










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.2 Medellín July/Dec. 2010
PATRONES DE DISTRIBUCIÓN ESPACIAL DE ESPECIES ARBÓREAS EN BOSQUES DE ALTA MONTAÑA DEL DEPARTAMENTO DE ANTIOQUIA, COLOMBIA
SPATIAL PATTERNS OF DISTRIBUTION OF TREE SPECIES IN HIGH MOUNTAIN FORESTS AT THE REGION OF ANTIOQUIA, COLOMBIA
Ronald Alfonso Montañez Valencia1; Carmen Yuliet Escudero Vásquez2 y Álvaro Javier Duque Montoya3
1 Ingeniero Forestal. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <ramontan@unal.edu.co>
2 Ingeniera Forestal. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <cyescude@unal.edu.co>
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <ajduque@unal.edu.co>
Recibido: Noviembre 29 de 2010; Aceptado: Enero 26 de 2011
Resumen. Se evaluó el patrón de distribución espacial de especies arbóreas a lo largo de un gradiente altitudinal en bosques de alta montaña. El estudio se realizó en tres (3) parcelas permanentes de 1 ha ubicadas en bosques alto andinos al norte de la cordillera Central de Colombia, clasificando los datos en dos estratos arbóreos: dosel (DAP≥10cm) y sotobosque (DAP<10cm). El objeto es responder si: 1. ¿Son similares los patrones de distribución entre dosel y sotobosque? 2. ¿Existen diferencias en el patrón de distribución a lo largo del gradiente altitudinal? Para definir el patrón de distribución de las especies a diferentes escalas se empleó el índice estandarizado de Morisita (Ip). El patrón de distribución predominante fue el gregario para especies de dosel y sotobosque en las tres áreas de estudio. El grado de agrupamiento de las especies de dosel aumentó a medida que incrementó la escala o el tamaño de la parcela. En contraste, en el sotobosque el gregarismo disminuyó con la escala espacial de análisis. Lo anterior sugiere que la denso-dependencia aumenta proporcionalmente con la altitud. Esta tendencia, no obstante, parece estar controlada por mecanismos reguladores contrastantes, tales como la limitación en dispersión y la especialización de hábitat, entre especies del dosel y el sotobosque respectivamente.
Palabras clave: Denso-dependencia, gregarismo, gradiente altitudinal, índice de Morisita, limitación en dispersión.
Abstract. The assessed the patterns of spatial distribution of tree species along an altitudinal gradient in high mountain forests it was evaluated. The study was carried out in three 1-ha plots located in high Andean forests on the northern part of the Central cordillera of Colombia, classifying the data into two categories of size: canopy (DBH>10 cm) and understory (DBH<10 cm). The main research questions were: 1. Are there differences in the spatial patterns of species distribution between both canopy and understory species? 2. How much and in which way can the spatial patterns of species distribution be affectted by the altitudinal gradient? The standardized Morisita index (Ip) was used to assess the pattern of species distribution at different spatial scales. The degree of clustering of canopy species increased along with the size of the plot. In contrast, understory species showed the oppositte pattern. The results suggest that density-dependence processes proportionally increased with the elevation. This trend, however, seems to be controlled by different mechanisms on either canopy or understory, such us dispersal limitation or habitat specialization, respectively.
Key words: Density-dependence, aggregated species, altitudinal gradient, Morisita index, dispersal limitation.
El estudio de la dispersión espacial de los individuos por especie es un factor fundamental para comprender o determinar el efecto de sucesos pasados sobre el patrón de distribución actual, lo cual permite generar hipótesis sobre los procesos biológicos o ambientales que estructuran los bosques tropicales (Dale, 1999; Levine, 1992). Evaluar el grado de agregación o dispersión de las especies, permite además identificar mecanismos y factores que mantienen y promueven la coexistencia de especies y la diversidad vegetal de los ecosistemas naturales (Hyatt et al., 2003).
Los principales mecanismos que se han planteado para intentar explicar los patrones de distribución de los árboles al interior de una comunidad vegetal, pueden resumirse en tres: 1. Procesos denso-dependientes tales como competencia excluyente o alta mortalidad de juveniles por ataques de plagas o enfermedades (Janzen, 1970; Connell, 1971), los cuales se han asociado con patrones de distribución uniforme o disperso; 2. Estocasticidad demográfica por mortalidad aleatoria, asociada a patrones de distribución aleatoria; 3. Limitación en dispersión (Hubbell 1979, 2001) o especialización de hábitat (Gentry, 1988), relacionada con patrones agrupados o distribución gregaria. Estos mecanismos hacen parte de teorías ampliamente conocidas que han intentado explicar la alta riqueza de especies en los bosques tropicales (Wright, 2002; Hubbell, 1979; Lieberman y Lieberman, 1994).
Todas las anteriores teorías ecológicas suponen que dichos mecanismos, en algunos casos excluyentes entre sí, actúan independientemente de las condiciones ambientales. Sin embargo, los cambios en las condiciones ambientales podrían conducir a una interacción entre varios de los mecanismos expuestos (Vormisto et al., 2004; Duque et al., 2009), o incluso a una supuesta diferenciación paulatina a lo largo del gradiente (López y Duque, 2010). Por ejemplo, se ha propuesto que la diversidad a disminuye con la altitud (Rosenzweig, 1995), mientras que la b aumenta como posible consecuencia del aumento de microhábitats (Kattan, Álvarez y Giraldo, 1994). De esta manera, lo que se esperaría a lo largo de un gradiente altitudinal es que el grado de gregarismo de las especies, aumente proporcionalmente con el incremento en la elevación sobre el nivel del mar.
El presente estudio, llevado a cabo en bosques alto andinos en la región norte de la cordillera Central de Colombia, Departamento de Antioquia, busca identificar los patrones de distribución en términos del grado de gregarismo, uniformidad o aleatoriedad de las especies arbóreas, según el índice de Morisita, en tres parcelas permanentes localizadas sobre un gradiente altitudinal. Las preguntas a responder fueron: 1) Son similares los patrones de distribución entre dosel y sotobosque? y 2) Existen diferencias en el patrón de distribución a lo largo del gradiente altitudinal? Respondiendo estas preguntas, se espera contribuir a la identificación de herramientas y oportunidades de manejo y conservación de los ecosistemas boscosos tropicales, y más específicamente, de los muy degradados ecosistemas andinos de alta montaña.
MÉTODOS
Área de estudio. Para el estudio, se usó información proveniente de tres parcelas permanentes ubicadas en el norte de la Cordillera Central en el departamento de Antioquia, Colombia (Figura 1). La reserva forestal “La Forzosa” se localiza al nororiente de la Cordillera Central, al sur oriente del municipio de Anorí a 1.750 msnm, entre las coordenadas geográficas 6°59´35´´N y 75°08´44´´W. Posee una temperatura mensual promedio entre 18 y 24°C y una precipitación promedio anual de 2.400 mm (Gutiérrez, 2002). “La Forzosa” se encuentra dentro de la zona de vida bosque muy húmedo premontano (bmh-PM), según Holdridge (1996). En el municipio de Angelópolis, se localiza la hacienda La Argentina, sitio de establecimiento de la segunda parcela, a una altitud de 2.133 msnm entre las coordenadas geográficas 6°09´32´´N y 75°41´99´´W. El sitio posee una temperatura mensual promedio entre 12 y 18°C y una precipitación promedio anual de 2.800 mm. Esta región está clasificada en la zona de vida bosque muy húmedo montano bajo (bmh-MB). Por último, la región de Belmira está ubicada al Norte del departamento de Antioquia, a una altitud es de 2.800 msnm (EPM, 1991), entre las coordenadas geográficas 6°36´90´´N y 75°39´46´´W. La temperatura mensual promedio oscila entre los 8 y 14°C, mientras que la precipitación promedio anual es de 2.100 mm. Esta región ha sido clasificada dentro de la zona de vida bosque muy húmedo montano (bmh-M) (Espinal, 1992).
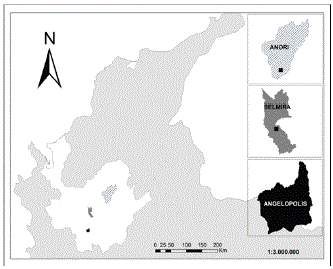
Figura 1. Localización de las tres regiones en la cordillera Central del departamento de Antioquia; Colombia, donde se establecieron parcelas permanentes de vegetación arbórea. Escala aproximada 1: 3’000.000.
Toma de datos
Método de muestreo. El protocolo de muestreo fue exactamente igual para las tres regiones. En cada una, se realizó el montaje de una parcela permanente de una hectárea (1 ha) de forma cuadrada (100 m x 100 m). En cada parcela se censaron todos los individuos arbóreos, incluyendo palmas y helechos arbóreos, con diámetro a la altura del pecho (DAP) ≥ 10 cm; a su vez, en cada parcela de 1 ha se ubicó una sub-parcela de 40 m x 40 m donde se censó la vegetación entre 1 ≤ DAP < 10 cm, teniendo en cuenta la vegetación leñosa además de los hábitos de crecimiento antes señalados.
Análisis de datos. Todos los individuos muestreados fueron determinados por los especialistas locales y nacionales hasta la máxima resolución botánica que fue posible en el Herbario de la Universidad de Antioquia (HUA). Para efectos del análisis, se dividieron los datos en dos categorías estructurales según el DAP, tomado a una altura constante del suelo de 1,30 m. Se clasificaron como especies de dosel a todos aquellos individuos cuyo DAP fue mayor o igual a 10 cm. Se definió como sotobosque, todos aquellos individuos cuyo DAP estuvo entre 1 cm y 9,9 cm. Esta clasificación, ya empleada en otros estudios (Duque et al., 2002; López y Duque, 2010), se hace más con fines prácticos para diferenciar categorías de tamaño que con el objetivo de diferenciar hábitos de crecimiento. Por este motivo, dentro de la categoría de sotobosque, se incluyen tanto juveniles como especies arbustivas de poco crecimiento.
Para los análisis de gregarismo se consideró un umbral de veinte (20) individuos como mínimo por especie arbórea en cada área y cada categoría estructural. Para la definición del patrón de distribución se usó el índice de Morisita, haciendo uso del programa Arc Gis v9.3. (ESRI, 2008) para asignar las coordenadas numéricas que definían la posición de los individuos dentro de cada parcela. Para el cálculo del índice de Morisita (Id), se crearon tamaños de cuadrantes de 10 x 10, 20 x 20 y 50 x 50 m en dosel, tomando como muestra el total de la parcela; en el sotobosque el tamaño de los cuadrantes varió en 5 x 5, 10 x 10, 20 x 20 m. La representación de cada una de las escalas obtendrán diferentes replicas dentro de la evaluación del índice de Morisita; es decir 100, 25 y 4 para dosel y 64, 16 y 4 en sotobosque.
Índice de Morisita (Id). Emplea los valores críticos de uniformidad (Mu) y agrupamiento (Mc), para ser utilizados en la homogeneidad del índice estandarizado de Morisita (Ip), con valores entre -1 y +1, donde Ip = 0 presenta una distribución aleatoria, Ip > 0 distribución agregada y Ip < 0 distribución uniforme (Cabrera y Wallace, 2007). El Ip se define como:
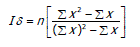
Donde, n es el número de cuadrantes, x es el número de individuos de la especie i y x2 es el número de individuos de la especie i, en el cuadrante i, elevado al cuadrado.
El Ip se realiza mediante los siguientes parámetros (Krebs, 1985). Sí,
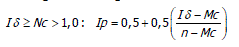
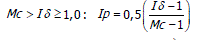
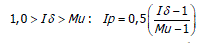
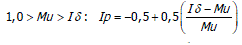
El Ip fluctúa de -1,0 a 1,0 con límites de confianza al 95%, denominando límite inferior a -0,5 y limite superior +0,5 determinando el grado de dispersión ó patrón de distribución de las especies de la siguiente manera: patrones aleatorios dan un valor de Ip igual a cero; patrones agregados por encima de cero; patrones uniformes por debajo de cero.
Valores críticos:

valor de Chi-cuadrado con n-1 grados de libertad, con 97,5% intervalo de confianza.
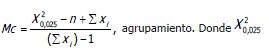
valor de Chi-cuadrado con n-1 grados de libertad, con 2,5% intervalo de confianza.
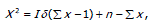
Donde, X2 es la prueba estadística, con distribución Chi-cuadrado.
La comparación de medias ( ) entre las escalas de análisis en estratos arbóreos a lo largo de un gradiente altitudinal, se evaluó empleando la distribución t, la desviación estándar (sd) y el número de especies (n), para construir límites de confianza superior e inferior que permiten evaluar el comportamiento de los patrones de distribución. Utilizando la expresión:
Limite superior e inferior 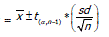
RESULTADOS
El total de especies analizadas que cumplieron con el mínimo de abundancia varió en función de estratos y rango altitudinal: en el sitio más bajo de Anorí se registraron 12 especies en dosel y 14 en sotobosque; en Angelópolis se encontraron 6 especies para dosel y 17 en sotobosque; y en la región de Belmira se reportaron 3 y 14 especies, respectivamente. El patrón de distribución espacial de las especies que predominó entre los estratos de dosel y sotobosque para las áreas de estudio, según el Ip fue el gregario (Tabla 1). Las cinco (5) especies que presentaron mayor grado de agrupamiento en dosel para toda la escala de análisis con el Ip fueron: Tibouchina lepidota (Bonpl.) Baill, Quercus humboldtii Bonpl, Brunellia sibundoya Cuatre, Protium tovarense Pittier y Guettarda crispiflora Vahl. Las cinco (5) especies que presentaron una mayor agregación en sotobosque fueron: Piper sp nov, Miconia multiplinervia Cogn, Psychotria jervisei (Standl.) CM. Taylor, Aphelandra scolnikiae Leonard y Miconia sp1.
Tabla 1. Listado de especies arbóreas, analizadas por el índice de Morisita (Ip) para establecer el patrón de distribución de especies arbóreas en la cordillera Central del departamento de Antioquia, Colombia.
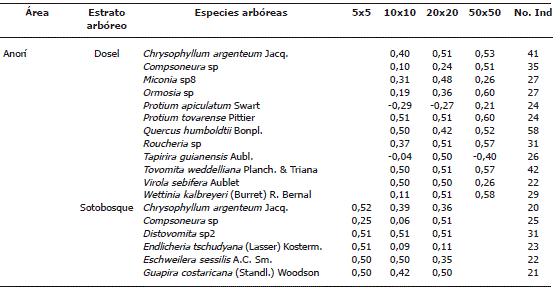
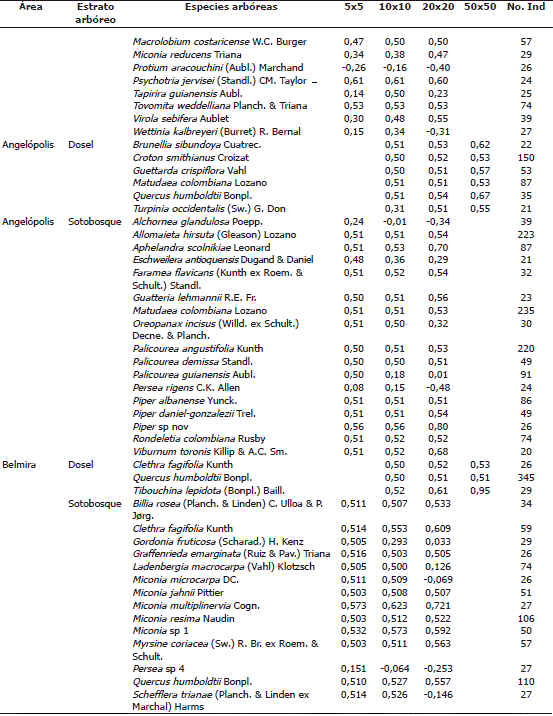
En el estrato arbóreo de dosel, Anorí presentó una tendencia al incrementó del gregarismo proporcional a la escala espacial. En Angelópolis y Belmira todas las especies de dosel presentaron un patrón de distribución gregario independiente de la escala. Cuando se consideró el sotobosque, el grado de agrupamiento de las especies disminuyó a medida que aumentó la escala de análisis. Según el Ip, en Anorí, dos especies de dosel tuvieron distribución uniforme, Tapirira guianensis Aubl y Protium apiculatum Swart. En el sotobosque en cambio el número de especies con patrón de distribución uniforme aumento con el aumento de la escala espacial. La representación de la distribución aleatoria fue nula (Tabla 2).
Tabla 2. Patrones de distribución a diferentes escalas para los estratos arbóreos en cada área de estudio en la cordillera Central del departamento de Antioquia, Colombia.
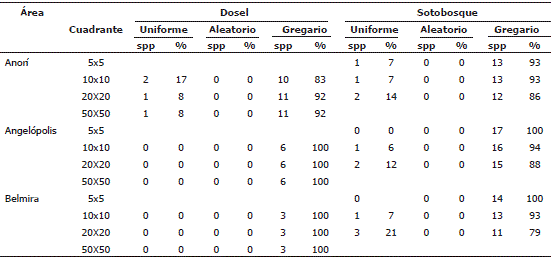
Con respecto al gradiente altitudinal, el patrón de gregarismo de las especies de dosel se mantuvo constante con respecto a la escala de análisis, siendo siempre mayor en promedio a mayor altitud (Figura 2). Al analizar las especies de sotobosque, a escalas espaciales finas, el comportamiento entre sitios fue similar, pero disminuyendo drásticamente en Belmira, el sitio más alto, a escalas espaciales más gruesas (20x20 m) (Figura 3). En resumen, el gregarismo en el dosel aumentó y en el sotobosque disminuyó con la escala de análisis, lo que muestra tamaños de parche de ocupación del dosel más grandes que los representados por las especies a nivel del sotobosque.
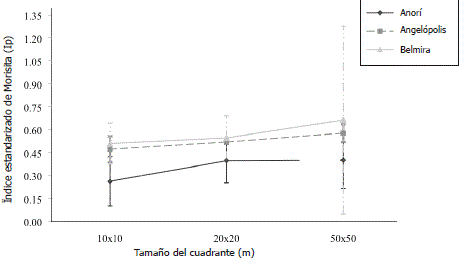
Figura 2. Comparación de patrones de distribución de especies arbóreas a lo largo de un gradiente altitudinal para el estrato dosel. Las líneas horizontales representan el valor promedio y las verticales los limites superior e inferior a escalas de 10x10, 20x20 y 50x50 m en la cordillera Central del departamento de Antioquia, Colombia.
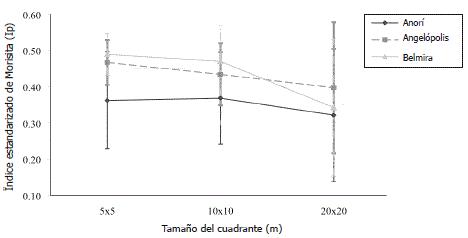
Figura 3. Comparación de patrones de distribución de especies arbóreas a lo largo de un gradiente altitudinal para el estrato sotobosque. Las líneas horizontales representan el valor promedio y las verticales los limites superior e inferior a escalas de 5x5, 10x10, 20x20 m en la cordillera Central del departamento de Antioquia, Colombia.
DISCUSIÓN
Este estudio, confirma lo reportado en otros bosques tropicales de tierras bajas donde se encontró que el patrón de distribución gregario fue el dominante (Hubbell, 1979; Condit et al., 2000). Por otra parte, el patrón de aleatoriedad observado fue nulo, lo cual sugiere la existencia de un patrón de mortalidad controlado por procesos determinísticos que evitan la distribución aleatoria de las especies dentro de los bosques tropicales. Así mismo, el patrón de distribución uniforme, el cual se asume puede emerger como producto del escape de individuos a distancias razonables de sus parentales (Janzen, 1970; Connell, 1971), tuvo muy baja representación. De esta manera, dentro de cada bosque, la limitación en dispersión (Hubbell, 2001) o la especialización de hábitat (Tilman, 1982), aparecen como los mecanismos determinantes de la distribución de las especies.
En las especies del dosel, donde el patrón de agrupamiento aumentó con el incremento del tamaño del área de muestra, se reduce la probabilidad de que la especialización de hábitat sea el mecanismo promotor de este tipo de distribución, ya que esto demuestra el poco efecto de la variación ambiental a escalas pequeñas sobre la distribución de las especies más grandes. Según lo anterior, en ecosistemas de montaña, se puede identificar a la limitación en dispersión (Hubbell, 2001) como el mecanismo de control determinante de la distribución de las especies arbóreas a escala local (Condit et al., 2002; Duque, Cavelier y Posada, 2003). En contraste, la disminución en el patrón de gregarismo de las especies de sotobosque con el aumento del área, parece indicar que la respuesta a la variación de hábitat a escalas mucho más finas de los individuos más pequeños es el mecanismo de control del grado de agrupamiento de las especies en este compartimiento del bosque. En esta medida, los resultados obtenidos apoyan la idea de que la especialización de hábitat o nicho es diferencial con respecto al tamaño de los individuos, siendo mayor durante los primeros estadios de desarrollo (Duque et al., 2002; Ruokolainen y Vormisto, 2002; Comita, Richard, Stephen, 2007).
El grado de gregarismo de las especies leñosas estudiadas mostró una variación positiva y sistemática con el incremento en altitud, tanto en las especies de dosel como en las de sotobosque. Esto significa que a mayor altitud mayor probabilidad de agrupamiento de individuos de una misma especie. Sin embargo, el mayor gregarismo de las especies de sotobosque en Belmira a escalas más pequeñas pero mayores a escalas más grandes en las especies de dosel, en comparación con los otros dos bosques, sugiere una mayor denso-dependencia a mayor altitud. No obstante, este patrón de cambio en la magnitud de los mecanismos reguladores de la dinámica del bosque, además de que requiere ser corroborado con datos temporales, puede estar mostrando un bosque perturbado o sucesional en proceso de cambio. De esta manera, concluir acerca de un cambio gradual y diferencial en los procesos que regulan la distribución de especies arbóreas a lo largo del gradiente altitudinal, resulta bastante sugestivo, pero prematuro aún con los datos aquí presentados. Validaciones futuras de esta tendencia, serán de extrema utilidad para entender mejor el funcionamiento de los bosques de montaña, lo cual permite implementar medidas de conservación más eficaces en estos ecosistemas, actualmente sometidos a altos niveles de amenaza y perturbación (Rodríguez et al., 2004).
Finalmente, es necesario resaltar las implicaciones propias del muestreo de este estudio debido a la alta diversidad de estos bosques del norte de los Andes (López y Duque, 2010), lo cual limita fuertemente el tamaño de muestra por especie; lo anterior, tiene consecuencias directas sobre el Ip, dada su consabida sensibilidad a tamaños de muestra menores a los 50 individuos (Krebs, 1985). Por este motivo, validaciones con base en métricas del gregarismo menos sensibles al tamaño de muestra, como por ejemplo el índice W (Condit et al., 2000) o el K de Ripley (Fortín y Dale, 2005), son necesarias para confirmar las tendencias observadas. Aún así, los resultados aquí presentados son los primeros que tratan de abordar estas preguntas asociadas con el patrón de cambio a lo largo del gradiente altitudinal, abriendo nuevas avenidas de investigación en los Andes tropicales y en los ecosistemas de montaña en general.
CONCLUSIONES
El presente estudio, abre y propone nuevas posibilidades de investigación acerca de los patrones de distribución de las especies arbóreas a lo largo de gradientes altitudinales. Aunque se confirma que los patrones gregarios no son una tendencia exclusiva de especies de tierras bajas, se logra en adición develar un posible cambio gradual asociado con la variación altitudinal, lo cual requiere de estudios adicionales acerca de las características funcionales y la respuesta fisiológica de las especies vegetales a los cambios en temperatura y otros factores ambientales asociados. Llama en especial la atención el patrón opuesto con respecto a la escala espacial de análisis develado entre las categorías de tamaño (dosel y sotobosque), lo cual tiene implicaciones importantes para entender mejor el funcionamiento y manejo de estos ecosistemas tropicales. Dada la escasez de estudios en ecosistemas de alta montaña, esta investigación espera servir de abrebocas y motivador para estudios similares que permitan identificar herramientas y oportunidades de conservación en los bosques tropicales.
AGRADECIMIENTOS
Los autores desean agradecer a CORANTIOQUIA y la Dirección de Investigaciones (DIME) de la Universidad Nacional de Colombia - Sede Medellín, por la financiación para el establecimiento de estas parcelas. A dos evaluadores anónimos quienes con sus comentarios contribuyeron a mejorar el texto. Así mismo, a todos aquellos que de una u otra forma contribuyeron para que el presente trabajo fuera posible.
BIBLIOGRAFÍA
Cabrera, W.H. y R. Wallace. 2007. Densidad y distribución espacial de palmeras arborescentes en un bosque preandino - amazónico de Bolivia. Ecología en Bolivia 42(2): 121-135. [ Links ]
Comita L.S., R. Condit and S.P. Hubbell. 2007. Developmental changes in habitat associations of tropical trees. Journal of Ecology 95(3): 482.492. [ Links ]
Condit, R., P.S. Ashton, P. Baker, S. Bunyavejchewin, S. Gunatilleke, N. Gunatilleke, S.P. Hubbell, R.B. Foster, A. Itoh, J.V. Lafrankie, H.S. Lee, E. Losos, N. Manokaran, R. Sukumar and T. Yamakura. 2000. Spatial patterns in the distribution of tropical tree species. Science 288(5470): 1414-1418. [ Links ]
Condit, R., N. Pitman, E.G. Jr. Leigh, J. Chave, J. Terborgh, B.R. Foster, P. Núñez, S. Aguilar, R. Valencia, G. Villa, H.C. Muller-Landau, E. Losos and S.P. Hubbell. 2002. b-diversity in tropical forest trees. Science 295(25): 666-669. [ Links ]
Connell, J.H., 1971. On the role of the natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. p. 298-310. In: P.J. den Boer y G.R. Gradwell (eds.), Dynamics of populations. Proceeding of the Advanced Study Institute. Centre for Agricultural Publishing and Documentation, Wageningeng. [ Links ]
Dale, MRT. 1999. Spatial pattern analysis in plant ecology. Third edition. Cambridge University Press, Cambridge, UK. 326 p. [ Links ]
Duque, Á., M. Sanchez, J. Cavelier and J.F. Duivenvoorden. 2002. Different floristic patterns of woody understorey canopy plants in Colombian Amazonia. Journal of Tropical Ecology 18(4): 499–525.
Duque, Á., J. Cavelier and A. Posada, 2003. Strategies of tree occupation at a local scale in terra firme forests in the Colombian Amazon. Biotropica 35(1): 20-27. [ Links ]
Duque, Á., J.F. Phillips, P. Von Hildebrand, C.A. Posada, A. Prieto, A. Rudas, M. Suescún and P. Stevenson. 2009. Distance decay of tree species similarity in protected areas on terra firme forests in Colombian Amazonia. Biotropica 41(5): 599-607. [ Links ]
Empresas Públicas de Medellín (EPM). 1991. Boletín Hidrometeorológico, Medellín. 91 p. [ Links ]
ERSI. 2008. Programa de computador ArcGis – ArcView 9, marca registrada de ESRI, Environment Systems Research Institute.
Espinal, L.S. 1992. Geografía ecológica de Antioquia: Zonas de vida. Editorial Lealón,. Facultad de Ciencias, Universidad Nacional de Colombia, Medellín. 146 p. [ Links ]
Fortin, M.J. and M. Dale. 2005. Spatial analysis. A guide for ecologists. Cambridge University Press, Cambridge, UK. 365 p. [ Links ]
Gentry, A.H. 1988. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden 75(1): 1-34. [ Links ]
Gutiérrez-C., P.D.A., 2002. Plan de manejo ambiental para el área de manejo especial la forzosa. Corporación Autónoma Regional del Centro de Antioquia, Corantioquia, Medellín. [ Links ]
Holdridge L. 1996. Ecología basada en zonas de vida. Instituto interamericano de Cooperación para la Agricultura. San José, Costa Rica. 4 reimpresión p. 8-12. [ Links ]
Hubbell, S.P. 1979. Tree dispersion, abundance, and diversity in a tropical dry forest. Science, New Series 203 (4387): 1299-1309. [ Links ]
Hubbell, S.P. 2001. The unified neutral theory of biodiversity and biogeography. Monographs in Population Biology 32. Princeton University Press, Princeton, New Jersey, USA. 448 p. [ Links ]
Hyatt, L.A., M.S. Rosenberg, T.G. Howard, G. Bole, W. Fang, J. Anastasia, K. Brown, R. Grella, K. Hinman, J.P. Kurdziel and J. Gurevitch. 2003. The distance dependence prediction of the Janzen-Connell hypothesis: a meta-analysis. Oikos 103(3): 590-602. [ Links ]
Janzen, D.H. 1970. Herbivores and the number of tree species in tropical forests. The American Naturalist 104: 501-528. [ Links ]
Kattan, G.H., H.L. Álvarez and M. Giraldo, 1994. Forest fragmentation and bird extinctions: San Antonio eighty years later: Source: Conservation Biology 8(1): 138-146. [ Links ]
Krebs, Ch.J. 1985. Ecología: estudio de la distribución y la abundancia. Segunda edición. Editorial Harla, México. 753 p. [ Links ]
Levine, S.A. 1992. The problem of pattern and scale in ecology. Ecology 73(6): 1943-1967. [ Links ]
Lieberman, M. and D. Lieberman. 1994. Patterns of density and dispersion of forest trees. p. 106-119. In: McDade L.A., H.A., Henspenheide and G.S. Hartshorn (eds). La selva: Ecology and Natural history of a neotropical rain forest. University of Chicago Press, Chicago, USA. 493 p. [ Links ]
López, W. y Á. Duque. 2010. Patrones de diversidad a en tres fragmentos de bosques montanos en la región norte de los Andes, Colombia. Revista de Biología Tropical 58(1): 483-498. [ Links ]
Rodríguez, N., D. Armenteras, M. Morales y M. Romero. 2004. Ecosistemas de los Andes Colombianos. Instituto de Investigaciones de Recursos Biológicos Alexander Von Humboldt, Bogotá D.C. 154 p. [ Links ]
Rosenzweig, M.L. 1995. Species diversity in space and time. University of Cambridge, Cambridge, Cambridgeshire, UK. 460 p. [ Links ]
Ruokolainen, K and J. Vormisto. 2002. The most widespread Amazonian palms tend to be tall and habitat generalists. Basic and Applied Ecology 1(2): 97-108. [ Links ]
Tilman, D. 1982. Resource competition and community structure. Monographs in Population Biology (MPB-17), Princeton University Press. 296 p. [ Links ]
Vormisto, J., J.C. Svenning, P. Hall and H. Balslev. 2004. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. Journal of Ecology 92(4): 577–588.
Wright, S.J. 2002. Plant diversity in tropical Forests: a review of mechanisms of species coexistence. Oecología 130: 1:14. [ Links ]

