










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.64 no.1 Medellín Jan./June 2011
PUCCINIA SCHULTESIANTHI, NUEVA ESPECIE BITELIOMÓRFICA SOBRE SOLANACEAE
PUCCINIA SCHULTESIANTHI, NEW BITELIOMORPHIC SPECIES ON SOLANACEAE
Diana Marcela Gallo Agudelo1; Víctor Manuel Pardo Cardona2; Mauricio Salazar Yepes3 y Pablo Buriticá Cespedes4
1 Ingeniera Agrónoma. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779. Medellín, Colombia. <dmgallo@unal.edu.co>
2 Profesor Titular Pensionado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias. A.A. 3840. Medellín, Colombia. <vmpardoc@unal.edu.co>
3 Profesor Asociado. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias. A.A. 3840. Medellín, Colombia. <masalazay@unal.edu.co>
4 Profesor Titular. Universidad Nacional de Colombia, Sede Medellín. Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <pburitica@unal.edu.co>
Recibido: Mayo 27 de 2010; Aceptado: Mayo 19 de 2011
Resumen. Se describe, proveniente de colecciones colombianas, la nueva especie biteliomórfica de Uredinales Puccinia schultesianthi colectada sobre Schultesianthus coriaceus, planta eminentemente neotropical-andina perteneciente a la familia Solanaceae. Se hacen algunas consideraciones taxonómicas acerca de su biología.
Palabras clave: Colombia, royas, Uredinales.
Abstract. Puccinia schultesianthi, a new biteliomorphic species of Uredinales on Schultesianthus coriaceus, Solanaceae, from Colombia is described. Some taxonomical and biological considerations are done.
Key words: Colombia, rust fungi, Uredinales.
Uno de los componentes más importantes de los ecosistemas es el de los Uredinales (hongos, royas), los cuales son parásitos obligados o raramente hemibiótrofos, sobre plantas hospederas que van desde helechos, gimnospermas hasta angiospermas altamente evolucionadas. Los Uredinales se encuentran distribuidos en todas las zonas de vida del mundo donde hay plantas y parecen haber tenido su origen en los trópicos (Hennen y Buriticá, 1980). Algunos Uredinales parasitan plantas cultivadas de gran importancia económica regional y mundial y otros a plantas denominadas “malezas”. En la actualidad se reconocen unas 8.000 especies incluidas dentro de unos 100 a 125 géneros “buenos” (Cummins y Hiratsuka, 2003), inventario que aumenta permanentemente como resultado de los trabajos de colección y reconocimiento.
La biota de los Uredinales colombianos ha sido estudiada desde el siglo XIX y los compendios más completos de la misma han sido publicados por Mayor (1913); Kern y Whetzel (1930); Kern et al. (1933) y Buriticá y Pardo (1996), esta última registró un total de 316 especies para el país. Desde el año de esta publicación a la fecha, Buriticá, Pardo y Salazar han incrementado las colecciones y el estudio del grupo, fruto de lo cual se encuentra en preparación un nuevo listado que incluirá las adiciones de los últimos 14 años.
En septiembre de 2004, ingresó al Museo Micológico de la Universidad Nacional de Colombia, Sede Medellín, una colección de uredinal, con un hospedante no reportado a nivel mundial y el cual en estudios preliminares permitió sospechar que podría tratarse de una especie nueva. Los resultados de este estudio se presentan y discuten a continuación.
MATERIALES Y MÉTODOS
Se realizaron cortes y raspados del uredinal proveniente de la colección sobre Schultesianthus coriaceus (Kuntze) Hunz. (Solanaceae) colectada en la región andina antioqueña. Las muestras fueron montadas en placas con lactofenol e hidrato de cloral, y las observaciones y mediciones fueron hechas en el microscopio de luz Carl Zeiss® “Axiostar plus”. Debido a la presencia de dos soros diferentes: uno de ellos con morfología de Aecidium Persoon (aecidioide) y el otro con morfología del género Puccinia persoon (Puccinioide), desarrollados a partir del mismo micelio y muy similar a los encontrados en Puccinia pampeana Spegazzini, se consideró importante determinar sus tipos de germinación.
Para las pruebas de germinación se realizaron colecciones frescas y se tomaron esporas de los dos tipos (aecidioide y puccinioide), se colocaron en un tratamiento de gota suspendida en agua estéril; para observar y determinar el tipo de germinación. Las observaciones bajo el microscopio se hicieron cada hora el primer día y luego cada 24 horas. En la tinción de núcleos se siguió la metodología de Hiratsuka et al. (1966), usando Giemsa al 5% acidificado.
Todas las observaciones y pruebas se efectuaron en el Museo Micológico de la Universidad Nacional de Colombia sede Medellín (MMUNM), en cuyo herbario se encuentran depositados el tipo de la especie y otras colecciones.
RESULTADOS Y DISCUSIÓN
En ninguna de las colecciones se observaron espermogonios ni morfologías de algún otro tipo de soro. En ocasiones se observó que ambos tipos de esporas se forman en el mismo soro, en un comienzo los aecidioides y posteriormente los puccinioides.
Pruebas de germinación. En las diferentes pruebas efectuadas no fue posible observar la germinación de las esporas puccinioides. Con las esporas aecidiodes se encontró que en las primeras pruebas con material herborizado no se observó germinación, pero en colecciones en las cuales los soros eran jóvenes (recién colectados) la germinación se presentó rápidamente, observándose que hora y media después de montada la prueba, comenzaban a germinar las primeras esporas (alrededor de 10 al 20%) y 24 h después con porcentaje entre 70 y 80%. La ausencia de germinación en las esporas puccinioides puede deberse a que se encuentran en reposo o dormancia. En P. pampeana se encontró que las esporas puccinioides poseen una sustancia autoinhibidora de la germinación que se solubiliza en el agua y que debe ser removida a través de un intenso y prolongado proceso de lavado con agua (Figueiredo y Carvalho Jr., 1994, 1995). En este último caso los autores piensan que las esporas puccinioides germinan en época distinta, mientras que las aecidiodes germinan durante todo el año. No se sabe si las condiciones descritas para P. pampeana se repitan en el caso de P. schultesianthi, pero la función de las esporas aecidiodes parece ser similar en ambas especies.
La germinación de las esporas aecidiodes mostró la formación de un tubo germinativo de crecimiento determinado, con septas formando cuatro divisiones (Figura 1C). A partir de estas divisiones se formaron otras ramificaciones. La tinción de la condición nuclear permitió determinar que en el tubo germinativo se movían 4 núcleos y que cada uno de ellos migraba a una de las cuatro celdas. Aspecto que permite inferir que el proceso ocurrido es el de la germinación de una teliospora, y nomenclatorialmente estas esporas serán, entonces, teliosporas endofiloides. Aunque en la germinación de las esporas endofiloides no hubo formación de basidiosporas fue constatada la ocurrencia de la meiosis a partir de la emisión del promicelio, con septación del mismo y la formación mediante ramificación de células monocarióticas que eventualmente darían origen a micelios monocarióticos compatibles. Lo anterior fue observado mediante tinción de las esporas germinantes con Giemsa al 5% acidificado (Figura 1C).
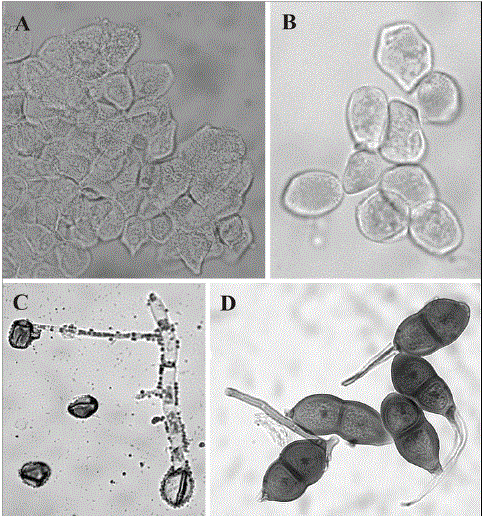
Figura 1. Puccinia schultesianthi (Holotipo). (A) Células peridiales. (B) Teliosporas aecidioides. (C) Germinación teliosporas aecidioides. (D) Teliosporas puccinioides.
Taxonomía. Puccinia schultesianthi Pardo, Gallo, Salazar y Buriticá, sp. nov. Figura 1 A-D.
Etimología. Para ambos estados se sigue el epíteto específico del hospedante.
Spermogoniis, aeciis et urediniis adhuc ignota. Teliis in Endophyllum amphigenis, in maculis circularis vel semicircularis, 1,5-2,0 cm diam, brunneis vel atrobrunneis, outside auriculis; rare dispositis in calycis adque sepalis; cellulae peridii rhomboideis vel subrhomboideis, 22,5-37,5 x 15-22,5 mm; pariete interiore verrocoso, exteriore Levi, hyalinis, umbo 2,5-5 mm crassa; sporis subglobosis vel ellipsoideis, 17,5-25 x 15-20 mm; pariete verrucosa, hyalinis, 0,5-1 mm crassa; poris germinationis obscuris. Teliis in Puccinia amphigenis, agregatis in maculis flavido-brunneis, circularis vel ellipsoideis, 2-12 mm diam, atrobrunneis, epidermide diu tectis, tardem nudis, compacti; sporis plerumque ellipsoideis rare fusiformis, 37,5-62,5 x 20-27,5 mm, apice apiculatis vel truncatis, medio constrictis; pariete flavido-brunneis, levibus, latere 2,5-5,0 mm crassa et apice 2,5-10 mm crassa; pedicello hyalino, persistenti, usque 125 mm longo. Mesosporis rara.
Espermogonios y anamorfos no observados. Teliosoros de Endophyllum anfígenos, en manchas foliares de circulares a semicirculares, de 1,5-2,0 cm de diámetro, de marrón a marrón oscuras, rodeadas por un halo decolorado, raramente en el cáliz o los sépalos; células del peridio presentes, romboides a subromboides, 22,5-37,5 x 15-22,5 mm, pared verrugosas hacia el lado interno y lisas hacia el exterior, hialina, 2,5-5 mm de grosor en el ápice; esporas subglobosas a elipsoides, 17,5-25 x 15-20 mm; pared verrucosa, hialina, 0,5-1 mm de grosor; poros germinativos no observados. Teliosoros de Puccinia anfígenos, agrupados en manchas marrón claro, de circulares a elipsoides, de 2-12 mm de diámetro, marrón oscuros, al comienzo cubiertos por la epidermis, después expuestos, a veces en lóculos, compactos; esporas predominantemente elipsoides, raramente fusiformes, 37,5-62,5 x 20-27,5 mm, con el ápice agudo o truncado, constrictas en el septo, pared marrón-amarillenta, lisa, 2,5-5,0 mm de grosor en los costados y de 2,5-10 mm en el ápice; pedicelo hialino, persistente, hasta 125 mm de longitud. Mesosporas escasas.
Holotipo. Sobre Schultesianthus coriaceus (O. Kuntze) Hunz., Solanaceae, COLOMBIA, Antioquia, Envigado, alto de las Palmas, vereda La esperanza, finca Las Araucarias, 2.500 m. de altura, M. Morales, septiembre de 2004, MMUNM 1610.
Se trata del primer uredinal colectado sobre el género andino Schultesianthus (Solanaceae) y no existe ninguna otra especie biteliomórfica con la cual pudiera compararse, excepto con P. pampeana. La morfología de ambos tipos de esporas es diferente en ambas especies y además en P. schultesianthi no hay formación de espermogonios. Los caracteres diagnósticos se resumen en la Tabla 1.
Tabla 1. Principales características morfológicas de las especies biteliomórficas registradas sobre Solanaceae.

La presencia de espermogonios no fue observada en los diferentes especímenes examinados así como tampoco la de ningún otro tipo de anamorfo. Las pruebas de germinación sugieren que la teliospora endofiloide, debido a su pronta germinación podría ser responsable por la diseminación. En P. pampeana se presenta también una rápida germinación de las esporas endofiloides (E. pampeanum) con formación de basidiosporas (Figueiredo et al., 1987). De otro lado, el telio puccinioide de P. pampeana parece funcionar como un estado de latencia y es necesario darle un tratamiento prolongado con agua para inducir su germinación. En P. schultesianthi este tipo de teliosporas parece actuar como estructura de resistencia y el mecanismo de su latencia debe ser estudiado. La presencia de dos teliomorfos, uno de rápida germinación y otro de latencia (Figura 1), así como la ausencia de espermogonios y de anamorfos y la ocurrencia ocasional de ambas esporas en el mismo soro, sugiere que P. schultesianthi podría tener un ciclo que incluye un micelio dicariótico formado a partir de somatogamia y sería un ciclo reducido biteliomórfico, en el cual el endofiloide es responsable por la mayor diseminación. Solamente la germinación de las esporas puccinioides podría aclarar si en estas esporas ocurre la formación de basidiosporas monocarióticas haploides o si hay otra de las variantes nucleares halladas en los Uredinales. Por ahora lo único claro es que el micelio dicariótico origina los dos tipos de esporas por separado o en sucesión en el mismo soro.
La nominación de esta especie presenta dificultades por poseer dos morfologías teliomórficas (Figura 1), sin que exista un criterio definido para saber con base en cual morfología se asigna el nombre. El criterio que se adopta en este artículo, es considerar que el estado teliomórfico ancestral es el puccinioide y que el endofiloide sería derivado.
La cobertura geográfica de lugares no colectados previamente y la continuada colección y estudio de los Uredinales presentes en el trópico, muestra que todavía es mucho lo que falta por conocer en términos de géneros y especies; estructuras y su función; fenómenos relacionados con los ciclos de vida; y, finalmente, con comportamientos biológicos, ecológicos y fitopatológicos. Las generalizaciones, para todo el orden, hechas con base en los distintos fenómenos que son mejor conocidos en los países de la región templada (Europa y Norte América), muestran estar lejos de proporcionar una realidad global.
Es evidente que la reducción del ciclo de vida de los Uredinales es una respuesta biológica asociada a ciertas familias de plantas hospedantes y a ciertos nichos ecológicos particulares. Algunas evidencias han sido encontradas en la familia Solanaceae en el neotrópico y en la familia Asteraceae de las grandes alturas de los Andes, en donde son muy comunes y endémicas las especies de la familia Pucciniosiraceae.
Durante el período de vida de los Uredinales de ciclo de vida parcialmente y completamente expandido parece existir una separación en el tiempo, entre el estado vegetativo asexual y el reproductivo sexual. En el talo de reproducción asexual (producción de anamorfos), finalmente ocurre la “maduración” sexual del individuo; el micelio “maduro” se encuentra predispuesto para producir únicamente teliosporas. En los Uredinales de ciclo de vida reducido, el micelio se encuentra exclusivamente produciendo el estado sexual que se refleja en la aparición de diferentes tipos de teliosoros y teliosporas, bien siendo unos iguales a sus parentales o en otro tipo de estructura de la espora de origen endofiloide.
Ante tantas variantes de ciclos de vida y función de las estructuras, que ahora comienzan a ser reconocidas y mejor estudiadas, es preciso definir y consolidar criterios taxonómicos que se ajusten a la mayoría de los grupos y especies.
AGRADECIMIENTOS
Los autores expresan sus agradecimientos al Herbario MEDEL por la determinación del hospedante.
BIBLIOGRAFÍA
Buriticá, P. y V.M. Pardo. 1996. Flora uredineana colombiana. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 20(77): 183-236. [ Links ]
Cummins, G.B. and Y. Hiratsuka. 2003. Illustrated genera of rust fungi. Third edition. American Phytopathological Society. Minnesota. 240 p. [ Links ]
Hennen, J.F. and P. Buriticá. 1980. A brief summary of modern rust taxonomic and evolutionary theory. Reports Tottori Mycological Institute 18: 243-256. [ Links ]
Figueiredo, M.B. e A.A. Carvalho. 1995. Presença de auto-inibidor nos teliosporos telióides de Puccinia pampeana e o seu papel na sobrevivencia da espécie. Summa Phytopathologica 21: 229-234. [ Links ]
Figueiredo, M.B. e A.A. Carvalho. 1994. Efeito da lavagem dos soros não germinação dos teliosporos telióides de Puccinia pampeana. Summa Phytopathologica 20: 101-104. [ Links ]
Figueiredo, M.B., C.P. Pimentel, O.M. Russomanno. e L.N. Coutinho. 1987. Biología da espécie biteliomórfica Puccinia pampeana Speg.-Endophyllum pampeanum (Speg.) Lindq., ferrugem da pimenta e do pimentão (Capsicum spp). Arquivos do Instituto Biológico, Sao Paulo 54(1/4): 1-10. [ Links ]
Hiratsuka, Y., W. Morf, and J.M. Powell. 1966. Cytology of the aeciospores and aeciospore germ tubes of Peridermium harknesii and P. stalactiforme of the Cronartium coleosporioides complex. Canadian Journal of Botany 44(12): 1639-1643. [ Links ]
Kern, F.D. and H.H. Whetzel. 1930. Mycological explorations of Colombia. Journal of Department of Agriculture Puerto Rico 14: 301-348. [ Links ]
Kern, F.D., H.W. Thurston and H.H. Whetzel. 1933. Annotated index of the rusts of Colombia. Mycologia 25(6): 448-503. [ Links ]
Mayor, E. 1913. Contribution à l’étude de uredinées de Colombie. Mémoires de la Société des Sciences Naturelles 5: 442-599.

