación y el período posparto en vacas multíparas. Durante los primeros tres meses de la gestación bovina, los ovarios continúan desarrollando OFs sucesivas con atresia del FD (Figura 1). En la primera onda folicular formada después de la concepción se desarrolla un FD de Ø similar a un folículo ovulatorio, pero los FDs de ondas sucesivas disminuyen su Ø (Ginther, Kastelic y Knopf, 1989; Henao y Trujillo, 2003). Durante el último tercio de la gestación continúa el crecimiento de folículos antrales, pero éstos no alcanzan el estado de madurez (Rexroad y Casida, 1975).
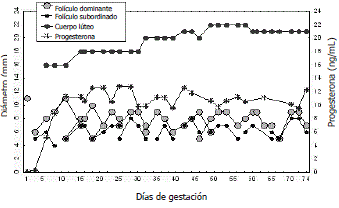
Figura 1. Dinámica folicular y lútea durante los primeros 74 días de gestación de una vaca Brahman (Henao y Trujillo, 2003).
A partir de la primera semana posparto la secreción temprana de la hormona folículo estimulante (FSH) promueve el reclutamiento de la primera cohorte folicular. Una alta proporción de ganado B. taurus presenta ovulación a partir del FD de la primera onda posparto (Savio et al., 1990), pero no ocurre lo mismo en ganado B. indicus debido a los patrones diferentes de secreción de hormona luteinizante (LH) con respecto a B. taurus. Las pulsaciones de LH a los 30 días posparto fueron inferiores en B. indicus que en B. taurus (D’occhio, Neish y Broadhurst, 1990).
Las deficiencias nutricionales y los efectos negativos del clima son frecuentes en el trópico colombiano, en el que algunas vacas Cebú estudiadas por Ruiz y Olivera (1999) no presentaron OFs durante los primeros días del período posparto y tardaron entre 26 y 78 días para iniciar la formación de la primera OF posparto. Durante los primeros seis meses estas vacas presentaron períodos prolongados en los que se observó desarrollo de folículos que alcanzaron un Ø máximo de 6 mm. Entre siete y doce meses posparto se observaron cambios en la dinámica folicular que condujeron a la primera ovulación, ocurrida entre 217 y 278 días posparto (249±12). La dinámica folicular del anterior grupo de vacas Cebú en amamantamiento presentó un comportamiento diferente al de las vacas Cebú a las que se retiró el ternero desde el día del parto, en las cuales la primera OF se inició ente seis y 10 días posparto (Toribio et al., 1995).
La dinámica folicular durante los primeros 60 días postparto de cinco vacas Brahman amamantando (Henao, Trujillo y Vásquez, 2002) se caracterizó por un desarrollo precoz de OFs conformadas por cohortes de 6,±0,4 folículos ≥ 4mm de Ø. La primera OF se empezó a observar desde el día seis posparto y estuvo conformada por 4,6±0,5 folículos ≥4 mm, de los cuales se formó un FD que alcanzó un Ø máximo de 10,4±0,5 mm a los 12,8±2,0 días posparto. De las cinco vacas evaluadas, tres presentaron la primera ovulación posparto a los 34,4±12,3 días sin signos de estro (estro silencioso), una vaca ovuló con el FD de la segunda OF y dos con el de la cuarta. El primer folículo ovulatorio midió 12,0±0,8 mm de Ø y fue semejante (P>0,05) al promedio de los Øs de los demás FDs. De las tres vacas que ovularon, dos presentaron una segunda ovulación, acompañada de signos de estro antes de los 60 días posparto y tuvieron un intervalo interovulatorio de 19±5 días y un intervalo parto primer estro de 49±8 días. Después de la primera ovulación se formó un cuerpo lúteo con un Ø máximo de 16,7±1,3 mm, una vida media de 11,3±3,8 días y un nivel máximo de 1,0±0,3 ng/ml de progesterona.
En zona tropical se evaluaron 11 vacas Brahman amamantando (Figura 2) y cinco sin amamantar (Figura 3) a partir del día 30±8 posparto hasta el día 140, tiempo durante el cual todas las vacas sin amamantamiento y siete amamantando presentaron al menos dos estros (Henao, Olivera y Maldonado, 2000). Se encontraron OFs con presencia de folículos dominantes que alcanzaron Øs máximos de 11,2±0,4 y de 10,9±0,5 mm (P>0,05), respectivamente, y subordinados de 7,3±0,4 y de 6,8±0,2 mm (P>0,05), respectivamente.
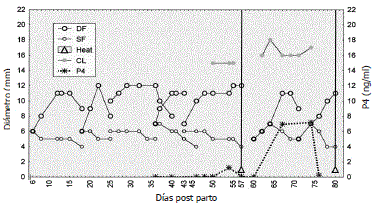
Figura 2. Dinámica folicular posparto de una vaca Brahman amamantando. DF: Folículo dominante. SF: Folículo subordinado. Cl: Cuerpo lúteo. P4: progesterona (Henao, Olivera y Maldonado, 2000).
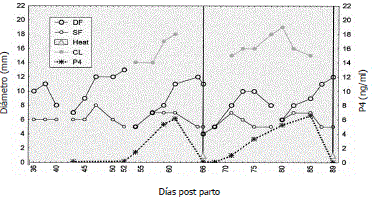
Figura 3. Dinámica folicular posparto de una vaca Brahman sin amamantamiento. DF: Folículo dominante. SF: Folículo subordinado. CL: Cuerpo lúteo. P4: progesterona (Henao, Olivera y Maldonado, 2000).
El amamantamiento afectó el tiempo a la primera ovulación y al primer estro posparto, siendo menor en las vacas sin amamantamiento. El 85,7% y el 40% de las vacas con y sin amamantamiento, respectivamente, presentó la primera ovulación posparto sin estar precedida de signos de estro. Durante el primer ciclo estral posparto las vacas Brahman sin amamantamiento presentaron una (40%) o dos (60%) OFs y las que amamantaban, dos (71,4%) o tres (28,5%).
Cuando las vacas Brahman amamantando que presentan anestro posparto prolongado se someten a supresión temporal del amamantamiento, ocurren cambios en su dinámica folicular que conducen a la presentación de la ovulación y terminación del anestro posparto (Figura 4) o a un incremento en el desarrollo del folículo dominante que posteriormente desarrolla atresia (Henao, Trujillo y Vásquez, 2000). Al comparar los Øs de los FDs antes de la suspensión temporal de la lactancia de vacas que ovularon (11,2±1,5 mm) con los de las que no ovularon (10,0±1,8 mm) se encontró una diferencia significativa (P<0,05). Estos resultados concuerdan con los de Giraldo et al., (2005) quienes reportaron la presencia de FDs de mayor Ø en vacas Cebú que ovularon con respecto a las que no ovularon después de una suspensión temporal de la lactancia por 72 horas. Cuando Mahecha et al. (2003) compararon los Øs de los FDs de vacas Cebú ciclando sometidas a interrupción temporal del amamantamiento con los de un grupo de vacas control ciclando en amamantamiento permanente, encontraron un Ø superior en las primeras.
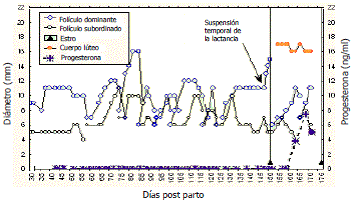
Figura 4. Dinámica folicular posparto de una vaca Brahman amamantando que presentó anestro posparto prolongado (Henao, Trujillo y Vásquez, 2000).
Período posparto de vacas primerizas. De 20 vacas Brahman primerizas en amamantamiento estudiadas por Henao y González, (2008) durante los primeros 60 días del período posparto, 18 desarrollaron OFs a partir de la primera semana posparto, una a partir de la segunda y otra a partir de la tercera. Durante los primeros 60 días posparto, en cada vaca se presentaron 7,6±1,9 OFs. En cada OF el número promedio de folículos ≥3mm fue 15±7,8. Los FDs crecieron a una tasa de 1,3±0,4 mm/día, alcanzaron un Ø de 10,7±1,7 mm y presentaron un intervalo interdominancia de 7,2±3,5 días. Los FSs alcanzaron un Ø máximo de 7,03±1,4 mm. En una vaca el FD de la segunda onda se tornó persistente, con una fase de meseta que duró 11 días; el FD de la tercera onda se tornó quístico, con una duración de 27 días y un Ø máximo de 19 mm. Durante el tiempo de la existencia del quiste folicular se desarrollaron dos OFs con FDs que alcanzaron Øs e intervalos interdominancia superiores a los de las compañeras. Las medidas de este quiste fueron inferiores a las que alcanzan las vacas B. taurus, en las cuales se considera como quística una estructura folicular que crece más de 20 mm y persiste por más de 10 días (Peter, 2004). Esta es la primera vez que se describe el estudio ultrasonográfico de un caso de quistosis folicular en vacas Brahman en Colombia (Figura 5. Datos no publicados).
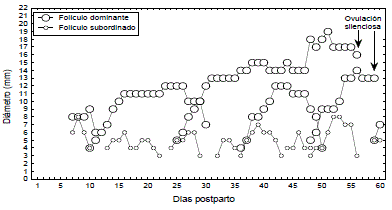
Figura 5. Representación de la dinámica folicular posparto de una vaca Brahman que desarrolló folículo persistente a partir de la segunda onda folicular y quiste folicular a partir del folículo dominante de la tercera onda (Información no publicada).
Las vacas Cebú primerizas presentan un anestro posparto más prolongado que las multíparas, debido a que el consumo de nutrientes, además de tener que suplir los requerimientos para mantenimiento de funciones fisiológicas y de lactación, debe suplir las necesidades que exige su etapa final de crecimiento antes de reiniciar su actividad cíclica reproductiva (Soto et al., 2001; Plasse, 1994). Esta condición fisiológica, unida a las alteraciones de salud que causan los parásitos internos y externos, al estrés ocasionado por las inclemencias climáticas tropicales y a la deficiente administración del ganado en sistemas extensivos de producción, justifican la realización de más investigaciones que favorezcan el esclarecimiento de la función ovárica durante el posparto de vacas Cebú primerizas.
CONCLUSIONES
Las vacas B. indicus presentan un número variable de OFs por ciclo estral y es más frecuente encontrar ciclos de tres OFs que en B. taurus. Las causas de variabilidad no han sido estudiadas y no se conoce con precisión el efecto que puedan tener sobre la eficiencia de la fertilidad.
El ganado B. indicus presenta un menor diámetro de los folículos dominantes y del cuerpo lúteo que el B. taurus, lo cual parece ser una característica genética. Se presenta variación en los diámetros de los folículos entre vacas del mismo grupo racial o entre diferentes razas y en varios estados fisiológicos. Las causas de estas variaciones no han sido estudiadas y podrían tener consecuencias sobre la fertilidad y la eficiencia de los protocolos de superovulación, maduración y fertilización in vitro.
El número de folículos reclutados en las cohortes foliculares es mayor en vacas y novillas B. indicus que en B. taurus. Las vacas B. indicus tienen intervalos más reducidos desde el celo hasta la secreción de LH y desde la secreción de LH hasta la ovulación que las B. taurus.
Existe una alta variación en el comportamiento del crecimiento y desarrollo de los folículos, influenciada por efectos genéticos y ambientales de diversa índole, que impiden establecer un patrón específico de dinámica folicular para B. indicus en cada etapa fisiológica. Esta situación motiva la realización de más estudios que involucren diferentes condiciones fisiológicas y aspectos ambientales en los que la función ovárica puede alterarse.
BIBLIOGRAFÍA
Adams, G.P., R.L. Matteri, J.P. Kastelic, J.C.H. Ko and O.J. Ginther 1992. Association between surges of follicle-stimulating hormone and the emergence of follicular waves in heifers. Journal of Reproduction and Fertility 94(1): 177-188. [ Links ]
Adams, G.P. 1994. Control of ovarian follicular wave dynamics in cattle: Implications for synchronization and superstimulation. Theriogenology 41: 19-24. [ Links ]
Adams, G.P., A.C.O. Evans and N.G. Rawlings. 1994. Follicular waves and circulating gonadotropins in 8-months-old prepubertal heifers. Journal of Reproduction and Fertility 100(1): 27-33. [ Links ]
Adams, G.P., R. Jaiswal, Singh, J and P. Malhi. 2008. Progress in understanding ovarian follicular dynamics in cattle. Theriogenology 69: 72-80. [ Links ]
Alvarez, P., L.J. Spicer, C.C. Chase Jr., M.E. M. E. Payton, T.D. Hamilton, R.E. Stewart, A.C. Hammond, T.A. Olson and R.P. Wettemann 2000. Ovarian and endocrine characteristics during an estrous cicle in Angus, Brahman and Senepol cows in a subtropical environment. Journal of Animal Science 78(5): 1291-1302. [ Links ]
Borges, A.M., C. Torres, J.R.M. Ruas, V.R. Rocha Júnior, G.R. and Carvalho. 2001. Dinâmica folicular ovariana em novilhas mestiças Holandês-Zebu. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 53(5): 595-604. [ Links ]
Calderón, R., A. Villa-Godoy, L. Lagunaes y P. Fajersson. 2000. Desarrollo folicular en novillas Cebú y Suizo Pardo en condiciones tropicales. Técnica Pecuaria México 38: 163-175. [ Links ]
Calvalho, J.B.P., N.A.T. Carvalho, E.L. Reis, M. Nichi, A.H. Souza and P.S. Baruselli. 2008. Effect of early luteolysis in progesterone-based times AI protocols in Bos indicus, Bos indicus x Bos taurus, and Bos taurus heifers. Theriogenology 69(2): 167-175. [ Links ]
G.T.R.M. Coutinho, J.H.M. Viana, W.F. Sá, L.S. Camargo, A.M. Ferreira, P.M. Palhão e L.A.G. Nogueira. 2007. Avaliação ultra-sonográfica da dinâmica folicular e lútea em vacas da raça Guzerá. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 59(5): 1089-1096. [ Links ]
De Ondiz Sánchez, F. Perea Ganchou, R. Cruz Arámbulo, G. Portillo Martínez y E. Soto Belloso. 2002. Evaluación ultrasonográfica del crecimiento del folículo ovulatorio en vacas anéstricas mestizas Cebú post-tratamiento con Norgestomet y eCG. Archivos Latinoamericanos de Producción Animal 10(1): 20-23. [ Links ]
D’occhio, M.J., A. Neish and L. Broadhurst. 1990. Differences in gonadotrophin secretion postpartum between Zebu and European breed cattle. Animal Reproduction Science. 22(4): 311–317.
Figueiredo, R.A., C.M. Barros, O.L. Pinheiro and J.M.P. Soler. 1997. Ovarian follicular dynamics in Nelore breed (Bos indicus) cattle. Theriogenology 47(8): 1489-1505. [ Links ]
Ginther, O.J., J.P. Kastelic and L. Knopt. 1989. Composition and characteristics of follicular waves during the bovine oestrous cycle. Animal Reproduction Science 20: 187-200. [ Links ]
Giraldo, C.A.; Z.T. Ruiz, L.F. Restrepo y M. Olivera. 2005. Interrupción temporal del amamantamiento (ITA) en vacas Cebú y su efecto en la función ovárica. Revista Electrónica de Veterinaria REDVET 6(12): 1-11. [ Links ]
Fortune, J.E. 1994. Ovarian follicular growth and development in mammals. Biology of Reproduction. 50(2): 225-232. [ Links ]
Henao, G. 1998. Descripción y comparación del restablecimiento del ciclo estral postparto en vacas Brahman sin y con amamantamiento en el trópico colombiano. Tesis Magister en Ciencias Reproducción Animal. Universidad de Antioquia, Facultad de Medicina Veterinaria y Zootecnia, Medellín. 28 p. [ Links ]
Henao, G., L.E. Trujillo y Vásquez, J.F. 2000. Cambios en la dinámica folicular de vacas Cebú anéstricas sometidas suspensión temporal de la lactancia. Revista Colombiana de Ciencias Pecuarias. 13(2): 121-129. [ Links ]
Henao, G, A.M. Olivera and J.G. Maldonado. 2000. Follicular dynamics during postpartum anestrus and the first estrous cycle in sukled or non-sukled Brahman (Bos indicus) cows. Animal Reproduction Science 63: 127-136. [ Links ]
Henao, G.; L.E. Trujillo y J.F. Vásquez, 2002. Actividad ovárica posparto de vacas Cebú en amamantamiento. Revista Facultad Nacional de Agronomía, Medellín 55(1): 1285-1302. [ Links ]
Henao, G. y L.E. Trujillo. 2003. Dinámica folicular durante la gestación temprana: estudio de un caso en Bos indicus. Revista Facultad Nacional de Agronomía, Medellín 56(1): 1779-1788. [ Links ]
Henao, G. y V. González. 2008. Relación de la variación del peso vivo y de la condición corporal con la dinámica folicular posparto en vacas cebú primerizas. Revista Facultad Nacional de Agronomía, Medellín 61(1): 4394-4399. [ Links ]
Lucy, M.C., J.D. Savio, L. Badinga, R.L. De La Sota and W.W. Thatcher. 1992. Factors that affect ovarian follicular dynamics in cattle. Journal of Animal Science 70(11): 3615-3626. [ Links ]
Machado, R; M.A.C.M. Bergamaschia, R.T. Barbosa, C.A. de Oliveira, M. Binelli. 2008. Ovarian function in Nelore (Bos indicus) cows after post-ovulation hormonal treatments. Theriogenology 15(7): 798-804. [ Links ]
Mahecha, L.; D. Henao, M. Cardona y A. Olivera, M. 2003. Interrupción temporal de la lactancia (ITA) y presentación de estro en vacas Cebú posparto. Revista Colombiana de Ciencias Pecuarias 16(3): 215-219. [ Links ]
Monniaux, D.; C. Huet, N. Besnard, F. Clément, F. Bosc, C. Pisselet, P. Monget and J.C. Mariana. 1997. Follicular growth and ovarian dynamics in mammals. Journal of Reproduction and Fertility (Suppl.) S1: 3-23. [ Links ]
Moreira V. J.H., A. De Moraes F., W. Ferreira De Sa e L.S. De Almeida C. 2000. Follicular dynamics in zebu cattle. Pesquisa Agropecuária Brasileira. 35(12): 2501-2509. [ Links ]
Moreira V., J.H., L.S. De Almeida C., A. De Moraes F., W. Ferreira De Sa, C.A. De Carvalho F. and A. De Pino M.J. 2004. Short intervals between ultrasonographicaly guided follicle aspiration improve oocyte quality but do not prevent establishment of dominant follicles in the Gir Breed (Bos indicus) of cattle. Animal Reproduction Science 84(1-2): 1–12.
Murphy, M.G., M.P. Boland, and J.F. Roche, 1990. Pattern of follicular growth and resumption of ovarian activity in postpartum beef suckled cows. Journal of Reproduction and Fertility 90: 523-533. [ Links ]
Pearson, R.A. and J.O. Ginther. 1984. Ultrasonography of the bovine ovary. Theriogenology 21(3): 495-504. [ Links ]
Perea G., F; González F., Rumualdo; Cruz A., Robert; Soto B., Eleazar; Rincón U., Edmundo; Gonzàlez S., Carlos y Villamediana M., P. 1998. Evaluación ultrasonográfica de la dinámica folicular en vacas y novillas mestizas. Revista Científica FCV/LUZ 8(1): 14-24. [ Links ]
Peter, A.T. 2004. An update on cystic ovarian degeneration in cattle. Reproduction in Domestic Animals 39(1): 1-7. [ Links ]
Plasse, D. 1994. Factores que influyen en la eficiencia reproductiva de bovinos de carne en América Latina tropical y estrategias para mejorarla. p. 82-120. En: Memorias Seminario Internacional de Manejo de la Reproducción Bovina. CIPEC, Cartagena. [ Links ]
Rajakoski. E. 1960. The ovarian follicular system in sexually mature heifers with especial reference to seasonal, cyclical, and left-right variations. Acta Endocrinológica, Copenhage 34 (Suppl. 3): S7-S68. [ Links ]
Reis e Silva, AR,. A. de los Reyes, M.L. Gambarini, R. Rumpf, C.C. Oliveira, B. Dias de Oliveira Filho. 2005. Dinâmica folicular por ultra-sonografía em novilhas pré-púberes da raça Gir. Archivos Latinoamericanos de Producción Animal 13(2): 51-55. [ Links ]
Rexroad, C.E. and L.E. Casida. 1975. Ovarian follicular development in cows, sows and ewes in different stages of pregnancy as affected by number of corpora lutea in the same ovary. Journal of Animal Science 41(4): 1090-1097. [ Links ]
Roa, N; T. Linares, T. Días and F. Chacin. 2006. Ondas foliculares ováricas en vacas Brahman y Mestizas (Bos indicus x Bos taurus), ubicadas en los llanos centrales venezolanos. Zootecnia Tropical 24(3): 297-306. [ Links ]
Rhodes, F.M., G. D´Ath and K.W. Entwistle. 1995. Animal and temporal effects on ovarian follicular dynamics in Brahman heifers. Animal Reproduction Science 38(4): 265-277. [ Links ]
Ruiz-Cortés, Z.T. and M. Olivera-Ángel. 1999. Ovarian follicular dynamics in suckled zebu (Bos indicus) cows monitored by real time ultrasonography. Animal Reproduction Science 54(4): 211-220. [ Links ]
Sartorelli E.S, L.M. Carvalho, D.R. Bergfelt, O.J. Ginther and C.M. Barros. 2005. Morphological characterization of follicle deviation in Nelore (Bos indicus) heifers and cows. Theriogenology 63(9): 2382-2394. [ Links ]
Savio, J.D., M.P. Boland, N. Hynes and J.F. Roche. 1990. Resumption of follicular activity in the early postpartum period of dairy cows. Journal of Reproduction and Fertility 88(2): 569–579.
Short, R.E., R.A. Bellows, R.B. Staigmiller, J.G. Berardinelli and E.E. Custer. 1990. Physiological mechanisms controlling anestrus and infertility in postpartum beef cattle. Journal of Animal Science 68(3): 799-816. [ Links ]
Soto, R., I. Rubio, C.S. Galina, E. Castillo and S. Rojas 2001. Effect of pre and post partum feed supplementation on the productive and reproductive performance of grazing primiparous Brahman cows. Tropical Animal Health and Production 33(3): 253-264. [ Links ]
Spicer, L.J., K. Leung, E.M. Convey, J. Gunther, R.E. Short and H.A. Tucker.1986. Anovulation in postpartum suckled beef cows. I. Association among size and number of ovarian follicles, uterine involution, and hormones in serum and follicular fluid. Journal of Animal Science 62(3): 734-741. [ Links ]
Sunderland, P.G. Knights, M.P. Boland, J.F. Roche, and J.J. Ireland. 1996. Alteration in intrafollicular levels of different molecular mass forms of inhibin during development of follicular and luteal phase dominant follicles heifers. Biology of Reproduction, abstract 54(2): 453-462. [ Links ]
Taylor, C. and R. Rajamahendran. 1991. Follicular dynamics corpus luteum grwth and regression in lactating dairy cattle. Canadian Journal of Animal Science 71(1): 61-68. [ Links ]
Toribio, R.E, J.R. Molina, M. Forsberg, H. Kindahl and L.E. Edqvist. 1995. Effects of calf removal at parturition on postpartum ovarian activity in Zebu (Bos indicus) cows in the humid tropics. Acta Veterinaria Scandinava 36(3): 343-352. [ Links ]
Torres-Junior, J.R. de S., M. de F.A. Piresb, W.F. de Sá, A. de M. Ferreira, J.H.M. Viana, L.S.A. Camargo, A.A. Ramos, I.M. Folhadella, J. Polisseni, C. de Freitas, C.A.A. Clemente, M.F. de Sá Filho, F.F. Paula-Lopes, P.S. Baruselli. 2008. Effect of maternal heat-stress on follicular growth and oocite competence in Bos indicus cattle. Theriogenology 69(2): 155-166. [ Links ]
Trevizoli F., H. 2007. Dinâmica folicular, concentração sérica de hormônio luteinizante e citologia vaginal de fêmeas nelore (Bos taurus indicus) submetidas à sincronização da ovulação. 59 p. Dissertação apresentada para obtenção do grau de Mestre em Ciência Animal junto à Escola de Veterinária da Universidade Federal de Goias. GOIÂNIA. [ Links ]
Webb, R.; R.G. Gosden, E.E. Telfer and R.M. Moor. 1999. Factors affecting folliculogenesis in ruminants. Animal Science 68(2): 257-284. [ Links ]
Zeitoun, M.; H.F. Rodríguez, and R.D. Randel. 1996. Ovarian follicular dynamics in suckled zebu (Bos indicus) cows monitored by real time ultrasonography. Therigenology 45(8): 1577-1581 [ Links ]

