










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.64 no.1 Medellín Jan./June 2011
EVALUACIÓN DE RESISTENCIA DE GENOTIPOS DE PLÁTANO Y BANANO A LA SIGATOKA NEGRA (MYCOSPHAERELLA FIJIENSIS MORELET.)
EVALUATION OF RESISTANCE OF PLANTAIN AND BANANA GENOTYPES TO BLACK SIGATOKA (MYSCOSPHAERELLA FIJIENSIS MORELET.)
Alejandra Cuéllar Quintero1; Elizabeth Álvarez Cabrera2 y Jairo Castaño Zapata3
1 Ingeniera Agrónoma. Programa Patología de Yuca y Frutas Tropicales. Centro Internacional de Agricultura Tropical (CIAT). Vía Cali - Palmira km 17. Valle del Cauca, Colombia. <alejata05@hotmail.com>
2 Fitopatóloga. Jefe del Programa de Patología de Yuca y Frutas Tropicales. Centro Internacional de Agricultura Tropical (CIAT). Vía Cali - Palmira km 17. Valle del Cauca, Colombia. <e.alvarez@cgiar.org>
3 Fitopatólogo. Profesor Titular. Universidad de Caldas. Calle 65 No. 26-10. Manizales, Colombia. <jairo.castano_z@udecaldas.edu.co>
Recibido: Mayo 27 de 2010; aceptado: Noviembe 28 de 2010.
Resumen. Con el objetivo de evaluar bajo condiciones de invernadero genotipos de plátano y banano frente a poblaciones de Mycosphaerella fijiensis Morelet de diferente virulencia y origen, se estableció una colección de 125 aislamientos monospóricos obtenidos en 10 departamentos de Colombia. Se seleccionaron 50 aislamientos por área geográfica y genotipo hospedante, para la caracterización morfológica y patogénica del hongo en Dominico Hartón. En las pruebas de patogenicidad y resistencia se realizaron inoculaciones de una suspensión acuosa de 5.000 conidias mL-1, la respuesta de las plántulas a la enfermedad se determinó mediante las variables periodo de incubación (PI), tiempo de evolución de los síntomas (TES), área bajo la curva del desarrollo de la enfermedad (ABCDE) y tasa de desarrollo de la enfermedad (r). La inoculación de los 50 aislamientos en plántulas de Dominico Hartón, permitió establecer cinco niveles de virulencia (muy alta, alta, media, baja y muy baja). La virulencia de los aislamientos no se relacionó con su origen geográfico ni con el genotipo del cual se obtuvieron; encontrándose en una misma zona y en un mismo genotipo, aislamientos de diferente virulencia. Los genotipos de plátano y banano mostraron un comportamiento diferencial frente a cinco aislamientos de diferente virulencia y origen, resultados que permitieron establecer tres niveles de reacción de los genotipos frente a los aislamientos (resistente, intermedio y susceptible), destacándose los genotipos Topocho, Maqueño, FHIA 20, FHIA 21 de plátano y los genotipos Sedita y FHIA 23 de banano por presentar un mayor nivel de resistencia, expresada como un progreso lento y menos severo de la enfermedad.
Palabras clave: Musáceas, patogenicidad, virulencia, resistencia.
Abstract. A study was conducted to evaluate plantain and banana genotypes with Mycosphaerella fijiensis Morelet isolates of different virulence and geographic origin. The pathogenicity test and morphological characterization of the M. fijiensis populations were performed with a group of fifty monosporic isolates representative of the geographic growing areas in Colombia. For the pathogenicity and resistance assays an aqueous spore suspension of 5.000 conidias mL-1 was used to inoculate seedlings of plantain and banana genotypes, disease response of genotypes was rated by measuring the variables incubation period (IP), time of evolution of symptoms (TES), area under disease progress curve (AUDPC) and rate of disease development (r).The inoculation of fifty isolates on Dominico Harton cultivar allowed to establish five levels of virulence (very high, high, medium, low and very low), which were not related to their geographic origin nor to genotype of the isolates. Isolates with different virulence level were present in the same zone and in the same genotype. The resistance assay showed that plantain and banana genotypes can present a differential behavior to the M. fijiensis isolates. The disease severity measured in the genotypes allowed to classify the in three disease reactions levels, resistance, intermediate and susceptible genotypes The plantain genotypes Topocho, Maqueno, FHIA 20 and FHIA 21, and the banana genotype Sedita and FHIA 23 were characterized for having a higher level of resistance, that was expressed as a lower disease severity and slower disease progress as compared with the other genotypes.
Palabras clave: Musaceous, pathogenicity, virulence, resistance.
La Sigatoka negra ocasionada por el hongo Mycosphaerella fijiensis Morelet [anamorfo Paracercospora fijiensis (Morelet) Deigthon], es considerada hasta el momento, la enfermedad foliar de mayor importancia que afecta a los cultivos de plátano y banano en todo el mundo, produciendo pérdidas en rendimiento de hasta 100%, si no se implementan medidas para su manejo (Orozco y Aristizábal, 2006).
La enfermedad provoca desórdenes significativos en el crecimiento vegetativo de la planta, la cual sufre un severo deterioro del área foliar y de la productividad del cultivo, al disminuir su capacidad fotosintética. De presentarse esta condición, la planta no logra extraer de las hojas los elementos nutritivos para llevarlos al racimo y llenarlo; éste puede presentar madurez prematura y la fruta no sirve para la exportación (Herrera, 2007).
Las medidas de manejo de la enfermedad se han basado en el uso de productos químicos que aunque permiten enfrentar de forma eficaz la enfermedad, presenta desventajas por sus efectos sobre el medio ambiente, sumado a la resistencia de las poblaciones del patógeno, adquirida por la aplicación de ciertos fungicidas sistémicos muy utilizados como los benzimidazoles y más recientemente los triazoles (Vega, 2002).
La problemática de la Sigatoka negra va mas allá de las pérdidas que ocasiona y de las consecuencias de su manejo; hay que tener en cuenta que el éxito de la enfermedad también es debido a la alta variabilidad genética y patogénica de las poblaciones del hongo (Carlier et al., 1996; Fullerton y Olsen, 1995), como a la plantación de variedades susceptibles. Es claro que el esfuerzo para el manejo óptimo de la enfermedad debe ser idealmente preventivo, en tal sentido la plantación de genotipos resistentes se constituye en la estrategia preventiva y económica para el productor en el control eficiente de la enfermedad.
Dadas las consideraciones anteriores, se hace necesario realizar investigaciones como la del presente trabajo, cuyo objetivo principal fue evaluar bajo condiciones de invernadero genotipos de plátano y banano mediante el uso de aislamientos de M. fijiensis de diferente nivel de virulencia y origen, esto con el fin de identificar genotipos resistentes a las cepas colombianas del patógeno y conocer los que sirvan como fuentes de resistencia a la enfermedad, que puedan ser utilizados en futuros programas de mejoramiento genético.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en el laboratorio e invernadero del Programa de Patología de Yuca y Frutas Tropicales del Centro Internacional de Agricultura Tropical (CIAT), Palmira, bajo condiciones controladas de temperatura y humedad relativa.
Colección de aislamientos. Con el objetivo de conformar un banco de aislamientos de M. fijiensis se tomaron muestras de tejido foliar de cultivos establecidos y plantas individuales con síntomas de la enfermedad de la Sigatoka negra, procedentes de 10 departamentos de Colombia y diferentes genotipos de plátano y banano.
De las muestras colectadas se procedió a realizar el aislamiento del patógeno, siguiendo dos metodologías: descarga de ascosporas y siembra directa de tejido afectado. Cada metodología se llevó a cabo en cámara de flujo laminar y bajo condiciones asépticas utilizando material estéril.
La metodología de descarga de ascosporas (Stover, 1963), consistió en tomar la muestra que presentaba los síntomas más avanzados de la enfermedad e incubarla en cajas plásticas transparentes con papel toalla humedecida. Pasado el tiempo de incubación del tejido y con ayuda de un estereoscopio, se seleccionaron y cortaron zonas de aproximadamente 2 cm2 que presentaban buen número de peritecios. Cuatro fragmentos de hoja fueron adheridos por disco de papel filtro de 9 cm de diámetro. Posteriormente, cada papel filtro con los fragmentos de tejido se sumergieron en agua destilada estéril durante 5 min. Los discos de papel filtro se colocaron en tapas de cajas Petri que contenían PDA (papa-dextrosa-agar, 39 g/L de agua), de modo que el envés de las hojas quedara hacia el agar, ya que es el lugar donde se encuentran los peritecios y de esta manera promover mayor descarga de ascosporas sobre el medio de cultivo.
Después de 15 h de descarga, los discos de papel filtro con los fragmentos de tejido fueron retirados y con ayuda de un microscopio compuesto, se identificaron las ascosporas. Éstas eran elípticas, de 14 a 20 mm de largo y 4 a 6 mm de ancho, hialinas y usualmente con dos células (Belalcázar et al., 1991). Posteriormente en el estereoscopio y con una aguja de disección, se tomaron ascosporas individuales y se pasaron a cajas Petri con medio PDA. Los aislamientos se incubaron a 27 ºC durante 20 d en oscuridad para la producción de micelio. Una vez obtenido el micelio, con un bisturí se retiró parte de éste y se colocó en un vial para microcentrífuga con 1 mL de agua destilada estéril y se maceró hasta obtener una suspensión homogénea. Se tomaron 250 mL de la suspensión y se distribuyeron uniformemente en cajas Petri con medio agar V-8 modificado (100 mL de jugo V-8, 0,2 g de CaCO3 y 20 g de agar por litro de agua, pH 6). Los cultivos se incubaron a 20 ºC bajo luz blanca continua, durante 2 semanas para la producción de conidias.
La metodología de siembra directa de tejido afectado, consistió en cortar fragmentos de tejido de aproximadamente 15 mm2 de tal manera que incluyeran tejido enfermo como tejido sano. Los fragmentos fueron lavados con agua deionizada durante 45 min, y se desinfectaron en hipoclorito de sodio al 1% por 1 min y etanol al 70% por el mismo tiempo, finalmente se lavaron con agua destilada estéril (Castaño y del Rio Mendoza, 1997). Los fragmentos secos se transfirieron a cajas Petri con PDA y se incubaron a 24 ºC durante 25 d. Una vez esporulado el hongo, se tomaron conidias individuales que se transfirieron una por cada caja Petri con el mismo medio. Finalmente los aislamientos monospóricos se incubarón a 24 ºC en oscuridad durante 25 días para la producción de conidias.
Para la identificación de Paracercospora fijiensis (anamorfo de M. fijiensis), se observaron conidias en azul de lactofenol, el cual produce una coloración más intensa en la cicatriz (hilio) o punto de inserción de las conidias en el conidióforo, presente en P. fijiensis y ausente en Pseudocercospora musae, anamorfo de M. musicola, causante de la Sigatoka amarilla (Aguirre et al., 1998).
Los aislamientos monospóricos se conservaron en papel filtro colonizado por el hongo a 4 y -20 ºC (Aricapa y Correa, 1994) y en glicerol al 15% a -80 °C (Abadie et al., 2008).
Caracterización morfológica y patogénica de aislamientos. De la colección de aislamientos colombianos de M. fijiensis se seleccionaron 50 procedentes de diferente zona geográfica y genotipo hospedante, los cuales se caracterizaron morfológica y patogénicamente.
Caracterización morfológica. Para cada aislamiento se tomaron datos morfológicos de longitud, ancho, y número de septas de 30 conidias. Para dichas variables se realizó un análisis de varianza con un modelo de bloques complemente al azar. Por su parte, la caracterización morfológica de la colonia se hizo a los 35 d de crecimiento en medio de cultivo PDA, donde se evaluó el diámetro, la textura y el color de la colonia, y se utilizó un diseño de bloques completos al azar, con un disco de 3 mm de diámetro del aislamiento por caja Petri y tres repeticiones por aislamiento.
Caracterización patogénica. Para la producción de inóculo, los fragmentos de papel filtro colonizados por el hongo fueron reactivados mediante su siembra en cajas Petri con medio PDA. Las cajas fueron incubadas en oscuridad a 24 ºC por 25 d para la producción de conidias. La inoculación del hongo se hizo asperjando uniformemente una suspensión conidial de una concentración de 5.000 conidias mL-1, sobre el envés de las dos hojas más jóvenes abiertas en plántulas de Dominico Hartón (genotipo susceptible a la Sigatoka negra) con 3 meses de edad.
Se evaluaron las variables: periodo de incubación (PI), definido como el tiempo (días) desde el momento de la inoculación hasta la aparición de los primeros síntomas; tiempo de evolución de los síntomas (TES), registrado como el tiempo (días) desde la aparición de los primeros síntomas hasta el estado de mancha con centro necrosado, área bajo la curva del desarrollo de la enfermedad (ABCDE) y tasa de desarrollo (r) del progreso de la enfermedad.
Las variables PI y TES se determinaron con base en la escala de estadios de síntomas de la Sigatoka negra establecida por Aránzazu et al. (2002), y las variables ABCDE y la r se determinaron con base en el uso de los diagramas estándares de Stover modificados por Gauhl en 1990 para Sigatokas (citado por Carlier et al., 2002), donde: grado 0=sin síntomas, 1=menos de un 1% de área foliar afectada, 2=1 a 5% de área foliar afectada, 3=6 a 15% de área foliar afectada, 4=16 a 33% de área foliar afectada, 5=34 a 50% de área foliar afectada y 6=51 a 100% de área foliar afectada.
Se utilizó un diseño de bloques completos al azar, con 50 aislamientos (tratamientos) y cinco repeticiones, y con una plántula como unidad experimental. Para cada una de las variables se hizo un análisis de varianza y mediante un análisis de componentes principales se caracterizaron los aislamientos en términos de las variables evaluadas. Para determinar grupos de aislamientos de características similares, se hizo un análisis de clúster con el método de UPGMA (group average, unweighted pair-group method using arithmetic averages) y el criterio R2.
Evaluación de resistencia. Cinco aislamientos de diferente virulencia y origen fueron inoculados en 10 genotipos de plátano y nueve de banano. Las cepas a su vez, fueron elegidas teniendo en cuenta el departamento (zona de mayor producción) y genotipo (plátano y banano) del cual fueron aisladas.
Para la producción de inóculo se utilizó la misma metodología anteriormente mencionada en la prueba de patogenicidad. Así mismo, se evaluaron las variables PI, TES, ABCDE y r.
Se utilizó un diseño de parcelas divididas con tres repeticiones por tratamiento, como factor parcela principal, los aislamientos y como subparcela, los genotipos en un diseño completamente al azar. Se hizo un análisis de varianza para las variables PI, TES, ABCDE y r. Mediante el análisis de componentes principales se caracterizaron las interacciones (genotipo-aislamiento) en términos de las variables evaluadas. Para determinar grupos de interacciones de características similares, se hizo un análisis de clúster con el método de UPGMA y el criterio R2. Todos los datos fueron procesados mediante el programa estadístico Statistical Analysis System (SAS, versión 9).
RESULTADOS
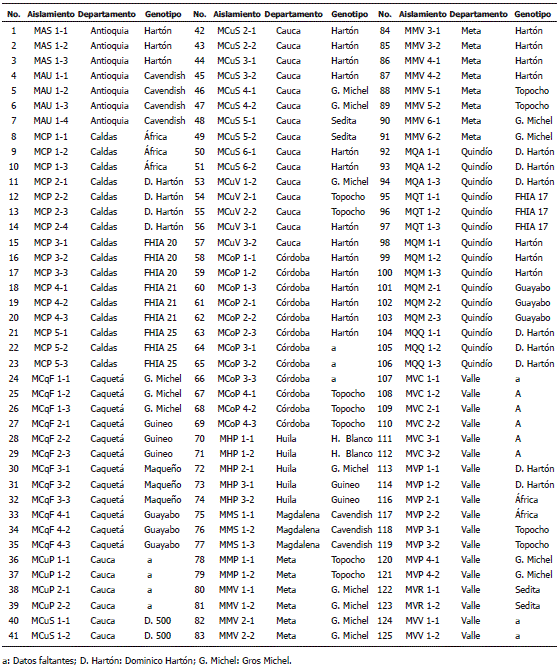
Colección y caracterización morfológica de aislamientos de M. fijiensis. Del procesamiento de las muestras foliares afectadas con síntomas típicos de la enfermedad de la Sigatoka negra, se conformó una colección de 125 aislamientos monospóricos de M. fijiensis (Tabla 1).
Tabla 1. Colección de aislamientos colombianos de Mycosphaerella fijiensis Morelet según su origen.
El análisis de varianza detectó diferencias altamente significativas (P<0,001) para las variables longitud, ancho y número de septas de las conidias para los 50 aislamientos seleccionados. La longitud de las conidias fluctuó entre 20 y 110 µm (CV 29,57%), el ancho entre 2,5 y 5 µm (CV 16,85%) y el número de septas entre 2 y 10 por conidia (CV 24,80%) (Tabla 2).
Tabla 2. Caracterización morfológica de 50 aislamientos de Mycosphaerella fijiensis Morelet.
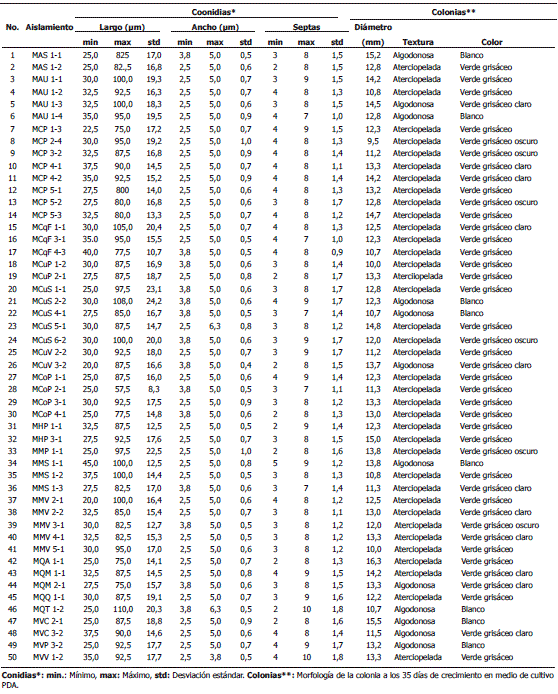
En la caracterización morfológica de la colonia a los 35 días de crecimiento en medio de cultivo PDA, se obtuvieron diferencias altamente significativas (P<0,001) para la variable diámetro. Mediante el criterio del valor crítico de Duncan (1,926) y el valor de la media general (12,72 mm) se conformaron tres grupos de aislamientos de acuerdo a su tamaño de colonia: Grupo A: aislamientos de colonias con diámetro entre 13,83 - 16,33 mm (media del grupo 14,68 mm). Grupo B: aislamientos de colonias con diámetro entre 12,00 - 13,82 mm (media del grupo 12,81 mm). Grupo C: aislamientos de colonias con diámetro entre 9,50 - 11,99 mm (media del grupo 10,74 mm). Según esta agrupación, los 50 aislamientos se distribuyen en un 24% en el grupo A, 50% en el grupo B y 26% en el grupo C (Tabla 2).
Con respecto al color de la colonia, es de anotar que en general para todos los aislamientos el color varió a través de los 35 días del experimento, el cual inicialmente fue verde grisáceo oscuro, luego verde grisáceo, verde grisáceo claro y finalmente blanco. Para el día 35, el 12% de los aislamientos tenían color verde grisáceo oscuro, el 48% verde grisáceo, el 22% verde grisáceo claro y el 16% blanco. En cuanto a la textura, en general las colonias pasaron de tener una textura aterciopelada a algodonosa para el día 35, el 76% de los aislamientos fueron aterciopelados y el 24% fueron algodonosos.
Se pudo observar que la textura de las colonias guarda relación directa con el color de las mismas, ya que cuando las colonias tenían un aspecto aterciopelado su color era verde grisáceo o con las diferentes tonalidades ya mencionadas para el mismo color y cuando las colonias tomaban un aspecto algodonoso su color se tornaba blanco (Tabla 2).
Caracterización patogénica de aislamientos de M. fijiensis. El análisis de varianza para cada una de las variables evaluadas (PI, TES, ABCDE y r) mostró diferencias altamente significativas (P<0,001) entre los aislamientos, lo que indica que hubo diferencias en el progreso de la enfermedad.
Mediante el análisis de componentes principales (ACP) se obtuvo la recodificación de los aislamientos en un nuevo sistema ortogonal. Con dicha información traducida y con el método UPGMA y el criterio de R2 se determinaron 8 grupos o clúster, cada uno de los cuales agrupó los aislamientos de características más similares entre sí. Para cada grupo constituido se calculó el promedio de cada variable, el cual se calificó dentro de una escala diseñada con base en los rangos obtenidos de PI, TES, ABCDE y r de todos los aislamientos (Tabla 3).
Tabla 3. Calificación de las variables PI, TES, ABCDE y r, para los grupos de aislamientos de Mycosphaerella fijiensis Morelet conformados del análisis de componentes principales (ACP) y análisis de clúster con el método UPGMA.
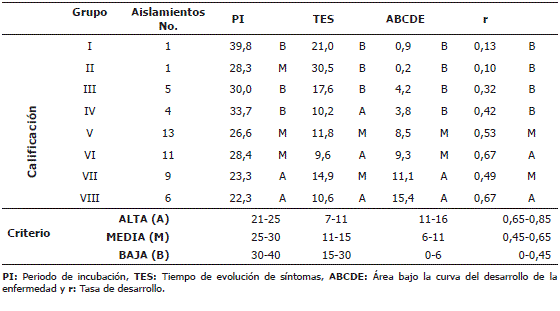
Las calificaciones de cada una de las variables dentro y entre los grupos conformados, permitió su distribución en cinco niveles de virulencia (muy alta, alta, media, baja y muy baja) (Tabla 4). Los aislamientos pertenecientes a los grupos I y II se consideran de muy baja virulencia, los aislamientos del grupo III de baja, los aislamientos del grupo V de media y los aislamientos del grupo VIII se consideran de muy alta virulencia.
Tabla 4. Virulencia de 50 aislamientos de Mycoshaerella fijiensis Morelet inoculados en plántulas de plátano Dominico Hartón en invernadero.
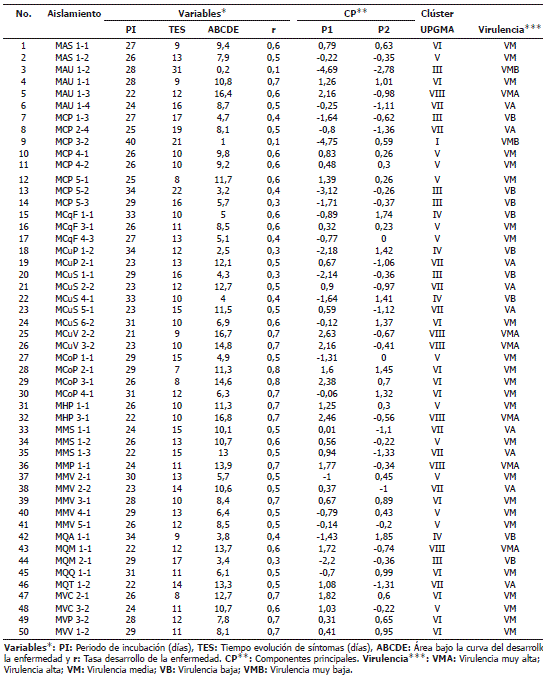
Los aislamientos que conformaron los grupos IV, VI y VII aunque estadísticamente difieren de los demás, presentaron tendencia a una virulencia baja, media y alta respectivamente.
Es de mencionar que todos los aislamientos mostraron diferentes niveles de virulencia y que ninguno fue avirulento o no produjo enfermedad en Dominico Hartón. A su vez, no se pudo establecer ninguna relación entre las características morfológicas y la virulencia de los aislamientos, ni relación entre el origen geográfico o varietal (plátano o banano) de los mismos con su virulencia.
Evaluación de resistencia en genotipos de plátano y banano. El análisis de varianza para las variables evaluadas mostró en todos los casos, interacciones altamente significativas (P<0,001) entre los aislamientos y los genotipos; lo cual indica el comportamiento diferencial de los genotipos frente a cada uno de los aislamientos inoculados.
Mediante el análisis de componentes principales se obtuvo la recodificación de las interacciones (genotipo-aislamiento) en un nuevo sistema ortogonal; con dicha información traducida y con el método UPGMA y el criterio de R2 se determinaron seis grupos o clúster, cada uno de los cuales agrupó las interacciones de características más similares. Para cada grupo constituido se calculó el promedio de cada variable, el cual se calificó dentro de una escala diseñada con base en los rangos obtenidos de PI, TES, ABCDE y r (Tabla 5).
Tabla 5. Calificación de las variables PI, TES, ABCDE y r para grupos de interacciones (genotipo-aislamiento), conformados del análisis de componentes principales (ACP) y análisis de clúster con el método UPGMA.
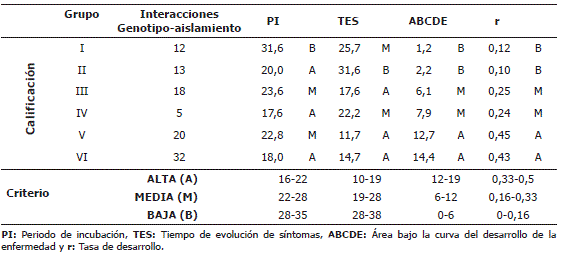
Las calificaciones dentro y entre los grupos de interacciones conformados permitieron su distribución en tres niveles de reacción (resistente - intermedio – susceptible). Los grupos de interacción I y II mostraron reacción resistente, III y IV reacción intermedia, y V y VI reacción susceptible. Es de mencionar que los grupos de interacciones que conformaron cada nivel de reacción difirieren entre sí por el valor del PI, lo cual indica que dicha variable no es determinante para el progreso de la enfermedad, ya que las plantas pueden expresar los síntomas tempranamente pero la enfermedad puede evolucionar de forma lenta. Las variables ABCDE y r, si explican bien la conformación de los grupos de interacción genotipo – aislamiento (Tabla 5).
A razón de que los grupos conformados son de interacciones y no de genotipos, un mismo genotipo puede estar presente en varios grupos debido a su diferente reacción, ante el conjunto de aislamientos utilizados.
Reacción de los genotipos Musa spp. frente a los aislamientos de M. fijiensis. La Tabla 6 muestra la susceptibilidad de muchos de los genotipos a la mayoría de los aislamientos, en ese sentido los genotipos de plátano África y los bananos Gross Michel, Giant y Valery Cavendish se comportaron de manera susceptibles a todos los aislamientos evaluados; otros genotipos de plátano como FHIA 20, Topocho, Maqueño, Dominico y Cubano negro, se comportaron de manera diferencial ante los aislamientos, presentando resistencia para al menos a uno de ellos, mientras que en banano este mismo comportamiento solo se observó en tres genotipos FHIA 23, Sedita y Gran Enano. El único genotipo en presentar resistencia a todos los aislamientos fue el plátano FHIA 21, el cual aún conserva su alta resistencia a la Sigatoka negra bajo condiciones de campo.
Tabla 6. Reacción de 19 genotipos Musa spp. frente a cinco aislamientos de Mycosphaerella fijiensis Morelet de diferente virulencia y origen.
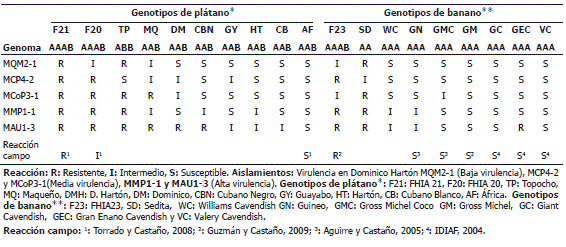
Por otra parte, la comparación de las reacciones de cada uno los aislamientos ante el conjunto de genotipos evaluados (comparación de filas Tabla 6) muestra un comportamiento diferencial de la virulencia del patógeno. Los aislamientos MQM2-1, MCP4-2 y MCoP3-1 presentaron la mayor virulencia debido a que afectaron la mayor cantidad de genotipos (13, 12 y 11 respectivamente); esto a diferencia del aislamiento MAU1-3 que evidenció una menor virulencia ya que sólo 5 genotipos expresaron susceptibilidad a esta cepa. El aislamiento MMP1-1 presenta una virulencia media debido a que 9 genotipos fueron susceptibles a este aislamiento. Esto es importante ya que si se observa la reacción de virulencia de éstos aislamientos en Dominico Hartón (Ensayo de patogenicidad), es claro que el comportamiento de alta o baja virulencia de una cepa específica frente a un solo genotipo, no indica que dicho comportamiento sea el mismo frente a cualquier otro genotipo donde se evalúe, esto queda claramente confirmado con el aislamiento MQM2-1 quien presentó una baja virulencia en Dominico Hartón pero que al evaluarlo frente a una mayor gama de genotipos, resultó ser el de mayor virulencia, y a su vez, con el aislamiento MAU1-3 quien presentó una alta virulencia en Dominico Hartón y fue el menos virulento ante los diferentes genotipos de estudio.
DISCUSIÓN
Colección y caracterización de aislamientos de M. fijiensis. La ausencia de contaminación en todos los aislamientos a partir del método de siembra directa de tejido afectado, en comparación con el método de descarga de ascosporas, en el cual se tuvo grandes contaminaciones en el momento de la obtención de los monoascospóricos; muestra que la siembra directa de tejido afectado es una metodología sencilla y efectiva para el aislamiento del hongo. Este resultado conlleva a reducir la dificultad previamente reportada para aislar M. fijiensis, presentándose como una mejor alternativa a la metodología que tradicionalmente se utiliza para el aislamiento del patógeno, consistente en la descarga de ascosporas (Du Pont, 1982; Abadie et al., 2008; Jiménez, 2008).
La conservación del hongo en papel filtro a temperaturas bajas resultó ser un método eficiente para el almacenamiento y para la posterior inducción de esporulación. En la literatura no se encuentran estudios previos de conservación del hongo mediante esta metodología, lo que hace que esta investigación se constituya en la primera referencia de conservación de M. fijiensis en papel filtro y en una metodología efectiva de almacenamiento y producción de inóculo.
Los resultados de la caracterización morfológica de los aislamientos, mostraron rangos en longitud, ancho y número de septas de las conidias que concuerdan con los indicados en la literatura para la identificación del hongo (Ploetz et al., 1994; Aguirre et al., 1998; Jiménez, 2008). En cuanto a las características de diámetro, color y textura de las colonias, coinciden con los informes previos de Manzo et al. (2001) y Jiménez (2008) para aislamientos de M. fijiensis.
Caracterización patogénica de aislamientos. La reacción observada de Dominico Hartón ante los 50 aislamientos evaluados (Tabla 4), infiere que todas la interacciones planta-patógeno son de carácter compatible, el 78% de los aislamientos tuvieron una compatibilidad entre media, alta y muy alta, lo que demuestra que este genotipo no posee genes de resistencia a las poblaciones colombianas de M. fijiensis. Esto se correlaciona con la reacción de susceptibilidad de este genotipo en campo observada por Hoyos y Castaño (2007) en Colombia y por Hernández et al. (2006) en Venezuela.
Este estudio, a su vez muestra que la estructura poblacional del hongo es muy variable ya que la virulencia de las cepas de M. fijiensis en Dominico Hartón no tiene ninguna asociación con su origen geográfico, ni con la especie de la cual fueron aisladas. Encontrándose en una misma zona geográfica, aislamientos de alta y baja virulencia, y a su vez, encontrando aislamientos de diferente agresividad provenientes de un mismo genotipo. Lo que se correlaciona con la alta diversidad genética encontrada en aislamientos de M. fijiensis obtenidos en una misma plantación e incluso entre aislamientos de una misma planta (Rivas et al., 2004). Esto se asocia con la reproducción sexual y ciclo de vida corto del patógeno, que provoca un elevado índice de recombinación genética (Manzo et al., 2005), y conlleva a una rápida aparición de poblaciones del hongo con nuevos genes de virulencia.
La alta variabilidad patogénica del M. fijiensis en Dominico Hartón demuestra la necesidad de evaluar a nivel de invernadero diferentes genotipos de Musa spp. frente a una diversa y amplia gama de aislamientos del hongo que representen su variación patogénica y genética real, que permita reconocer genotipos resistentes a las cepas del colombianas del patógeno, y a su vez, facilite la identificación de fuentes de resistencia confiables a la Sigatoka negra.
Evaluación de resistencia en genotipos de plátano y banano. Las interacciones planta-patógeno observadas en el complejo Musa spp. y M. fijiensis se manifestaron de acuerdo a la hipótesis del gen x gen de Flor (1971), donde la resistencia resultó de la interacción genética entre los genes de resistencia de la planta y los genes de avirulencia del patógeno; la ausencia, mutación o pérdida de uno de estos genes en cualquiera de los dos organismos (planta o patógeno) es determinante para que la interacción deje de ser de resistencia y se manifieste como susceptible. En ese sentido, la susceptibilidad o interacción de compatibilidad observada en los genotipos África, Gross Michel, Giant y Valery Cavendish a todas las cepas utilizadas, demuestra que ninguno de estos genotipos posee los genes de resistencia asociados a los genes de virulencia de los aislamientos utilizados. El comportamiento de estos genotipos también explica la reacción de susceptibilidad en campo observada en los genotipos África y Gross Michel en Colombia (Hoyos y Castaño, 2007), y de Giant y Valery Cavendish en República Dominicana (IDIAF, 2004; Galván y Rengifo, 2004), e indica su condición de susceptibilidad a la enfermedad en cualquier zona de Colombia donde se cultiven o deseen cultivar.
Por otra parte, el comportamiento diferencial de los genotipos Musa spp. a las diferentes cepas del hongo, muestra mayores reacciones de resistencia en los genotipos de plátano que en los bananos, esto confirma lo mencionado por Tezenas (1990) y Belalcázar et al. (1991), quienes afirman que la resistencia a enfermedades en Musaceas es mayor en aquellos materiales con genoma B (Balbisiana “Plátanos”) que aquellos cultivares con genoma A (Acuminata “Bananos”); no obstante, también declaran que los cultivares diploides con genoma AA a pesar de no poseer genoma B, presentan características de alta resistencia al patógeno; y que aquellos genotipos con alta frecuencia de genoma B, es decir ABB expresan mayor resistencia que cultivares con genoma AAB. Esto es importante, ya que si se observa la Tabla 6, se identifican mayores reacciones de resistencia en las variedades de plátano Topocho con genoma ABB y banano Sedita con genoma AA que en los demás genotipos con genoma AAA y AAB. Sin embargo, aunque Maqueño presenta genoma AAB resultó ser un material interesante debido a que no exterioriza reacción de susceptibilidad ante ninguno los aislamientos evaluados. Este comportamiento lo convierte al igual que a Topocho y Sedita en un material promisorio tanto para el manejo preventivo de la enfermedad como para estudios de mejoramiento genético.
Por otro lado, las mayores reacciones de resistencia de los genotipos FHIA (Tabla 6) en comparación con las respuestas de los demás genotipos ante el conjunto de aislamientos del patógeno evaluados, confirman su resistencia de carácter horizontal (FHIA, 2008), la cual se adapta a la variabilidad patogénica de los aislamientos colombianos de M. fijiensis. Las mayores reacciones de resistencia de FHIA 21 que en FHIA 20, concuerdan con la resistencia en campo observada por Torrado y Castaño (2008). Las reacciones de resistencia en FHIA 23 a las cepas colombianas del patógeno aquí detectadas, son también importantes a razón de que este banano ha demostrado ser resistente a las Sigatokas negra y amarilla en condiciones de campo (Guzmán y Castaño, 2009).
Las reacciones observadas de los distintos genotipos a las diferentes cepas del patógeno, indican la alta diversidad genética de los genotipos de Musa spp. y evidencian la alta variabilidad en la virulencia de M. fijiensis, ya que todos los aislamientos presentaron comportamiento diferencial ante el conjunto de genotipos evaluados. Esto coincide con lo planteado por Fullerton y Olsen (1995) quienes utilizaron 63 aislamientos del patógeno, obtenidos de diferentes hospedantes de plátanos y bananos procedentes de países productores de Asía y América; en donde una vez inoculados los aislamientos, se demostraron diferencias en su virulencia y en el progreso de la enfermedad en los 20 genotipos estudiados.
Los genotipos Topocho, Maqueño y Sedita, y los híbridos FHIA 20, FHIA 21 y FHIA 23 que presentaron mayores reacciones de resistencia a las cepas colombianas de M. fijiensis podrían constituirse en fuentes de resistencia en programas de mejoramiento genético o en materiales promisorios para control de la enfermedad; teniendo en cuenta que antes de introducir genotipos resistentes identificados en este y otros estudios de detención temprana, se deberían evaluar ante un mayor número de aislamientos y bajo condiciones naturales como validación final de dicho comportamiento, e igualmente evaluarlos frente a otras características como producción y respuesta a otros problemas fitosanitarios, que también deben ser consideradas al momento de realizar un elección del material a cultivar.
CONCLUSIONES
La siembra directa de tejido afectado en medio de cultivo PDA, resultó ser un método muy útil y efectivo para el aislamiento del patógeno y para la obtención de cultivos monospóricos en comparación con el método de descarga de ascosporas. El método de conservación en papel filtro mostró ser una herramienta eficiente para el almacenamiento del hongo por largos periodos de tiempo, además de ser importante porque mantiene la esporulación de los aislamientos y conserva las características de crecimiento y virulencia de los mismos. Se encontraron diferencias en la morfología y virulencia de 50 aislamientos de Mycophaerella fijiensis inoculados en plántulas de plátano Domino Hartón. La virulencia de los aislamientos M. fijiensis no se relacionó con su origen geográfico ni con el genotipo del cual se obtuvieron. Los genotipos de plátano y banano mostraron un comportamiento diferencial frente a cinco aislamientos, lo cual indica la presencia de razas del patógeno. Los genotipos Topocho, Maqueño, FHIA 20, FHIA 21 de plátano, y los genotipos Sedita y FHIA 23 de banano presentaron un mayor nivel de resistencia. El método de evaluación temprana de resistencia a M. fijiensis en cultivares de plátano y banano, es confiable y reproducible, ya que permite evaluar respuestas marcadas según el nivel de resistencia y susceptibilidad del cultivar; esto representa un ahorro en tiempo y recursos en comparación con las pruebas hechas a nivel de campo.
AGRADECIMIENTOS
Se agradece a Germán Alberto Llano, Girlena Aricapa y Myriam Cristina Duque, por su valioso conocimiento brindado y orientación. A Silverio González Director de FEDEPLATANO, Luis Alfredo Rivera, Vicente Rey, Pedro Zapata, Carlos Jara, entre otros, por apoyar esta investigación, por medio de la obtención de muestras, germoplasma y sobre todo por la ayuda desinteresada y conocimientos.
BIBLIOGRAFÍA
Abadie, C., M. Zapater, L. Pignolet, J. Carlier and X. Mourichon. 2008. Artificial inoculation on plants and banana leaf pieces with Mycosphaerella spp., responsible for Sigatoka leaf spot diseases. Fruits 63(5): 319–323.
Aguirre, M., J. Castaño y L. Zuluaga. 1998. Método rápido de diagnóstico de Mycosphaerella musicola Leach y M. fijiensis Morelet agentes de la Sigatoka amarilla y Sigatoka negra en Musáceas. Fitopatología Colombiana 23(1): 45-47. [ Links ]
Aguirre, M. y J. Castaño. 2005. Epidemiología de Mycosphaerella fjiensis Morelet y Mycosphaerella musicola Leach, en siete genotipos de musáceas. Fitopatología Colombiana 29(1): 7-11. [ Links ]
Aránzazu, F., J. Valencia, M. Arcila, C. Castrillón, M. Bolaños, P. Castellanos, J. Pérez, y J. Rodríguez. 2002. El cultivo de plátano. Corporación Colombiana de Investigación Agropecuaria (CORPOICA), Litoas, Manizales, Colombia. 114 p. (Manual Técnico). [ Links ]
Aricapa, M. y F. Correa. 1994. Almacenamiento de hongos sobre papel filtro. Programa Patología de Arroz. CIAT. Ascolfi Informa, Boletín bimestral 20(3): 29-30. [ Links ]
Belalcázar, S., J. Toro y R. Jaramillo. 1991. El cultivo del plátano (Musa AAB Simmonds) en el trópico. Instituto Colombiano Agropecuario (ICA), Subgerencia de Investigación; Centro Internacional de Investigaciones para el Desarrollo (CIID). Red Internacional para el Mejoramiento del Banano y el Plátano (INIBAP), Cali, Colombia. 376 p. [ Links ]
Carlier J., M. Lebrun, M. Zapater, C. Dubois and X. Mourichon. 1996. Genetic structure of the global population of banana black leaf streak fungus Mycosphaerella fijiensis. Molecular Ecology 5(4): 499-510. [ Links ]
Carlier, J., D. De Waele and J. Escalant. 2002. Global evaluation of Musa germplasm for resistance to Fusarium wilt, Mycosphaerella leaf spot diseases and nematodes. (A. Vézina and C. Picq, eds). INIBAP Technical Guidelines 6. The International Network for the Improvement of Banana and Plantain, Montpellier, France. 68 p. [ Links ]
Castaño, J. y L. Del Río Mendoza. 1997. Manual para el diagnóstico de hongos, bacterias, viru s y nematodos fitopatógenos. Zamorano Academic Press, Universidad de Caldas, Facultad de Ciencias Agropecuarias, Manizales, Colombia. 210 p. [ Links ]
Du Pont. 1982. Black and yellow sigatoka. Improyed identification and management. Du Pont Latin América, Coral Gables, Florida. 17 p. [ Links ]
Flor, H.H. 1971. Currents status of the gene-for-gene concept. Annual Review of Phytopathology 9: 275-296. [ Links ]
Fundación Hondureña de Investigación Agrícola (FHIA). 2008. Programas de Investigación. En: Banano y plátano, http://www.honduras.com/fhia/default-esp.htm 1p.; consulta: agosto 2009. [ Links ]
Fullerton, R.A. and T.L. Olsen. 1995. Pathogenic variability in Mycosphaerella fijiensis Morelet cause of black Sigatoka in banana and plantain. New Zealand Journal of Crop and Horticultural Science 23(1): 39-48. [ Links ]
Galván, E. y D. Rengifo. 2004. Evaluación de bananos locales e introducidos en condiciones de producción orgánica. pp. 19-26. En: Instituto Dominicano de Investigaciones Agropecuarias y Forestales (IDIAF). (ed.). Musáceas, resultados de investigación. Santo Domingo, República Dominicana. 117 p. [ Links ]
Guzmán, O.A. y J. Castaño. 2009. Reacción de los híbridos de plátano FHIA-20 y de banano FHIA-23 a las Sigatokas negra (Mycosphaerella fijiensis Morelet) y amarilla (Mycospaherella musicola Leach). Agronomía 17(1):35-43. [ Links ]
Hernández, Y., F. Portillo, M. Portillo, C. Navarro, M. Rodríguez y J. Velazco. 2006. Densidad estomática en materiales de plátano (Musa AAB, AAAB y ABB) susceptibles y resistentes a Sigatoka negra (Mycosphaerella fijiensis Morelet). Revista de la Facultad de Agronomía 23(3): 292-297. [ Links ]
Herrera, M. 2007. Manejo y control de la Sigatoka negra en plátano y banano. Asociación de Ingenieros Agrónomos del Valle de Cauca. Revista ASIAVA 77: 12-15. [ Links ]
Hoyos, J.E. y J. Castaño. 2007. Evaluación de resistencia de Mbouroukou (África 1) y FHIA 03 a la Sigatoka negra (Mycosphaerella fijiensis Morelet) y Sigatoka amarilla (Mycosphaerella musicola Leach). Agronomía 15(2): 67-76. [ Links ]
Instituto Dominicano de Investigaciones Agropecuarias y Forestales (IDIAF). 2004. Musáceas, resultados de investigación. Santo Domingo, República Dominicana. 117 p. [ Links ]
Jiménez, I. 2008. Effect of the nutritional status of banana (Musa spp.) on leaf disease infestation by Mycosphaerella fijiensis Morelet in Ecuador. Tesis postgrado. Katholieke Universiteit Leuven, Division of Crop Biotechnics, Bélgica. 149 p. [ Links ]
Manzo, G., M. Santos y G. Guzmán. 2001. Caracterización morfológica de Mycosphaerella fijiensis Morelet de la región del Pacífico centro de México y su desarrollo en medios de cultivo. Revista Mexicana de Fitopatología 19(1): 66-71. [ Links ]
Manzo, G., G. Guzmán, C. Rodríguez y A. James. 2005. Biología de Mycosphaerella fijiensis Morelet y su interacción con Musa spp. Revista Mexicana de Fitopatología 23(1): 87-96. [ Links ]
Orozco, M.L. y M. Aristizábal. 2006. Manejo de las sigatokas del plátano (Musa AAB) y su relación con el clima. Universidad de Caldas. Departamento de Fitotecnia. Resumen de investigación. Boletín Fitotecnia No. 110. [ Links ]
Ploetz, R.C., G.A. Zentmyer, W.T. Nishijima, K.G. Rohrbach and H.D. Ohr. 1994. Part I. Banana. pp. 2-22. In: Compendium of Tropical Fruit Diseases. APS Press. The American Phytopathological Society, St. Paul, MN, USA. 88 p. [ Links ]
Rivas, G., M. Zapater, C. Abadie and J. Carlier. 2004. Founder effects and stochastic dispersal at the continental scale of the fungal pathogen of bananas Mycosphaerella fijiensis. Molecular Ecology 13(2): 471-482. [ Links ]
Statistical Analysis System (SAS) Institute Inc. 2004. SAS Online Doc, Versión 9, Cary, NC. [ Links ]
Stover, R.H. 1963. Leaf spot of bananas caused by Mycosphaerella musicola: Associated ascomycetous fungi. Canadian Journal of Botany 41(10): 1481-1485. [ Links ]
Tezenas du Montcel, H. 1990: Susceptibility of various cultivated bananas to Sigatoka diseases. pp. 272-289 In: Fullerton, R. and R. Stover (ed.). Sigatoka leaf diseases of bananas. Proceedings of an international workshop held at San Jose, Costa Rica, March 28-April 1 1989. International Network for the Improvement of Banana and Plantain, Montpellier, France. [ Links ]
Torrado, M. y J. Castaño. 2008. Incidencia y severidad de las sigatokas negra (Mycosphaerella fijiensis Morelet) y amarilla (Mycosphaerella musicola Leach et Mulder) del plátano según los estados fenológicos. Agronomía Colombiana 26(3): 435-442. [ Links ]
Vega, G. 2002. La Sigatoka negra (Mycosphaerella fijiensis Morelet) en el plátano. Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, Granma. Cuba.

