










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.63 no.2 Medellín July/Dec. 2010
ANILLOS ANUALES Y CLIMA EN RHIZOPHORA MANGLE L. DE LA BAHÍA DE CISPATÁ, COLOMBIA
ANNUAL GROWTH RINGS AND CLIMATE IN RHIZOPHORA MANGLE L. FROM THE CISPATÁ BAY, COLOMBIA
Jorge Andrés Ramírez Correa1; Eliana Cristina Molina Grajales2 y Manuel Bernal Escobar3
1Ingeniero Forestal. Estudiante de doctorado. Université du Québec à Montréal. Centre d’étude de la Forêt. 141 av du President-Kennedy, Québec, Canada, H2X 1Y4. <nicroltda@gmail.com>
2Ingeniera Forestal. Estudiante de doctorado. Université du Québec en Abitibi-Temiscamingue. 445, boulevard de l’université, Rouyn-Noranda, Quebec, Canada, J9X 5E4.<ecmolinag@gmail.com>
3Ingeniero Forestal. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <mbernal@unal.edu.co>
Recibido:Diciembre 17 de 2009; Aceptado: Diciembre 24 de 2010.
Resumen. Por su localización en el ecotono tierra-mar Rhizophora mangle es un organismo con gran sensibilidad al cambio climático. La existencia de anillos anuales climáticamente sensibles en esta especie revela un enorme potencial para la dendroclimatologia tropical. Aunque se ha documentado la existencia de anillos anuales en el género Rhizophora delimitados por cambios en la densidad de poros, aquí se demuestra mediante el uso de la tomografía computarizada que los anillos en R. mangle también están definidos por cambios de densidad de la madera. La comparación entre el número de máximos de densidad de madera (DM) y de poros (DP) en secciones trasversales con la edad de los árboles, reveló la anualidad de los anillos de crecimiento de R. mangle. A mayor precipitación mayor DP y menor DM; y viceversa. La DM y la DP se interpretan como rasgos adaptativos que le permiten a R. mangle adaptarse a los pulsos anuales en salinidad relacionados con las descargas de los ríos en la bahía de Cispatá.
Palabras clave: Dendroclimatología, densitometría, densidad de poros, densidad de madera, tomografía computarizada, manglar, salinidad y crecimiento.
Abstract. Because of its location in the land-sea ecotone Rhizophora mangle is an organism with great sensitivity to climate change. The existence of climate sensitive annual rings in this specie highlights its potential relevance for tropical dendroclimatology. Even though had been previously established the existence of annual rings, delimited by changes in pore-density, in Rhizophora genus, using computed tomographies, is also possible demonstrate, that the rings in R. mangle can also be defined by changes in wood density. Comparing wood density maxima (DM) and pore density maxima (PD) in tree cross-sections reveal the annual nature of R. mangle tree-rings. With increased rainfall PD increases and DM decreases, and vice versa. PD and DM are interpreted as adaptive traits that allow R. mangle to adapt to the conditions produced by annual pulses in salinity in the bay of Cispatá related to high and low discharges of the rivers.
Key words: Dendroclimatology, densitometry, pore density, wood density, computer tomography, mangrove, salinity and growth.
Rhizophora se adapta a sustratos anaeróbicos intercambiando gases a través de las lenticelas localizadas en las partes altas de las raíces (Von Prahl, 1989). Además de estas fluctuaciones de altísima frecuencia, los mangles se han adaptado a fluctuaciones de varios meses de duración: durante los períodos lluviosos los ríos y arroyos aportan abundante agua dulce a los manglares reduciendo la salinidad de sus aguas, en tanto que durante el estiaje el aporte de aguas dulces se reduce significativamente aumentando la salinidad. Rhizophora puede soportar estos cambios de salinidad mediante el mecanismo de la ultrafiltración selectiva llevado a cabo en las raíces, que limitan la entrada de la sal pero no del agua y los nutrientes (Von Prahl, 1989). Ello significa que estos árboles se encuentran sometidos cada año a pulsos o tensiones ambientales que generan estreses hídricos debido a los cambios de salinidad.
Los anillos de crecimiento de los árboles del género Rhizohpora se observan a simple vista como bandas alternas de colores claros y oscuros. Su naturaleza anual fue primero demostrada por Menezes, Berger y Worbes (2003) en Rhizophora mangle L. de Brasil y posteriormente por Verheyden et al. (2004) en Kenia en R. mucronata Lamb. De acuerdo con (Howard, 1934; Carreras, 1988; Verheyden et al., 2004, 2005) los anillos de crecimiento de los árboles de este género, se conforman de bandas alternas de poros escasos y numerosos.
Menezes, Berger y Worbes (2003) demostraron también que el ancho de los anillos de R. mangle almacenaba información ambiental relacionada con la salinidad del sustrato. Luego, Verheyden et al. (2004, 2005) encontraron que las diferencias de porosidad permitían delimitar anillos anuales de crecimiento en R. mucronata de Kenia y que éstas almacenaban información ambiental. También Yu et al. (2004) encontraron anillos anuales en R. apiculata en China, discernibles por las fluctuaciones del dC13 en la celulosa de la madera, útiles para monitorear el incremento del nivel del mar. También se descubrieron anillos anuales en la especie de mangle Laguncularia racemosa (L.) Gaertn. f. (Duque et al., 2008).
La Tomografía Computarizada de rayos X (TC) fue desarrollada por el ingeniero electricista británico Sir Godfrey Newbold Hounsfield en 1979; su invento se empleó en diferentes materiales, incluyendo secciones transversales de troncos (Van Kaick y Delorme, 2005), antes de proponerlo como herramienta de diagnóstico médico (Hounsfield, 1980). Desde entonces el método se ha extendido a numerosas aplicaciones industriales y científicas incluyendo la dendrocronología (De Vore et al., 2006, Okochi et al., 2007). La TC es una imagen por secciones tomada a partir de una máquina de rayos X (tomógrafo) que irradia rayos X y posteriormente los escanea. El dispositivo, envía varios haces simultáneamente desde diferentes ángulos con una potencia determinada, a una sección de espesor constante de un objeto. La reducción de la potencia luego de atravesado el objeto, depende del coeficiente de atenuación del material irradiado que es proporcional a su densidad siguiendo una función exponencial negativa (Ley de Beer-Lamberth). La TC tiene gran potencial en la dendrocronología, pero ha sido poco empleada tal vez por el alto costo de los equipos. Hasta ahora parece que ninguna investigación ha empleado la TC en estudios dendroecológicos en el género Rhizophora ni en ninguna otra especie de árbol tropical.
Este estudio tuvo como objetivos: i) Demostrar la anualidad de los anillos de R. mangle de la bahía de Cispatá (Caribe colombiano); ii) Demostrar que la TC es una herramienta útil en la dendroclimatología con árboles tropicales; iii) Relacionar la densidad de la madera y de los poros de R. mangle con la precipitación en la bahía de Cispatá.
MATERIALES Y MÉTODOS
Localización. El estudio se llevó a cabo en los manglares en torno de la bahía de Cispatá, departamento de Córdoba, Colombia, aproximadamente entre 9°35’-9°41’ N y 75°80’-75°87’ W (Figura 1). Entre 1938 y 1945 el río Sinú cambió su curso y comenzó un nuevo delta en el sitio conocido como Tinajones (Serrano, 2004). Aunque un pequeño brazo con dos ramales principales continuó aportando agua dulce a la bahía de Cispatá, algunos sitios se salinizaron afectándose los manglares llegando en algunos casos a morir. En 1994 se iniciaron algunos trabajos de restauración en la zona afectada consistentes en restablecer los canales naturales de drenaje que estimularan el ingreso del agua dulce y el lavado de la sal mediante las lluvias. También se establecieron pequeñas plantaciones de R. mangle en los sitios más recuperados.
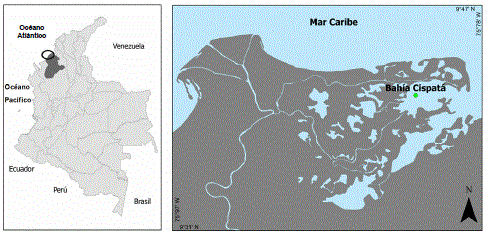
Figura 1. Localización del área de estudio empleada para establecer la relación entre anillos anuales y clima en Rhizophora mangle L.
La precipitación promedia (1995-2005) en la cercana estación climatológica El Limón en San Bernardo del Viento (Colombia), fué 1267± 254 mm anuales (± D.E.), según datos mensuales suministrados por IDEAM (Instituto de Meteorología, Hidrología y Estudios Ambientales). La temperatura promedia es 27,1 ± 1°C (±D.E.). El clima es monzónico; en promedio entre diciembre y marzo cuando soplan los vientos Alisios los meses son secos con precipitaciones (P) menores de 60 mm mensuales (P<60), entre abril y noviembre los meses son húmedos con precipitaciones por encima de los 60 mm mensuales (P≥60) según el sistema de Walter y Lieth (1967) (Figura 2).
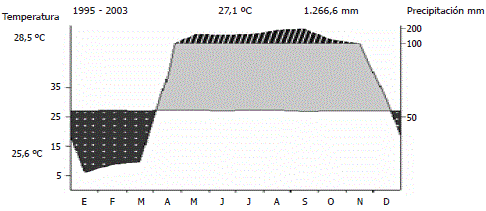
Figura 2. Diagrama climático del área de estudio según Walter y Lieth (1967) elaborado con datos climáticos provenientes de la estación El Limón, San Bernardo del Viento (Colombia). El área negra con puntos blancos indica los meses secos. Las áreas con bandas oblicuas negras y de color gris corresponden a los meses húmedos.
Muestreo. En abril de 2005 se cortaron 8 secciones transversales de aproximadamente 2 cm de espesor del tronco de árboles que crecían en pequeñas plantaciones coetáneas de R. mangle, establecidas con fines de restauración entre 1995 y 1999 en torno de la bahía de Cispatá. Ello implica que se conoce la edad de las plantaciones y permite inferir la edad probable de los árboles dado que estos permanecieron en vivero durante máximo un año. Las secciones se cortaron por encima de las raíces aéreas más gruesas, tan cerca del suelo como fue posible y se secaron al aire hasta que alcanzaron el equilibrio con el ambiente (aproximadamente 13% de humedad en Medellín, Colombia).
Determinación de la densidad intraanular de los anillos. Con el fin de obtener los perfiles de densidad de la madera de los árboles se empleó un escáner “Helicoidal Pro-Speed General Electric Scanner®” perteneciente al Hospital San Vicente de Paúl de Medellín, donde se irradiaron con rayos X las secciones transversales. De cada sección se irradió un disco transversal de 1 mm de espesor durante 2 s a 200 mA y 120 keV. Con el software suministrado por General Electric® para leer las TC y el software Image J 1.34 (Abramoff, Magelhaes y Ram, 2004), se estudiaron los perfiles de densidad de la madera de dos diámetros en cruz (cuatro radios) desde un punto en la periferia, pasando por el centro del disco, hasta el otro extremo del disco. Estos perfiles provienen de trasformar los tonos grises generados por los rayos X en densidad radiológica unidades Hounsfield (HU). De esta manera el software produce un densitograma continuo para cualquier diámetro seleccionado del disco de madera irradiada, en el cual se observan máximos y mínimos de densidad. HU es un número adimensional: HU=[(mi-magua)/magua]•1000, donde mi el coeficiente de atenuación de los rayos X de la madera irradiada y magua el coeficiente de atenuación del agua (Martínez et al., 2002). Para el agua y el aire HU tiene valores de 0 y –1000, respectivamente. HU se relaciona con la densidad física (ρ in g/cm3) mediante la ecuación ρ = 1 + 0,0001 (HU). Se promediaron las cuatro observaciones de densidad radiológica (HU) máxima y mínima de cada anillo. Luego se promediaron las densidades máximas y mínimas del anillo de cada año (suponiendo que los anillos son anuales) de todas las secciones transversales. El ancho de los anillos se midió en los radios del densitograma previamente ampliados hasta 5X midiendo la distancia entre sucesivos puntos de máxima densidad con exactitud aproximada de 0,1 mm. Como en la densidad de la madera se promediaron los resultados de las mediciones.
Posteriormente, con el fin de determinar la densidad de poros de la madera, las secciones transversales se pulieron progresivamente con lija desde grado 80 hasta 600. Cada sección trasversal se frotó con polvo de tiza blanca para resaltar los poros. En cada sección se seleccionaron mediante observación con estéreo-zoom tres radios con la mayor resolución posible de los anillos. Con una cámara digital Mega PCCAM MC 1300 1.0, adosada a uno de los oculares del estéreo-zoom y conectada a un computador cargado con el software MiniSee 2003, se dibujó en cada radio seleccionado, un rectángulo de 2 mm de ancho desde la médula hasta la corteza. Luego se dibujaron dos ventanas rectangulares de 0,4 mm de ancho por 2 mm de altura al inicio de la madera temprana y al final de la madera tardía y se determinó la densidad de poros en ambas ventanas con aumento de 20X. Para cada anillo se promediaron las tres observaciones tomadas al mismo anillo en cada radio. Luego se promediaron las densidades máximas y las mínimas del anillo de cada sección, con las correspondientes al mismo año de las demás secciones.
Funciones respuesta. Se establecieron regresiones lineales en las cuales la variable dependiente fueron diversas expresiones de la densidad de la madera (DM) y de poros (DP) así como del ancho de los anillos en función de diversas expresiones de la precipitación: total, meses secos, meses húmedos, meses con menos de 60 mm incluyendo rezagos de un año. Los resultados se evaluaron con base en el coeficiente de correlación de Pearson. Los coeficientes de Pearson que resultaban significativos al menos para el 95% de confianza se validaron con la prueba de F.
RESULTADOS
Anillos y densidad de poros. En la Figura 3 se presenta una sección transversal de R. mangle. Una observación cuidadosa revela que los anillos, según Kaennel y Schwingruber, (1995) son semiporosos, conformados por una zona con poros escasos y alta densidad de madera (oscura), y otra de poros abundantes y baja densidad de madera (clara), correspondientes a los leños tardío y temprano, respectivamente. No se observan bandas de parénquima o esclerénquima marcando el fin de los anillos, lo que dificulta medir con exactitud y objetivamente el ancho de los anillos.

Figura 3. Fotografía digital de una sección transversal pulida de R. Rhizophora mangle. Se observan zonas de menor y mayor densidad de poros correspondiendo, en su orden, a los leños tardío y temprano. La barra en la parte superior es de 1 cm de longitud y la flecha indica la dirección del crecimiento.
Anillos y densidad de la madera. La Figura 4 presenta la tomografía de la sección transversal 1 (Tabla I). Esta muestra expone seis anillos reales delimitados por bandas anulares de tonos grises claros (madera de alta densidad), precedidos por bandas continuas anulares de colores grises oscuros (madera de baja densidad). También se observan bandas discontinuas interpretadas como anillos falsos. Debajo de la TC se incluye un densitograma que muestra el perfil de la DM a través del un diámetro de la sección transversal, con 6 valores máximos muy notables de densidad máxima, coincidiendo con los anillos de crecimiento tanto en el lado derecho como en el izquierdo. En el lado derecho se insinúan otros valores máximos de DM coincidentes con bandas grises claras discontinuas (anillos falsos). Obsérvese que a pesar de que las diferencias en porosidad entre la madera temprana y tardía son difusas (Figura 3), las diferencias intranulares de DM, por el contrario, son abruptas. De lo anterior se infiere que, como en las regiones templadas, el leño temprano de R. mangle tiene madera de densidad más baja que el tardío. Dado que las muestras se tomaron en abril de 2005, justo al culminar la temporada seca (Figura 2), se debe entender que la madera tardía normalmente se forma durante estos meses; por tanto, la madera temprana se debe formar en promedio entre abril y noviembre. Por esta razón el año fenológico se extiende desde abril hasta marzo, desplazando cuatro meses respecto al año solar. Aunque las regresiones se calcularon con las unidades de densidad radiológica (HU), al lado derecho del densitograma (Figura 4) se indica la densidad física (en g/cm3).
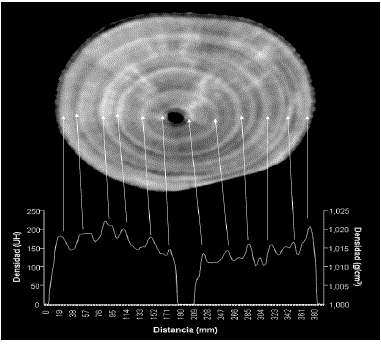
Figura 4. En la parte superior se muestra la imagen escaneada (muestra 4, Tabla I) de una TC correspondiente a una sección transversal de Rhizophora mangle L. de 1 mm de espesor y unos 380 mm de diámetro. Las bandas claras anulares marcan el límite de los anillos reales, las discontinuas son anillos falsos. En la parte inferior se muestra un densitograma cuyos puntos de máxima densidad se corresponden con las bandas grises claras y los mínimos con las bandas grises oscuras de la imagen de la TC. Obsérvese el cambio abrupto de densidad de la madera versus los cambios graduales a difusos de la densidad de los poros (Figura 3); a la izquierda la densidad radiológica (unidades Hounsfield), a la derecha densidad física (g/cm3).
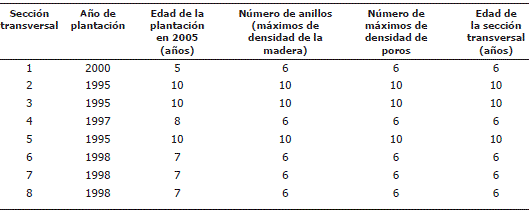
Tabla 1. Comparación entre la edad de las plantaciones (año de plantación menos año de extracción de la muestra) y el número de anillos contados en las secciones transversales de Rhizophora mangle L.
Cabe resaltar la tendencia opuesta de la DM y la DP, el coeficiente de Pearson es negativo entre los máximos (r = -0,81, P = 0,0044) y mínimos (r = -0,62, P = 0,0539) para cada anillo entre las dos variables.
Naturaleza anual de los anillos. De acuerdo con la Tabla 1 se encontró en todas las muestras igual número de puntos de máxima DP y DM (igual si fuesen puntos de mínimas DP y DM). Sin embargo, solo en las secciones 2, 3 y 5 el número de puntos de máxima de DP y DM corresponden exactamente con la edad de las plantaciones y con la edad de las secciones transversales. En las demás secciones la edad de las plantaciones supera en uno o dos años los valores máximos de DP y DM. En relación con estas aparentes inconsistencias es preciso distinguir entre la edad del árbol, la edad de la de plantación y la edad de la sección transversal. La edad de la plantación no tiene en cuenta el tiempo que el árbol permaneció en el vivero. Por ello es posible que un árbol tenga hasta máximo un año más de edad que el que revelaría la fecha de la plantación tomando en cuenta el período que permaneció en el vivero.
Por otra parte, dado que el año fenológico va de abril a mayo y las secciones se cortaron en abril después del período seco, durante 2005 solo se había formado el leño tardío correspondiente al año fenológico iniciado en abril de 2004, tal como se observa en la Figura 4. Por ello es totalmente razonable que la edad de la plantación tenga un año más que el número de picos de densidad observados en la sección transversal. Adicionalmente, la edad de los árboles solo coincidiría muy cercanamente con la edad de las secciones transversales si estas secciones se hubiesen cortado de la base de los árboles. Muestras tomadas a mayores alturas del tronco son cada vez más jóvenes y tienen menos anillos. La abundancia de raíces aéreas en los árboles no permitió muestrear en la base todos los árboles y algunas secciones transversales debieron cortarse a alturas de 0,7 a 1,0 m sobre el suelo.
Parece entonces razonable que los años faltantes entre el número de puntos de máxima densidad tanto de vasos como de madera se explican satisfactoriamente por el tiempo en vivero y por los años requeridos por estos árboles para alcanzar la altura a la cual se cortó cada muestra. Por lo anterior en todos los árboles aquí muestreados el número de máximos de DM y DP, arroja la edad de la sección transversal y no necesariamente la del árbol o la de la plantación. Por tanto, la DM y DP aportan evidencia razonable de la anualidad de los anillos en R. mangle.
Información climática contenida en las características de los anillos. Las correlaciones de la DM tanto máxima como mínima con las diversas expresiones de la precipitación tienen signos opuestos. En cuanto a la DP sucede lo contrario (Tabla 2). No obstante, aunque no todas las correlaciones fueron significativas, la tendencia se mantiene en las variables estudiadas. La DP máxima aumenta significativamente cuando en el año anterior aumenta la precipitación total. El ancho de anillos no correlacionó significativamente con ninguna de las expresiones de precipitación analizadas. La validación con la prueba de F muestra que, efectivamente, los coeficientes de correlación con P<0,05 son significativos. En la Figura 5 se presentan las correlaciones significativas encontradas.
Tabla 2. Coeficientes de correlación de Pearson de las regresiones lineales (y=a+bx) de la función respuesta entre la densidad de la madera (DM) y la densidad de poros (DP) en Rhizophora mangle L. en función de diversas variables de la precipitacióna en la bahía de Cispatá, Colombia. Las correlaciones en negrita son significativas para P<0,05. La probabilidad aparece debajo de los coeficiente de correlación. Entre paréntesis los valores de F calculados para la validación.
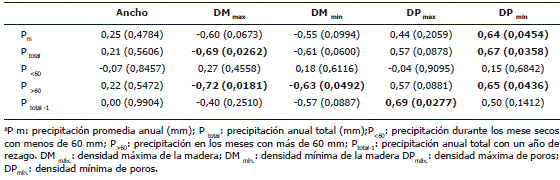
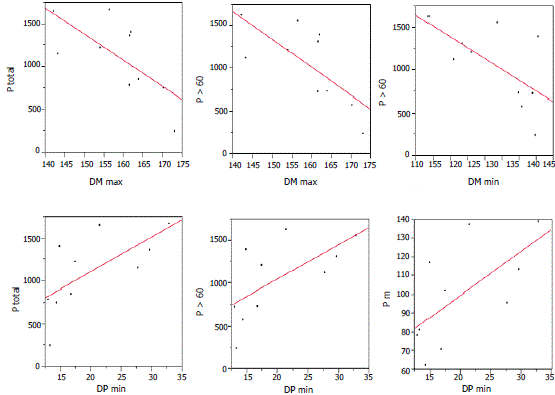
Figura 5. Regresiones significativas encontradas entre densidad de la madera (DM) y densidad poros (DP) en Rhizophora mangle L. y las diferentes expresiones de la precipitación analizadas en la bahía de Cispatá, Colombia.
DISCUSIÓN
La densitometría de rayos X se ha empleado en las investigaciones dendroclimáticas desde que el forestal francés Hubert Polge la descubrió en 1963 debido a que las variaciones intraanulares de la densidad de la madera almacenan más información ambiental que el ancho de los anillos de crecimiento (Cook y Kairiukstis, 1992). La densitometría es de uso cotidiano en algunos laboratorios de dendrocronología del primer mundo. No obstante, el costo de los equipos ha limitado su empleo. En comparación con la TC, la densitometría convencional implica el uso de barreno de incremento y sierras gemelas para obtener muestras de espesor constante (usualmente de 1 mm), las cuales se irradian con rayos X y, posteriormente, la placa se lee en un microdensitómetro. La TC lleva a cabo todos estos pasos simultáneamente de manera más eficiente, rápida y precisa pues no requiere sierras gemelas puesto que el corte es virtual pudiéndose cambiar el espesor con el fin de maximizar la resolución de la imagen, no requiere revelado ni placas físicas porque la información es electrónica, y no requiere microdensitómetro.
Como se aprecia en la Figura 3 la separación de los anillos con base en la porosidad es bastante subjetiva debido al cambio gradual o difuso del tamaño y el número de poros. Por el contrario, el cambio en la densidad de la madera es abrupto (Figura 4) haciendo de la TC un medio más objetivo y eficaz para detectar los anillos de crecimiento. Más aun, el descubrimiento de los anillos falsos es más difícil con los cambios de porosidad que con los cambios de densidad de la madera producidos con la TC.
Los anillos de R. mangle poseen atributos que comparten los anillos con poros difusos y los semiporosos. Se opta por estos últimos debido a la reducción gradual de su tamaño al tiempo que su número aumenta desde el leño temprano hasta el tardío (contrario a DP).
Se considera que una de las posibles causas para la inexistencia de correlación entre el ancho de los anillos y las variables climáticas es la baja exactitud con que estos fueron medidos en las tomografías, 0,1 mm. Generalmente los instrumentos apropiados para la dendrocronología miden con una precisión de 0,001 mm; sin embargo, dichos instrumentos resultaron inútiles en esta especie por la dificultad de fijar exactamente el fin de los anillos por la inexistencia de bandas de parénquima, como es usual en otras especies con anillos con poros difusos o semiporosos.
La muestra utilizada en este estudio, de 8 árboles, permitió demostrar el potencial dendroclimático y el empleo de la TC en R. mangle, seguramente una muestra más grande contribuiría a establecer relaciones más robustas entre las características de los anillos y las variables climáticas. Igualmente, se abre la posibilidad de llevar a cabo reconstrucciones climáticas a partir del desarrollo metodológico utilizado en este estudio con muestras más longevas.
Los resultados aquí reportados ratifican la anualidad de los anillos de Rhizophora previamente reportados por otros autores (Menezes, Berger y Worbes, 2003; Verhayden, 2004, 2005; Yu et al., 2004). En este estudio también se confirma que los cambios de porosidad almacenan información climática relacionada con la disponibilidad de agua dulce y se demuestra la existencia de diferencias de DM entre el leño temprano y tardío en Rhizophora, por medio de TC.
En los resultados se encontró que cuando diversas variables relacionadas con la precipitación aumentan la DM máxima y mínima tiende a reducirse. Lo contrario en cuanto a la DP (Tabla 2, Figura 5). La discusión siguiente trata de probar la hipótesis de que el factor que induce tales cambios anatómicos es la sequía fisiológica periódica a que se ven sometidos los árboles de R. mangle debido a los aumentos episódicos de salinidad del agua del mar. Hallé, Oldeman y Tomlinson (1978) han propuesto a R. mangle como paradigma de crecimiento continuo entre los árboles dicotiledóneos debido a la ausencia de pronunciada segmentación en los brotes; por tanto, descartan que las bandas claras y oscuras observadas a simple vista en la madera sean anillos y, menos aún, anuales (Hallé, Oldeman y Tomlinson, 1978). Aunque estos autores escribieron hace más de 30 años, aun son muy citados para avalar la inexistencia de anillos anuales en especies tropicales. Hasta cuando se disponga de evidencias directas de mediciones de las tasas de crecimiento secundario, como serían las lecturas periódicas -al menos mensuales- de bandas dendrométricas permanentes durante por lo menos un año completo, tal vez no se sabrá a ciencia cierta si en Rhizophora existe o no latencia periódica del cambium que marque los anillos de crecimiento. Ante esta ausencia de información se podrían reconciliar las observaciones de Hallé, Oldeman y Tomlinson (1978) con las evidencias de anillos de crecimiento anuales en Rhizophora que demuestran crecimiento secundario anual rítmico, aceptando que el crecimiento de Rhizophora es continuo pero solo en el sentido de que siempre presenta tasas positivas de crecimiento, aunque estas tasas no son constantes sino variables a través del año: períodos de altas tasas de crecimiento y madera menos densa y más porosa que se alternan con períodos de menores tasas crecimiento y madera más densa y con menos vasos vasculares. De esta manera una reducción significativa en el crecimiento, aunque las tasas nunca lleguen a cero debido a la supuesta no latencia del cambium, podría ser suficiente para marcar los anillos y dejar huellas en la anatomía de la madera como las aquí encontradas.
En R. mangle ocurren cambios fenológicos durante todo el año indicando la inexistencia de quiescencia en las yemas. No obstante, existe considerable evidencia de que tanto en las regiones subtropicales como tropicales Rhizophora presenta períodos de máxima actividad anual (Gill y Tomlinson, 1971; Lugo y Snadeker, 1974; Cintrón y Schaefer, 1983; Lasso y Cantera, 1995; López y Ezequiel, 2002; Mehlig, 2006). Por tanto, parecería lógico esperar que el cambium vascular tuviese un comportamiento similar con períodos de alta y baja actividad debido a las características pulsantes de los ecosistemas de manglar que generan variaciones en la densidad y tamaño de los poros y de la madera (Figuras 3 y 4). El carácter pulsante de los manglares está relacionado con las variaciones intraanuales de tensores que como la salinidad del agua de mar varía en función de los mayores o menores aportes de agua dulce a las bahías: crecientes y estiajes. La salinidad influye en el comportamiento ecofisiológico de Rhizophora produciendo variaciones estructurales en la anatomía de la madera relacionadas con la conducción del agua en la planta: DP y DM.
Rhizophora posee un sistema de ultrafiltrado que impide el ingreso de sal a la planta hasta un umbral que algunos han establecido en 50 partes/1000 (Spalding, 2004; Kathiresan y Bingham, 2001; Lasso y Cantera, 1995; Lüttge, 1997; Raghavendra, 1991; Von Prahl, Cantera y Contreras, 1990; Ball, 1988). Cuando la salinidad supera dicho umbral la planta debe invertir la mayoría de la energía del ATP producida por la respiración en evitar el ingreso de la sal. Ello trae como consecuencia una significativa y periódica reducción de la fotosíntesis, la transpiración, el área foliar y el crecimiento de la planta (Kozlowski, Kramer y Pallardy, 1991; Ball, 1988; Lasso y Cantera, 1995; Lüttge, 1997) que, se estima, causan las modificaciones intra anulares de DM y DP en Rhizophora. La salinidad de los manglares produce potenciales osmóticos bajos y, por ello, sequía fisiológica. Por esta razón la abundancia de agua en los manglares es irrelevante en términos de su disponibilidad para el crecimiento de estas especies. A mayor salinidad durante la estación seca, mayor será la sequía fisiológica a la que están sometidos los árboles y menor el potencial osmótico que los árboles deben superar para obtener agua dulce (Tyree y Sperry, 1989; Raghavendra, 1991). Cuando el potencial osmótico es muy bajo el sistema hidráulico consistente en columnas continuas de agua que se extienden desde las raíces hasta las hojas es vulnerable. Ello implica que pueden pasar burbujas de aire desde vasos vecinos a través de sus punteaduras (cavitación) e interrumpir el flujo dentro de algunos de ellos (embolia) (Maherali, Pockman y Jackson, 2004). Rhizophora supera cíclicamente el límite de vulnerabilidad al embolismo (aproximadamente – 6,0 MPa) reduciendo su conductividad hidráulica hasta en 50% (Tyree y Sperry, 1989; Raghavendra, 1991; Melcher et al., 2001). Por ello se puede afirmar que la alta salinidad actúa en Rhizophora como un factor de estrés químico responsable de la formación de los anillos de crecimiento; la precipitación pluvial es solo una variable sustitutiva (proxy) de la salinidad.
Consistente con las diferentes adaptaciones a la sequía fisiológica, la alta DP del leño temprano le permiten a Rhizophora conducir gran cantidad de agua cuando la salinidad es menor (Martínez y Piñol, 1999; Kathiresan y Bingham, 2001; Meinzer, 2003) y la menor DP del leño tardío reduce el riesgo de cavitación cuando la salinidad aumenta al conducir menor cantidad de agua (Hallgren, 2004; Meinzer, 2003; Martínez y Piñol, 1999; Zimmermann, 1978). Contrario a lo hallado en R. Mangle, Verheyden et al., (2004) encontraron en R. mucronata número de poros en el leño tardío formado durante la estación seca. No obstante, ello parece contrario a la adaptación a la sequía fisiológica. Se sabe, por ejemplo, que Annona glabra, especie acompañante de los manglares en México, desarrolla mayor porosidad cuando crece en medios más salinos para transportar mayor cantidad de agua dulce mientras esta disponible (Yañes y Terrazas, 2001). Adaptaciones similares se han encontrado también en especies de regiones áridas (Kathiresan y Bingham, 2001; Tyree y Sperry, 1989).
Como sucede en muchas especies de las regiones templadas (Schweingruber, 1988), en R. mangle el leño tardío es más denso que el temprano. Ello se debe a que las fibras del leño tardío son más pequeñas y con paredes celulares más delgadas y lúmenes mayores que las del leño temprano. Adicionalmente, en este leño hay menor cantidad de poros y por ende espacios vacíos. Desde una perspectiva ecofisiológica el leño más denso de la madera tardía, otorga mayor seguridad al riesgo de cavitación y embolismo durante la temporada seca y de mayor salinidad. Stratton, Goldstein y Meinzer (2000) encontraron en 6 especies de bosques secos de Hawaii que la conductividad hidráulica de la madera aumentaba 5 veces como consecuencia de pequeñas reducciones en la DM. Meinzer, (2003) también estableció en 27 especies procedentes desde bosques húmedos tropicales hasta desiertos que la DM explicó 96% de la variación en el potencial hidráulico de las hojas; según este trabajo, se gana resistencia a la cavitación al aumentar la DM, al tiempo que se reduce la conductividad hidráulica. La conductividad hidráulica también la restringen en Rhizophora las punteaduras escaleriformes (Carlquist, 1988; Kathiresan y Bingham, 2001; Meinzer, 2003) y substancias viscosas polímeras existentes en la savia que limitan las tasas de flujo y la transpiración. Estas características combinadas con la alta eficiencia en la utilización del agua, típicas de Rhizophora, reducen sus tasas de utilización cuando los contenidos de sal son altos (Zimmermann, 1978; Kathiresan y Bingham, 2001). De acuerdo con Melcher et al. (2001) R. mangle ajusta las propiedades hidráulicas del sistema de transporte del agua, así como el potencial osmótico de las hojas, de acuerdo con las condiciones medio ambientales para el crecimiento.
CONCLUSIONES
Por primera vez en el trópico se emplea la TC en estudios dendroclimáticos mostrándola como una herramienta eficaz, económica y de fácil acceso para relacionar variables ambientales con la densidad de la madera.
R. mangle de la bahía de Cispatá, Colombia, presenta anillos anuales los cuales se caracterizan por variaciones en la DM y DP. El leño temprano de mayor cantidad de poros y madera menos densa se forma durante el período lluvioso (abril-noviembre). El leño tardío, con atributos opuestos al leño temprano, se forma entre diciembre y marzo; presenta madera más densa y menor densidad de poros.
A medida que aumenta la precipitación pluvial la DM máxima y mínima es menor. Lo opuesto con DP máximas y mínimas.
Los ritmos de formación del leño temprano y tardío en R. mangle parecen estar relacionados con estrés hídrico o sequía fisiológica producida por las altas concentraciones de sal durante los períodos de sequía (leño tardío) cuando, aparentemente, los aportes de agua dulce del río Sinú son menores, seguidos por períodos de abundantes aportes de agua dulce (leño temprano). La precipitación es en realidad una variable sustitutiva de la salinidad del agua en la bahía de Cispatá.
R. mangle es una especie tropical con gran potencial dendroclimático dada su ubicación en el ecotono mar-tierra y a que sus anillos de crecimiento almacenan información ambiental relevante en la densidad tanto de la madera como de los vasos.
AGRADECIMIENTOS
Los autores agradecen a la Dirección de Investigaciones Medellín (DIME) de la Universidad Nacional de Colombia, Sede Medellín la financiación parcial de esta investigación, al Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM) por el suministro de la información climática y al Laboratorio de Bosques y Cambio Climático de la Facultad de Ciencias Agropecuarias de la misma Sede por las facilidades para llevar a cabo la investigación. Igualmente están muy agradecidos con el Hospital San Vicente de Paúl por su colaboración en la toma de las tomografías computarizadas.
BIBLIOGRAFÍA
Abramoff, M.D., P.J. Magelhaes and S.J Ram. 2004. Image processing with ImageJ. Biophotonics International 11(7):36-42. [ Links ]
Ball, M.C. 1988. Ecophysiology of mangrove. Trees 2(3):129-142. [ Links ]
Carlquist, S. 1988. Comparative wood anatomy: systematic, ecological, and evolutionary aspects of dycotyledon wood. Springer, Berlin. 432 p. [ Links ]
Carreras, R. 1988. Caracteres anatómicos de la madera de especies típicas de manglares. Consideraciones ecológicas. Revista Forestal Barracoa 18(1): 7-18. [ Links ]
Cintrón, G. y Y. Schaeffer-Novelli. 1983. Introduccion a la ecología del manglar. Oficina Regional de Ciencia y Tecnología de la UNESCO para América Latina y el Caribe, ROSTLAC, Montevideo. 109 p. [ Links ]
Cook, E.R. and L.A. Kairiukstis. 1992. Methods of dendrochronology. Kluwer, Dordrecht. 394 p. [ Links ]
De Vore, M.L., P. Kenrick, K.B. Pigg, and R.A. Detcahama. 2006. Utility of high resolution X-ray computed tomography (HRXCT) for paleobotanical studies: an example using London Clay fruit and seeds. American Journal of Botany 93(12):1848-1851. [ Links ]
Duque, G.C., C. Henriques, M.L. Gomes and C.S. Lisi. 2008. Annual growth rings in the mangrove Laguncularia racemosa (Combretaceae). Trees 22(5):663-670. [ Links ]
Gill, A.M. and P.B. Tomlinson. 1971. Studies on the growth of red mangrove (Rhizophora mangle L.) II. Growth and differentiation of aerial roots. Biotropica 3(1): 63-77. [ Links ]
Hallé, F., Oldeman R.A.A. and P.B. Tomlinson. 1978. Tropical trees and forest: an architectural analysis. Springer, New York. 441 p. [ Links ]
Hallgren, S.W. 2004. Tree physiology: shoot growth and canopy development. p. 1600-1606. Elsevier., Encyclopedia of Forest Sciences. [ Links ]
Hounsfield, G.N. 1980. Computed medical imaging. Science 210: 22-28. [ Links ]
Howard, A. 1934. A Manual of the timbers of the world, their characteristics and uses. MacMillan, London. 672 p. [ Links ]
Kaennel, M. and F.H. Schwingruber (Compilers). 1995. Multilingual glossary of dendrochronology. Swiss Federal Institute for Forest Snow and Landscape Research, Haup, Berne. 467 p. [ Links ]
Kathiresan, K. and B.L. Bingham. 2001. Biology of mangroves and mangrove ecosystems. Advances in Marine Biology 40: 81-251. [ Links ]
Kozlowski, Th., Kramer, T. and S.G. Pallardy. 1991. The phisiologycal ecology of woody plants. Academic, San Diego. 657 p. [ Links ]
Lasso, J. y J.R. Cantera. 1995. Estructura y composición de manglares como indicadores biológicos de condiciones ambientales: comparación entre un bosque riberino y uno de barra en la bahía de Buenaventura, Pacífico colombiano. p. 300-324. En: Delta del río San Juan, bahías de Málaga y Buenaventura, Pacífico colombiano, Tomo II. Colciencias- Universidad Eafit - Universidad del Valle, Cali, Colombia. [ Links ]
López-Portillo, J. y E. Ezequiel. 2002. Los manglares de México: una revisión. Madera y Bosques. Número Especial: 27-51. [ Links ]
Lugo, A. and S.C. Snadeker. 1974. The ecology of mangroves. Ecology Evolution and Systematic 5:39-64. [ Links ]
Lüttge, U. 1997. Physiological ecology of tropical plants. Springer, Berlin. 384 p. [ Links ]
Martínez, L.C., R.C. Rodríguez, C. Muñoz y C. López. 2002. Un método para convertir el número de Hounsfield en densidad electrónica y para la obtención de la energía efectiva en los escáneres CT. Revista de Física Médica 2(1):19-25. [ Links ]
Martínez-Vilalta, J. and Piñol, J. 1999. Arquitectura hidràulica, vulnerabilitat a l’embolisme i resistència a la sequera en plantes llenyoses. Orsis 14:129-157.
Maherali, H., W.T., R.B. Pockman and Jackson. 2004. Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology 85(8): 2184–2199.
Mehlig, U. 2006. Phenology of the red mangrove, Rhizophora mangle L., in the Caeté Estuary, Pará, equatorial Brazil. Aquatic Botany 84:158–164.
Meinzer, F.C. 2003. Functional convergence in plant responses to the environment. Oecologia 134:1–11.
Melcher, P.J., G. Goldstein, F.C. Meinzer, D.E. Yount, T.J. Jones, N.M. Holbrook and C.X. Huang. 2001. Water relations of coastal and estuarine Rhizophora mangle: xylem pressure potential and dynamics of embolism formation and repair. Oecologia 126: 182–192.
Menezes, M., U. Berger and M. Worbes. 2003. Annual growth rings and long-term growth patterns of mangrove trees from the Bragança peninsula, North Brazil. Wetlands Ecology and Management 11: 233-242. [ Links ]
Okochi, T., Y. Hoshino, H. Fujii and T. Mitsutasi. 2007. Nondestructive tree-ring measurements for Japanese oak and Japanese beech using micro-focus X-ray computed tomography. Dendrochronologia 24:155-164. [ Links ]
Raghavendra, A.S. 1991. Physiology of trees. Wiley, New York. 509 p. [ Links ]
Schweingruber, F. 1988. Tree Rings. Basics and applications of dendrochronology. Reidel Publishing Company, Dordrecht. [ Links ]
Serrano, B.E. 2004. The Sinú river delta on the northwestern Caribbean coast of Colombia: bay infilling associated with delta development. Journal of South American Earth Sciences 16: 623-631. [ Links ]
Spalding, M.D. 2004b. Mangroves. p. 1704-1712. In: Burley J., Evans J. & Youngquist J. (eds.). Encyclopedia of Forest Sciences. Academic Press, California. [ Links ]
Stratton, L., G. Goldstein and F.C. Meinzer. 2000. Stem water storage capacity and efficiency of water transport: their functional significance in a Hawaiian dry forest. Plant and Cell Environment 23: 99-106. [ Links ]
Tyree, M.T. and J.S Sperry. 1989.Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Physiology and Molecular Biology 40: 19-38. [ Links ]
Van Kaick, G. and S. Delorme. 2005. Computed tomography in various fields outside medicine. European Radiology (Supplement 4): D74-D81. [ Links ]
Verheyden, A., J. Gitundu, K.H. Beeckman and N. Koedam. 2004. Growth rings, growth ring formation and age determination in the mangrove Rhizophora mucronata. Annals of Botany 94: 59-66. [ Links ]
Verheyden, A., A.F. De Ridder, N. Schmitz, H. Beekman and N. Koedam. 2005. High-resolution time series of vessels density in Kenyan mangrove tree reveal kink with climate. New Phytologist 167: 425-435. [ Links ]
Von Prahl, H. 1989. Manglares de Colombia. Banco de Occidente. Villegas Editores, Bogotá, D.C. 203 p. [ Links ]
Von Prahl, H., J.R. Cantera y R. Contreras. 1990. Manglares y hombres del Pacífico. Fondo FEN-Colombia. Editorial Presencia, Bogotá. 193 p. [ Links ]
Walter, H. and Lieth, H. 1967. Klimadiagramm-Weltatlas. VEB Gustaf Fischer Verlag, Jena. [ Links ]
Yañes-Espinosa, L. and T. Terrazas. 2001. Wood and bark anatomy variation of Annona glabra L. under flooding. Agrociencia 35: 51-63. [ Links ]
Yu, K.F., J.X. Zhao, T.S. Liu, P.X. Wang, J.L. Qiun and T.G. Cheng. 2004. Alpha cellulose d13C variation in mangrove tree rings correlates well with annual sea level trend between 1982 and 1999. Geophysical Research Letters 31(11):L11203-1 – L11203-4.
Zimmermann, M.H. 1978. Structural requirements for optimal water conduction in tree systems. p. 517-532. In: Tropical trees as living systems. Cambridge University Press, London, U.K. [ Links ]

