










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.2 Medellín July/Dec. 2010
EFECTO DEL PROPILENGLICOL SOBRE LA UREMIA, GLICEMIA Y LA PRODUCCIÓN DE COMPONENTES DE LA LECHE
EFFECT OF PROPYLENEGLYCOL ON URAEMIA, GLYCAEMIA AND MILK COMPONENTS PRODUCTION
Sandra Guisela Durango Morales1; José Ignacio Ruiz Diaz2 y Rubén Darío Galvis Góez3
1 Zootecnista. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <shelitazoo@gmail.com>
2 Zootecnista. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <nachord18@hotmail.com>
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias.. A.A.1779, Medellín, Colombia. <rdgalvis@unal.edu.co>
Recibido: Agosto 31 de 2009; Aceptado: Enero 12 de 2011.
Resumen. Con el objetivo de determinar el efecto de la inclusión de propilenglicol (PG) sobre las concentraciones plasmáticas de urea y glucosa y sobre la cantidad de leche producida y su concentración de sólidos, se analizó un grupo de 10 vacas entre su segunda y quinta lactancia. Los animales recibieron al día 45 posparto PG y durante 3 días consecutivos, éste se suministró diariamente en dosis de 250 g durante cada ordeño con la ración de suplemento comercial. Los animales fueron muestreados antes y después de suministrar el PG. Se estimaron balances de proteína degradable en rumen (PDR), proteína no degradable en rumen (PNDR) y energía neta de lactancia (ENL). El balance promedio de PDR antes y después de suministrar el PG fue de 53,17 (+/-7,32) y 59,79 (+/-12,7) como porcentaje de los requerimientos respectivamente. Se encontró una diferencia estadísticamente significativa para el balance de ENL. Respecto al contenido de sólidos en leche no hubo una diferencia significativa para los porcentajes de grasa y de proteína, sin embargo se evidenció una disminución en la cantidad total de grasa producida por vaca y un aumento en la cantidad total de proteína producida por vaca. Ni la producción de leche ni las concentraciones plasmáticas de glucosa y urea se afectaron con el suministro del PG.
Palabras clave: Calidad composicional, glucosa plasmática, urea plasmática, precursores gluconeogénicos, energía neta de lactancia.
Abstract. In order to determine the effect of including propyleneglycol (PrGl) on plasmatic concentrations of urea and glucose as well as milk produced and solids concentration, a group of 10 cows was studied between second and fifth lactation. The animals received PrGl at 45 day postpartum during 3 consecutive days. The estimated variables were rumen degradable protein (RDP), undegradable rumen protein (URP), and net energy for lactation (NEL). The RDP average before and after administering PrGl, was 53.17 (±7.32) and 59.79 (±12.7) as a percentage of requirements, respectively. There was a statistically significant difference for the balance NEL. Regarding content solids in milk, there was no significant difference in the fat and protein percentage; however, a decrease in total amount of fat produced and a increased in the total amount protein per cow were registered. Milk production and plasmatic concentrations of glucose and urea were not affected with PrGl supply.
Key words: Compositional quality, glucose plasmatic, BUN, gluconeogenic precursors, net energy for lactation.
Los sistemas especializados de producción lechera en Colombia utilizan intensivamente la fertilización nitrogenada. Sin embargo, ésta ocasiona cambios importantes en la calidad nutritiva de los forrajes puesto que éste incrementa el nivel de proteína cruda pero a partir de nitrógeno no proteico, lo cual aumenta la proteína degradable en rumen (PDR) e incrementa la demanda de los carbohidratos no estructurales. Estos dos parámetros de calidad nutritiva de los forrajes alteran significativamente el funcionamiento metabólico del animal.
Galvis, Correa y Ramírez (2008) evaluaron el efecto de los excesos de PDR sobre la disponibilidad de precursores gluconeogénicos, encontrando que el grupo de vacas que presentó el valor más alto de amonio hepático, también registró los balances más bajos de proteína metabolizable, PDR y proteína no degradable en rumen (PNDR), esto puede indicar que la elevación en los valores de amonio hepático no obedecieron al suministro dietario de fuentes proteicas; de tal modo que la elevación del amonio hepático posiblemente se debió al uso de aminoácidos corporales como fuente de energía, tanto para oxidación como para gluconeogénesis. Es así como Overton et al. (1998a, 1998b) sugieren que en vacas peri parturientas, las cuales, se caracterizan por un balance energético negativo (BEN), la tasa de gluconeogénesis a partir de alanina es más del doble comparada con la tasa observada en el preparto. Por su parte Reynolds, Harmon y Cecava (1994), afirman que la utilización de aminoácidos para gluconeogénesis da cuenta del 50% del amonio que se utiliza para la síntesis de urea. Lo anterior concuerda con los resultados reportados por Galvis, Correa y Ramírez (2008), dado que en su investigación reportó una relación significativa entre los valores de amonio hepático y el BEN, una elevación en los valores de amonio hepático que no se correspondieron con los niveles de amonio plasmático ni con el balance de PDR, una relación directa entre los valores de amonio hepático y piruvato hepático, una relación inversa entre los niveles de amonio hepático y el balance de energía. Todo lo anterior sugiere que la elevación de amonio hepático, provino directamente de la desaminación oxidativa de aminoácidos dada la continua remoción de los aminoácidos corporales para mantener la tasa de gluconeogénesis; así gran parte de los aminoácidos, fueron utilizados como fuente de energía, elevando las concentraciones de amonio en el tejido hepático; al mismo tiempo, esto genera una continua síntesis de piruvato. La relación inversa entre las concentraciones de amonio hepático y el balance de energía neta de lactancia indica que los animales que presentaron un balance de energía mayor necesitaron utilizar menor cantidad de aminoácidos como fuente de energía que los animales con un balance energético menor.
Lo anterior supone un aporte importante de carbonos de los aminoácidos hacia la síntesis de glucosa, debido a la continua utilización de glucosa por las vacas lactantes y a su disponibilidad limitada. Todo esto es debido a las características propias de las dietas para vacas lecheras en el trópico alto colombiano, las cuales son deficitarias en carbohidratos no estructurales que representan el sustrato principal que genera en el rumen, la mayor cantidad de propionato, principal precursor gluconeogénico.
En principio, hay dos mecanismos proveedores de glucosa; su absorción desde el intestino delgado y su formación por gluconeogénesis a partir de precursores que se absorben o se movilizan en el cuerpo (Knowlton, et al., 1998). Dada la extensa fermentación ruminal de carbohidratos, la absorción intestinal de glucosa es poco significativa. Por lo tanto, la gluconeogénesis se constituye en el principal mecanismo proveedor de glucosa. En este punto se hace necesario sopesar el costo que implicaría el que el sustrato principal sea el propionato o los aminoácidos gluconeogénicos. De esta manera, el problema de la gluconeogénesis se concentra en la cantidad de propionato y aminoácidos gluconeogénicos disponibles. Según Reynolds et al. (2003) durante el periodo de transición a la lactancia la contribución que hace el propionato a la síntesis de glucosa oscila entre 50 y 60%; el lactato oscila entre el 15 y 20%, mientras que el glicerol solo aporta un 4%. Por diferencia, se estima que los aminoácidos aportan entre 20 y 30% de la glucosa sintetizada vía gluconeogénesis (Overton y Waldron, 2004).
El propilenglicol (propano-1,2-diol) (PG), es un compuesto que realiza un metabolismo similar al propionato (Nielsen e Ingvartsen, 2004), por lo tanto, puede ser utilizado como un precursor gluconeogénico exógeno con el fin de poder evaluar variables de interés zootécnico que se vean afectadas con dicho precursor. En los sistemas de producción lechera especializada en Colombia los animales están expuestos a déficit de precursores gluconeogénicos, lo cual los obliga a utilizar aminoácidos como fuente de carbonos para la gluconeogénesis. Bajo esta situación es lógico pensar que la suplementación con PG, disminuya la utilización de aminoácidos por las rutas metabólicas diferentes a la formación de proteínas de la leche. En la presente investigación se evaluó el efecto de la adición de PG sobre las concentraciones plasmáticas de urea y glucosa, y sobre la producción de proteína láctea, variables consideradas como indicadores del balance de precursores gluconeogénicos.
MATERIALES Y MÉTODOS
Localización. El trabajo se realizó en el Centro Agropecuario Paysandú de la Universidad Nacional de Colombia, sede Medellín, ubicado a 2400 msnm, con una temperatura promedio de 14 °C y una humedad relativa promedio de 80% en una formación ecológica de bmh-MB, según la clasificación de Holdridge.
Animales. Se utilizaron 10 vacas Holstein, las cuales se encontraban entre su segunda y quinta lactancia y en su día 45 posparto.
Dieta. Los animales pastorearon praderas de pasto kikuyo (Pennisetum clandestinum), sometidas al manejo tradicional con fertilización compuesta, y recibieron suplementación alimenticia con concentrado comercial acorde con su peso y nivel de producción.
En este grupo de animales se realizó una toma de muestras de sangre y leche el día previo al experimento (día 44 posparto), las cuales sirvieron de control. El día 45 posparto fue suministrado a los 10 animales 500 g de PG divididos en dos dosis diarias por tres días consecutivos de 250 g mezclados con el suplemento concentrado durante el ordeño de la mañana y la tarde.
Toma de muestras. Para cada uno de los animales se tomaron muestras de sangre el día previo al experimento (día 44 posparto) y el día último del experimento (día 48), en ambos casos entre las 09:00 y las 10:00 horas. Para la obtención de la sangre se hizo punción en la vena yugular derecha con tubos al vacío. Las muestras fueron centrifugadas a 5000 rpm durante 10 min para separar el suero del plasma y se envasaron en alícuotas de 2,0 mL y fueron conservadas a -8 °C hasta su análisis.
El día 44 y 45 posparto se tomó una muestra de leche durante el ordeño de la tarde y de la mañana respectivamente, constituyéndose éstas, como el muestreo control (antes del experimento). Luego de que los animales fueron sometidos a las condiciones experimentales se procedió a tomar una muestra de leche el día 48 durante el ordeño de la mañana y de la tarde. A cada muestra de leche se le realizó un análisis de porcentaje de grasa y proteína utilizando el equipo Ecomilk®.
Análisis en sangre. En las muestras de suero se cuantificaron las concentraciones de urea mediante el kit urea/Bun color (Biosystems) 11537. Mientras que para estimar la concentración de glucosa se utilizó el kit glucosa (glucosa oxidase/peroxidase) (Biosystems). Las lecturas de ambos kits se realizaron en un espectrofotómetro HITACHI®.
Balance nutricional. La información de la valoración nutricional del pasto y del concentrado fue tomada del estudio realizado por Barrientos y Muñoz (2006), por su parte los requerimientos de nutrientes fueron estimados con el modelo del National Research Council - NRC (1989). Se hizo el balance de energía neta de lactancia (ENL), PDR y PNDR, para cada animal y en cada periodo de muestreo, los cuales se expresaron como porcentaje de los requerimientos del animal. Para estimar la ENL aportada por el forraje se utilizó la propuesta de Mertens (1983). La estimación del contenido de ENL del suplemento alimenticio, así como la estimación del consumo de materia seca se realizó utilizando la metodología propuesta por el NRC (1989).
Análisis estadístico. Para el análisis de los resultados se realizó un diseño completamente al azar y una comparación de medias a través de la prueba de Duncan; así mismo, se realizó un análisis de regresión entre algunas variables. Para cada análisis se utilizó el paquete estadístico SAS.
RESULTADOS
La Tabla 1 describe las características de los animales utilizados en el experimento.
Tabla 1. Características de los animales utilizados para evaluar el efecto del propilenglicol sobre variables de interés zootécnico.
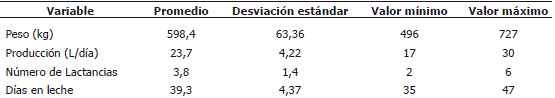
Los animales fueron homogéneos para los días en lactancia (39,3 +/- 4,37) más no así para las demás variables como: peso, producción y número de lactancias. Dado que se conoce que los días en lactancia afectan significativamente las variables evaluadas en este experimento, se consideró necesario incluir su efecto en el análisis estadístico.
La Tabla 2 describe la comparación entre medias para las variables consideradas. Como se puede observar el balance de ENL fue afectado significativamente tanto por la adición del PG como por los días en lactancia.
Tabla 2. Comparación entre medias para establecer el efecto del propilenglicol sobre variables de interés zootécnico.
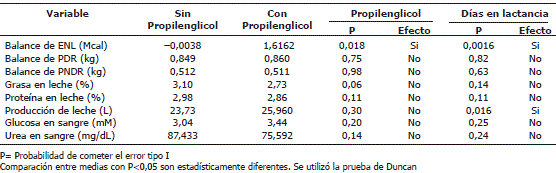
La producción de leche sólo se afectó significativamente por el efecto de los días en lactancia. Por el contrario la adición de PG y los días en lactancia no afectaron las demás variables del experimento.
DISCUSIÓN
Características de los animales utilizados. Como criterio de inclusión de los animales en este experimento se tuvo que las vacas estuvieran alrededor de su pico de producción de leche y que a su vez el tiempo de inicio y finalización de todo el experimento no fuera superior a tres meses, esto para evitar el efecto de los días en lactancia y de la época del año. Dada las anteriores condiciones es de esperar que en un hato de 60 vacas como lo es el del Centro Agropecuario Paysandú fuera necesario incluir animales entre la segunda y la sexta lactancia, esto para evitar los efectos de la primera lactancia y de la avanzada edad sobre las variables a estudiar. Sin embargo, esto no evitó el efecto de la edad sobre la talla y la producción de leche, lo que consecuentemente origino una gran heterogeneidad en peso y producción de leche.
Comparación entre medias de las diferentes variables del experimento. Antes de ser suministrado el propilenglicol, se encontró que éstas tenían un BEN. Se esperaba entonces que al suministrar el precursor gluconeogénico, éste balance energético se volviera menos negativo lo cual se cumplió pues antes de someter a las vacas al tratamiento, el balance promedio de energía neta de lactancia se encontraba en -0,0038 Mcal de ENL y luego del suministro del PG paso a ser de 1,62 Mcal, presentando diferencias estadísticamente significativas (P<0,05). Este aumento en el ENL era de esperarse dado que las vacas recibieron un mayor aporte de energía proveniente del PG, el cual según Miyoshi, Pate y Palmquist (2001) tiene un contenido de energía bruta de 5,66 Mcal/kg, la cual es equivalente a 2,97 Mcal/kg de ENL, valor considerablemente alto debido a que la naturaleza química de este no origina perdidas en el proceso digestivo. Sin embargo, este valor debe ser de mayor magnitud dado que para calcularlo se utilizó la información de pérdidas de energía digestible y metabolizable de alimentos convencionales, los cuales se espera que presenten mayores pérdidas dadas la naturaleza compleja de éstos. Esta situación se origina en la ausencia de información de los valores de energía metabolizable y de energía neta de lactancia del PG.
Los valores de glucosa en sangre estuvieron acorde con los reportados por otros autores para animales bajo condiciones similares de producción (Galvis y Correa, 2003; Galvis, Correa y Ramírez, 2008). Al evaluar los niveles de glucosa en plasma antes y después del tratamiento, no se encontró ninguna diferencia estadísticamente significativa, contrario a lo esperado ya que en teoría al adicionar un precursor gluconeogénico los niveles de glucosa en sangre deberían aumentar. Cozzi et al. (1996), no encontraron ningún efecto significativo del PG sobre incremento de glucosa, insulina y disminución de los ácidos grasos no esterificados (NEFA) al utilizar un rango de dosis entre 0-400 g/día. Al igual que Cozzi et al. (1996), Pickett, Piepenbrink y Overton (2003) no encontraron ninguna diferencia estadísticamente significativa con una dosis de PG de 500 mL/día sobre el incremento de glucosa en sangre; sin embargo, Grummer (1993), encontró una diferencia estadísticamente significativa sobre el incremento de la glucosa en sangre utilizando una dosis de 1 L por día de PG durante diez días pre-parto. Teniendo en cuenta los resultados obtenidos en estos estudios, se puede esperar incrementos en los niveles de glucosa en sangre utilizando dosis más altas de PG, sin embargo, Hindhede (1976), citado por Bergman, (1993) reportó que dosis entre 800 g y 1800 g de PG producen salivación y ataxia en vacas secas. Es probable que el suministro de PG, si aumente la disponibilidad total de glucosa en el animal, pero que los mecanismos homeostáticos para el control de la glicemia sanguínea, no permitan que estos cambios se hagan evidentes, de esta forma el excedente de glucosa es utilizado para procesos diferentes como la síntesis de glicógeno y la producción de leche.
Con respecto a la producción de leche, se presentó un incremento de la misma después de la aplicación del tratamiento; sin embargo, esta respuesta no se debe a un efecto directo del tratamiento (P>0,05) sino al efecto que tuvieron los días en lactancia (P<0,05), lo que era de esperarse dado que en esta etapa de lactancia se observan incrementos constantes en la producción de leche hasta alcanzar el pico de producción. Estos resultados no están acordes con los mencionados por Miyoshi, Pate y Palmquist (2001), quienes encontraron que la inclusión de PG no afectó positivamente la producción de leche, posiblemente debido a su efecto negativo sobre el consumo, dada su baja palatabilidad cuando se suministro a dosis de 518 y 688 g/día, respectivamente. En el presente estudio no se presento disminución en el consumo de concentrado cuando a este se le incorporó el PG, posiblemente debido a dosis relativamente bajas (250 g) mezcladas uniformemente en raciones de entre 2 y 3 kg de suplemento.
Andersen et al. (2002) afirman que el incremento energético en la ración asociado al uso de PG está relacionado directamente con la producción de leche; sin embargo, en esta investigación el aumento en el balance de ENL no estuvo asociado a aumentos en la producción de leche. Esto pudo ser debido a que estas vacas estaban en un balance energético promedio cercano a cero (-0,0145%, de los requerimientos), lo que sugiere que este aporte no tuvo la oportunidad de cambiar sustancialmente el estatus energético de los animales, dada su condición favorable per se.
Tanto la adición de PG como los días en lactancia no afectaron el porcentaje de proteína en leche (P>0,05). Sutton (1989) afirmó que un aumento en la densidad energética de la ración utilizando el PG podría llevar a un incremento en el porcentaje de proteína en la leche; así mismo Griinari et al. (1997); Knowlton et al. (1998), asumen que al incluir el PG en la dieta, el requerimiento de aminoácidos para la gluconeogénesis disminuye con lo que se ganaría un ahorro de éstos aminoácidos que posteriormente se utilizarían para la síntesis de proteína láctea. De ésta manera la ausencia de respuesta al incremento en el porcentaje de proteína en la leche encontrado en el presente estudio al adicionar el PG en la dieta, podría deberse a que la dosis utilizada no fue suficiente para generar reservas de aminoácidos que pudieran ser utilizados en la síntesis de ésta proteína. Una situación que pudo impedir que la adición de PG aumentara el porcentaje de proteína en leche esta relacionada con la etapa de lactancia en la cual se llevo a cabo este experimento: los primeros 60 días de lactancia se caracterizan por un aumento progresivo en la producción de leche, lo que se corroboró al registrar los días en lactancia (DEL) (DEL= 39,3, 23,73 kg/d; DEL=44,3, 25,96 kg/d; P=0,016), de este modo los incrementos en volumen de producción implicaron incrementos en la utilización de amino ácidos para la síntesis de proteína láctea; siendo así, es de esperar que en esta fase de lactancia el ahorro de aminoácidos atribuido a la adición de PG fue utilizado para mantener un porcentaje mínimo de proteína láctea, más que para aumentarlo, debido al efecto de dilución por el aumento gradual en producción de leche; esto se pudo evidenciar con el análisis de producción total de proteína láctea por vaca, pues en solo 5 días de diferencia las vacas aumentaron el promedio de producción de leche en 2,23 kg/d, disminuyeron el contenido promedio de proteína en la leche en 0,12%; sin embargo, el promedio de proteína láctea producido por vaca aumento en 0,0353 kg/d. Este análisis está acorde con lo indicado por Nielsen y Ingvarsten (2004), donde precisan que el PG no afecta el porcentaje de proteína en la leche en vacas en lactancia temprana.
Aunque estadísticamente no se puede concluir que la adición de PG causó una disminución en el contenido de grasa de la leche (P=0,06), es necesario tener en cuenta que este valor está muy cerca del límite de aceptación (P=0,05), y por lo tanto se considera que se evidenció una disminución desde 3,10% a 2,73%, lo que implica una caída de 0,37 unidades porcentuales, equivalente a 0,11%. Contrario a lo sucedido con la cantidad promedio de proteína láctea producida por vaca por día la cantidad promedio de grasa láctea producida por vaca por día, disminuyó de 0,7356 a 0,7087 kg/día. Adicionalmente, los días en lactancia estuvieron lejos de afectar significativamente el porcentaje de grasa en la leche (P=0,14), por consiguiente la disminución tanto en el porcentaje de grasa en leche, como en el total de grasa producida por vaca por día, son atribuibles a la adición de PG. Algunos estudios han reportado que diferentes dosis de PG tienden a disminuir la concentración de grasa en leche; al respecto, Fisher et al. (1973) observaron una disminución significativa en el porcentaje de grasa en leche al suministrar 360 g de PG/día, dicho resultado se lo atribuye a la disminución de los ácidos grasos no esterificados en la plasma y a su consecuente disminución en su disponibilidad para la glándula mamaria. Adicionalmente Nielsen y Ingvarsten (2004), plantean que la disminución de la proporción de acetato/propionato en el rumen originada por la suplementación con PG, limita la síntesis de ácidos grasos en la glándula mamaria, lo que afecta en última instancia la producción de grasa láctea.
El promedio de concentración de urea en plasma antes y después del suministro de PG fue de 87,43 y 75,59 mg/dL respectivamente. Estos valores se encuentran más altos que los obtenidos por otros autores, es así como Galvis et al. (2003), reportaron valores entre 18,4 y 24,0 mg/dL, en tanto que Galvis, Correa y Ramírez (2008), refirieron valores entre 35,03 y 42,75 mg/dL. La diferencia con los valores referenciados en investigaciones anteriores pudo ser debida a balances de PDR significativamente diferentes, es así como en la presente investigación el balance promedio de PDR antes y después de suministro de PG fue de 53,17 (+/-7,32) y 59,79 (+/-12,7) como porcentaje de los requerimientos, respectivamente; mientras que para Galvis y Correa (2003), al día 35 de lactancia el balance de proteína cruda osciló entre 0 y 20% de los requerimientos; y para Galvis, Correa y Ramírez (2008), el balance promedio de PDR oscilo entre 2,75 y 13,14% de los requerimientos. Estos resultados sugieren una tendencia al aumento en las concentraciones de urea en sangre en respuesta al exceso de PDR, que se hace evidente solo cuando se contrastan diferentes investigaciones con variaciones grandes entre si, respecto al balance de PDR. El promedio de concentración de urea en sangre disminuyó después de aplicado el tratamiento con el PG; sin embargo, esta disminución no fue estadísticamente significativa (P>0,05). Esta leve variación en la concentración de urea sanguínea era de esperarse si se observa el gran exceso de PDR (Tabla 2) que presentaron los animales durante el período de estudio, y esto según Annison y Bryden (1999) origina una alta tasa de transformación de amonio ruminal en urea, por lo cual el efecto del PG sobre esta variable no pudo ser significativo.
Es de resaltar que estudios con bajas dosis de PG (210 g/d), realizados en otras condiciones de producción como el de Shingfield et al. (2002), causaron disminuciones significativas de la urea en leche, si bien estos investigadores no hacen referencia los efectos sobre la urea sanguínea, es de esperar que se hayan presentado disminuciones, dado que según Ghonda y Linberg (1994) y Galvis, Correa y Ramírez (2008), existe una alta correlación entre los valores de urea en leche y en sangre.
En el presente experimento utilizando dosis más altas de PG no se pudo evidenciar un efecto significativo. Lo anterior pudo ser debido a las condiciones particulares de producción en el trópico alto colombiano, que aunadas a las condiciones metabólicas adversas de la lactancia temprana pudieron impedir una respuesta significativa en los niveles de urea plasmática. De este modo según Galvis, Correa y Ramírez (2008) sugieren que dada la magnitud del déficit energético de la vaca en lactancia temprana, esta utiliza una gran cantidad de aminoácidos como fuente de energía, lo que genera una fuente de producción de amoniaco adicional a la generada por los excesos de PDR. De este modo esta situación demandaría una mayor cantidad de moléculas como el PG para disminuir significativamente la utilización de aminoácidos como fuente de energía, lo que no permitiría una reducción significativa en la urea plasmática. Resultados similares fueron reportados por Hidalgo et al. (2004), quienes suministraron 250 mL de PG a novillas durante siete días, sin observar efectos sobre las concentraciones de urea en plasma, aunque estos autores no discuten al respecto, es de esperar que en novillas, las cuales están sometidas a un balance nutricional y metabólico favorable, no haya un cambio significativo en los niveles de urea plasmática, ya que estos animales se encuentran en un estado anabólico que genera muy poca cantidad de desechos nitrogenados.
BIBLIOGRAFÍA
Andersen, J.B., T. Larsen, M.O. Nielsen and K.L. Ingvartsen. 2002. Effect of energy density in the diet and milking frequency on hepatic long chain fatty acid oxidation in early lactating dairy cows. Journal Veterinary Medicine 49(4):177-183. [ Links ]
Annison, E.F. and W.L. Bryden. 1999. Perspectives on ruminant nutrition and metabolism. II. Metabolism in ruminant tissues. Nutrition Research Reviews 12(1): 147-177. [ Links ]
Barrientos, S.M. y Y. Muñoz. 2006. Efecto de niveles crecientes de nitrógeno no proteico dietario sobre algunas variables metabólicas y nutricionales en vacas lactantes, Trabajo de grado. Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias, Medellín. 22 p. [ Links ]
Bergman, E. 1993. The Pools of cellular nutrients: Glucosa. p. 173-196. In: P.M. Riis (ed.). Dynamic biochemistry of animal production. Elsevier Science Publishers, The Netherlands. Chapter 9. [ Links ]
Cozzi, G., P. Berzaghi, F. Gottardo, G. Gabai, and I. Andrighetto. 1996. Effect of feeding propylene glycol to mid-lactating dairy cows. Animal Feed Science Technology 64(1): 43-51. [ Links ]
Fisher, L.J., J.D. Erfle, G.A. Lodge and F.D. Sauer. 1973. Effects of propylene glycol or glycerol supplementation of the diet of dairy cows on feed intake, milk yield and composition, and incidence of ketosis. Canadian Journal of Animal Sience. 53(2): 289-296. [ Links ]
Galvis, R. y H. Correa. 2003. Interacciones entre el metabolismo y la reproducción en la vaca lechera: es la actividad gluconeogénica el eslabón perdido? Revista Colombiana de Ciencias Pecuarias 15(1):36-50. [ Links ]
Galvis, R., H. Correa y N. Ramírez. 2008. Proteína degradable en rumen, metabolismo del nitrógeno y precursores gluconeogénicos en vacas lactantes. Documento para optar a la categoría de Profesor Asociado. Departamento de Producción Animal, Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín. [ Links ]
Gonda, H. and J. Linndberg. 1994. Evaluation of dietary nitrogen utilization in dairy cows based on urea concentrations in blood, urine and milk and on urinary concentrations of purine derivatives. Acta Agriculturae Scandinavica, Section A - Animal Science 44(4): 236-245. [ Links ]
Griinari, J.M., M.A. McGuire, D. Dwyer, D.E. Bauman, D.M. Barbano and W.A. House, 1997. The role of insulin in the regulation of milk protein synthesis in dairy cows. Journal of Dairy Science 80(10): 2361-2371. [ Links ]
Grummer, R. R. 1993. Etiology of lipid–related metabolic disorders in periparturient dairy cows. Journal of Dairy Science 76(12): 3882-3896.
Knowlton, K.F., T.E. Dawson, B.P. Glenn, G.B. Huntington and R.A. Erdman. 1998. Glucose metabolism and milk yield of cows infused abomasally or ruminally with starch. Journal of Dairy Science 81(12): 3248-3258. [ Links ]
Hindhede, J. 1976. Propylenglykol til malkekøer omkring kælvning. Meddelelse, Statens. Husdyrbrugsfrosøg 146: 1-4. [ Links ]
Hidalgo, C.O., E. Gómez, L. Prieto, P. Duque, F. Goyache, L. Fernández, I. Fernández, N. Facal and C. Díaz. 2004. Pregnancy rates and metabolic profiles in cattle treated with propyleneglycol prior to embryo transfer. Theriogenology 62 (3-4): 664–676.
Mertens, D.R. 1983. Using neutral detergent fiber to formulate dairy rations and estimate the energy content forages. p. 60. In: Proceedings of the Cornell Nutrition Conference For Feed Manufacturers.Syracuse, NY, Cornell Univ., Ithaca, NY. [ Links ]
Miyoshi, S., J.L. Pate, and D.L. Palmquist. 2001. Effects of propylene glicol drenching on energy balance, plasma glucose, plasma insuline, ovarían function and conception in dairy cows. Animal Reproduction Science 68(1-2): 29-43. [ Links ]
Nielsen, N.I. and K.L. Ingvartsen. 2004. Propylene glycol for dairy cows a review of the metabolism of propylene glycol and its effects on physiological parameters, feed intake, milk production and risk of ketosis. Animal Feed Science and Technology 115: 191-213. [ Links ]
National Research Council (NRC). 1989. Nutrient requirements of dairy cattle. Sixth Revised Edition. National Academy of Sciences, Washington D.C. 168 p. [ Links ]
Overton, T.R, J.K. Drackley, C.J Otteman-Abbamonte, A.D. Beaulieu and J.H. Clark. 1998a. Metabolic adaptation to experimentally increased glucose demand in ruminants. Journal of Animal Science 76(7): 2938-46. [ Links ]
Overton, T.R., J.K. Drakley, G.N. Douglas, L.S. Emert and J.H. Clark. 1998b. Hepatic gluconeogénesis and whole body protein metabolism of periparturuent dairy cows as affected by sources of energy and intake of the prepartum diet. Journal of Dairy Science 81 suppl, 1: 295. [ Links ]
Overton, T.R. and M.R. Waldron. 2004. Nutritional management of transition dairy cows: Strategies to optimize metabolic health. Journal of Dairy Science 87:(E. Suppl. 1):E105-E119. [ Links ]
Pickett, M.M, M.S. Piepenbrink and T.R. Overton. 2003. Effects of propyleneglycol or fat drench on plasma metabolites, liver composition and production of dairy cows during the periparturient period. Journal of Dairy Science 86(6): 2113-2121. [ Links ]
Reynolds, C,K, D.L. Harmon and M.J. Cecava. 1994. Absorption and delivery of nutrients for milk protein synthesis by portal-drained viscera. Journal of Dairy Science 77(9): 2787-2808. [ Links ]
Reynolds, C.K., P.C. Aikman, B. Lupoli, D.J. Humphries and D.E. Beever. 2003. Splanchnic metabolism of dairy cows during the transition from late gestation through early lactation. Journal of Dairy Science 86(4): 1201-1217. [ Links ]
Statistical Analysis System (SAS). 1995. User’s guide: Statistics. Version 1995. SAS Institute Inc., Cary, North Carolina, USA. 1028 p.
Shingfield, K.J., S. Jaakkola and P. Huhtanen. 2002. Effect of forage conservation method, concentrate level and propyleneglycol on diet digestibility, rumen fermentation, blood metabolite concentrations and nutrient utilization of dairy cows. Animal Feed Science and Technology 97(1): 1-21. [ Links ]
Sutton, J.D., 1989. Altering milk composition by feeding. Journal of Dairy Science 72(10): 2801-2814. [ Links ]

