










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em
SciELO
Similares em
SciELO -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
versão impressa ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín v.64 n.1 Medellín jan./jun. 2011
REVISIÓN: DEGRADACIÓN DE PLAGUICIDAS MEDIANTE HONGOS DE LA PUDRICIÓN BLANCA DE LA MADERA
PESTICIDES DEGRADATION BY WHITE ROT FUNGI: A REVIEW
Juan Carlos Quintero Díaz1
1 Profesor Asociado. Universidad de Antioquia. Departamento de Ingeniería Química. Grupo de Bioprocesos. A.A. 1226. Medellín, Colombia. < jcquinte@udea.edu.co>
Recibido: Agosto 06 de 2009 ; Aceptado: Febrero 22 de 2011.
Resumen. Los hongos de la pudrición blanca de la madera, se han caracterizado por su capacidad para degradar y mineralizar la lignina empleando un sistema enzimático extracelular compuesto principalmente de tres enzimas Ligninoperoxidasa (LiP), Manganeso peroxidasa (MnP) y Lacasa. Durante los últimos veinte años se ha orientado la atención a estos hongos y su sistema enzimático ligninolítico para estudiar la capacidad para degradar un amplio rango de compuestos xenobióticos como plaguicidas, colorantes, explosivos, etc. Sin embargo, se ha observado que gran número de compuestos entre ellos los plaguicidas no responden al proceso degradativo de las enzimas ligninoliticas y esto ha permitido descubrir recientemente nuevos mecanismos empleados por estos hongos como son los sistemas oxidativos de las monooxigenasas del citocromo P-450 y reductivo de las transferasas, ya ampliamente conocidos en animales superiores e identificados como fase I y fase II del metabolismo. En esta revisión se describen estos tres tipos de mecanismos degradativos hasta ahora conocidos que son empleados por los hongos para la degradación de contaminantes ambientales y se analizan algunos casos de plaguicidas donde se involucran estos mecanismos en su degradación.
Palabras clave: Hongos ligninolíticos, biodegradación, enzimas ligninolíticas.
Abstract. Wood white rot fungi are characterized by their capacity of degradation and mineralization of lignin by means of an enzymatic extracellular system, which mainly consists of lignin peroxidase (LiP), Manganese peroxidase (MnP) and Laccase. During the last twenty years, these fungi and their enzymatic ligninolytic system have been the focus of attention to study the degradation capacity of a wide range of xenobiotics as pesticides, dyes, explosives, etc. However, a large number of xenobiotics are not responding to ligninolytic enzymes biodegradation process. This situation has permitted the discovering of new mechanisms used by fungi as citochrome P-450 monooxygenases oxidation system, and transferases’ reductive system, widely identified in phase I and II of superior animals’ metabolism. The tree types of known degradation mechanisms used by fungi in environmental contaminants degradation and some other examples of degradation mechanisms in pesticides will be described and analyzed in this review.
Key words: White rot fungi, biodegradation, ligninolytic enzymes.
Las plagas son responsables de más del 40% de las pérdidas de la producción potencial de alimentos durante su producción y de un 20% tras las cosechas (Paoletti y Pimentel, 2000). Ante esta situación, los plaguicidas químicos han proporcionado una mejora significativa en la producción agrícola, sin embargo, también han ejercido un importante efecto adverso en el ambiente debido a su toxicidad, bioacumulación y persistencia en suelo, agua y aire. La mayor persistencia se observa en el suelo y se encuentra entre 3 y 10 años para el DDT (diclorodifeniltricloroetano) y en más de 11 años para el HCH (hexaclorociclohexano) (Dua et al., 2002).
El uso masivo de los plaguicidas químicos data de la década de los 40, siendo los organoclorados DDT y HCH los más empleados para el control de plagas agrícolas y vectores de enfermedades como el paludismo y el tifus. Se ha estimado que desde esa fecha se produjeron en el mundo más de 3.000.000 t de DDT (Wong et al., 2005) y 10.000.000 t de HCH (Li, 1999). Estos dos plaguicidas contabilizaron cerca del 80% del total de los plaguicidas producidos en los años 1950-1970.
En mayo de 2001, en el marco de la “Convención de Estocolmo sobre Contaminantes Orgánicos Persistentes (POPs)”, 127 países del mundo firmaron un tratado con el fin de proteger la salud humana y el ambiente de los efectos nocivos de los POPs, siendo su primer objetivo prohibir y eliminar 12 de los POPs más tóxicos a los que denominaron la “docena sucia” que incluye entre ellos a nueve plaguicidas: aldrín, endrín, dieldrín, heptachlor, chlordano, mírex, toxafeno, DDT y hexachlorobenceno (HCB).
La aplicación de los plaguicidas y los residuos generados, su mal manejo y disposición incontrolada, son las principales causas de contaminación ambiental, principalmente del recurso suelo, debido a su elevada adsorción sobre la materia orgánica, su baja volatilidad y estabilidad química, que les confiere un bajo grado de movilidad y una elevada persistencia. A pesar de estas características, la volatilización y la disolución en cuerpos de agua son importantes mecanismos de dispersión de la contaminación (Semple et al., 2003).
Existen diferentes alternativas para el tratamiento de residuos sólidos y líquidos contaminados con POPs, siendo las más usadas las tecnologías físico-químicas como la incineración, la desorción térmica y la oxidación foto catalítica, Kearney y Roberts (1998); LaGrega et al. (1996). En los últimos años, las nuevas tecnologías han hecho énfasis en el desarrollo y aplicación de estrategias de remediación sostenibles. Estas estrategias, denominadas en su conjunto como “tecnologías de biorremediación”, son menos costosas y menos destructivas del medio comparadas con las tecnologías de tratamientos físicos y químicos (Atlas y Unterman, 1999). Tratamientos biológicos off-site, de suelos contaminados muestran costos promedio de 170 euros/m3, con relación a tratamientos de disposición en rellenos, lixiviación o térmicos, que oscilan entre 200 y 300 euros/m3 (Summersgill y Scott, 2005).
La “biorremediación” es un tratamiento tecnológico de descontaminación que usa sistemas biológicos para catalizar la destrucción o transformación de componentes peligrosos en el ambiente (Atlas y Unterman, 1999). El empleo de plantas para la eliminación de contaminantes del suelo, se denomina comúnmente como fitorremediación mientras que el término biorremediación se ha limitado al empleo de microorganismos o sus enzimas.
Existe un gran número de trabajos de investigación en laboratorio y en campo, orientados a la degradación de plaguicidas empleando bacterias. Este amplio uso se debe a que son fácilmente cultivables y manipulables genéticamente, crecen rápidamente, y son capaces de usar los plaguicidas como fuente de nutrientes, aunque requieren de una previa adaptación al medio y a los contaminantes que van a degradar (Caplan, 1993; Liu y Suflita, 1993; Furukawa, 2003).
A partir de la década de los 80, se ha incrementado el interés acerca de la posibilidad de usar en biorremediación hongos de la pudrición blanca de la madera. Estos hongos pertenecen a la clase Basidiomicete y son conocidos por degradar la lignina de la madera. Uno de los primeros trabajos sobre el tema indica que estos hongos, pueden degradar una amplia variedad de contaminantes ambientales (Bumpus et al., 1985). Las principales ventajas de estos hongos son la tolerancia a concentraciones considerablemente altas de contaminantes y su capacidad para crecer a bajos valores de pH. Además, gracias a la extensión de sus hifas, puede alcanzar contaminantes en el suelo, que no son biodisponibles ni biodegradables para otros organismos, y dado que requieren de sustratos lignocelulósicos para su crecimiento, es posible adicionar a los sitios contaminados, residuos de muy bajo costo como viruta de madera, carozo de maíz o paja de trigo, para promover su crecimiento e incrementar la degradación de los contaminantes (Aust y Benson, 1993; Barr y Aust, 1994).
En esta revisión se presentan los diferentes mecanismos enzimáticos empleados por los hongos de la pudrición blanca de la madera para degradar contaminantes orgánicos persistentes como son los plaguicidas y se analizan los mecanismos que son usados por estos hongos en su biodegradación.
Mecanismos de biodegradación. Se conocen tres principales mecanismos enzimáticos que son empleados por los hongos de la pudrición blanca de la madera para degradar contaminantes ambientales, dos de tipo oxidativo y uno reductivo: i) sistema de degradación de la lignina, que realiza ataques oxidativos a moléculas orgánicas por medio de radicales libres generados por las enzimas ligninolíticas peroxidasas; ii) fase I del metabolismo, mecanismo oxidativo en que intervienen las enzimas citocromo P-450 monooxigenasas y iii) fase II del metabolismo donde un conjunto de enzimas cataliza reacciones de conjugación reduciendo los contaminantes. Estos mecanismos degradan o modifican los contaminantes sin ser empleados como substratos para su crecimiento, es decir, la degradación se hace por cometabolismo. Aún no se han desarrollado trabajos para evaluar si algunos compuestos intermediarios de la degradación de xenobióticos son empleados como substratos.
Enzimas ligninolíticas. Los hongos de pudrición blanca de la madera, son el más importante grupo de microorganismos responsables de la biodegradación del polímero natural más complejo que existe, la lignina (Kirk y Farrell, 1987). Como se mencionó anteriormente estos hongos degradan la lignina de la madera para acceder a la hemicelulosa y celulosa, los cuales son sus verdaderos sustratos primarios. Estos sustratos también son adecuados para mantener una adecuada producción de enzimas en procesos de biorremediación sobre suelos contaminados (Quintero et al., 2006). El metabolismo ligninolítico, generalmente denominado “sistema de degradación de lignina” (SDL) es inducido por deficiencia o limitación de nutrientes, principalmente nitrógeno y carbono (Bucke, 1998; Moreira et al., 2000).
El SDL es el primero y más importante sistema catabólico de degradación de xenobióticos que usan estos hongos. La acción de este sistema es estrictamente extracelular y en el intervienen un grupo de hemoproteinas extracelulares denominadas enzimas peroxidasas, conocidas como lignino peroxidasas (LiPs) y manganeso peroxidasas (MnP), un grupo de enzimas productoras de H2O2 (glioxal oxidasa, aryl alcohol oxidasa, entre otras), un grupo de oxidasas (lacasas), alcohol veratrílico, manganeso y ácidos orgánicos como oxálico o malónico.
Las MnP ejercen su acción oxidativa indirectamente a través de la formación de radicales Mn+3 a partir del Mn+2. El Mn+3 se estabiliza formando quelatos con ácidos carboxílicos (ej. oxálico, malónico, málico, tartárico, láctico) que actúan como mediadores difusibles de baja masa molecular eliminando electrones e hidrógenos inespecíficamente a moléculas orgánicas (Hofrichter, 2002). Las LiPs pueden ejercer por si misma la acción oxidativa o formar radicales libres a partir de algunos compuestos orgánicos como el alcohol veratrílico (AV) o el dimetoxibenceno (DMB) de manera análoga a como ocurre con las MnP (Cameron et al., 2000). Estos radicales libres, son sustancias altamente oxidantes que pueden difundirse y penetrar en matrices en las que las propias enzimas (MnP y LiP) no lo pueden hacer y así aumentar la biodisponibilidad de sustratos y xenobióticos implicados en este tipo de metabolismo microbiano. Los ciclos catalíticos de las enzimas peroxidasas (LiP y MnP) en su reacción con H2O2 para formar los radicales libres Mn+3 y AV+ se presentan a continuación:

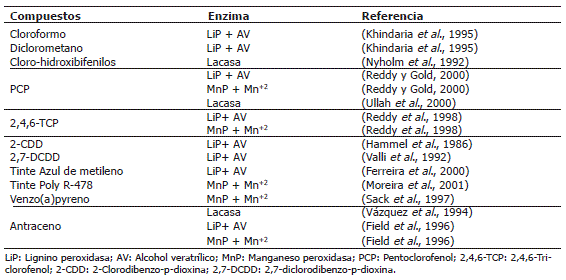
Se ha demostrado que el mecanismo oxidativo de las enzimas ligninolíticas, cataliza la degradación de gran cantidad de xenobióticos incluyendo hidrocarburos poliaromáticos, cloroaromáticos, dioxinas, tintes entre otros, que presenten potenciales de ionización (PI) ≤ 7,55 eV (Christian et al., 2005; Hammel et al., 1986). La enzima lacasa cataliza la oxidación de sustratos fenólicos y otros sustratos como antraceno y benzo(a)pyreno con PI ≤ 7,45 eV (Collins et al., 1996). Sobre este planteamiento, el conocimiento previo de los PI de contaminantes orgánicos de interés, por ejemplo determinándolo por voltametría cíclica (Riahi et al., 2007), permitiría saber si las enzimas ligninolíticas de estos hongos son o no útiles para su biodegradación. En el Tabla 1 se presentan algunos compuestos los cuales, se ha demostrado, son degradados por las enzimas ligninolíticas.
Tabla 1. Compuestos orgánicos degradados por enzimas ligninolíticas.
Ciertos co-oxidantes como compuestos orgánicos sulfurados (Glutatión (GSH), L-cisteina, etc.), así como ácidos grasos insaturados y sus derivados (ej. Ácido linoléico, Tween 80) son oxidados por el sistema de las MnP para formar radicales tiol y peroxil altamente reactivos (Figura 1), los cuales pueden atacar compuestos recalcitrantes que normalmente no son atacados directamente por las MnP, como son los hidrocarburos aromáticos policíclicos de entre tres y seis anillos (Bogan y Lamar, 1996; Hofrichter, 2002). Este mecanismo conocido como peroxidación, eleva la capacidad oxidativa del sistema ligninolítico a compuestos con PI hasta 8,1 eV (Bogan y Lamar, 1995).
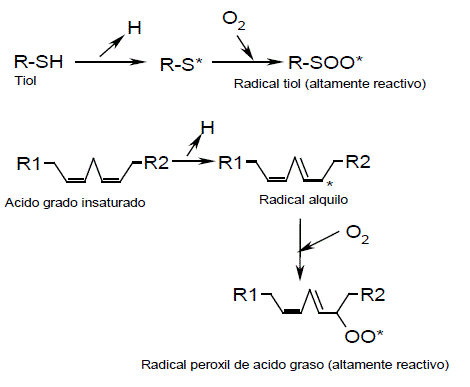
Figura 1. Formación de radicales reactivos mediados por las enzimas MnP (Hofrichter, 2002).
Muchos investigadores han relacionado la capacidad degradativa de los hongos de pudrición blanca con la actividad de su sistema enzimático ligninolítico; sin embargo, en numerosos trabajos se ha comprobado que estos hongos degradan xenobióticos (ej. clorofenoles, nitroaromáticos, plaguicidas) bajo condiciones de cultivo no ligninolíticas y además se ha comprobado que muchos de ellos no son degradados por las enzimas ligninolíticas en condiciones in vitro. Por otro lado, el número de trabajos en los que se atribuye la degradación de estos compuestos a sistemas enzimáticos diferentes al ligninolítico, en donde intervienen una serie de reacciones de oxidación, hidroxilación y reducción aumenta constantemente.
Varios de los metabolitos producidos por la degradación fúngica de contaminantes orgánicos que incluyen fenoles, epóxidos, dihidrodioles y quinonas, son producidos por reacciones similares a las conocidas farmacológicamente como fase I del metabolismo (Sutherland, 1992). En esta fase los procesos de biotransformación involucran las enzimas citocromo P-450 monooxigenasas y epóxido hidrolasa. En la fase II del metabolismo, los contaminantes orgánicos son conjugados con sulfato, ácido glucorónido, glutatión, u otras moléculas (Cerniglia et al., 1982). Las reacciones de las fases I y II son procesos bioquímicos que en plantas y animales superiores modifican la toxicidad de los contaminantes orgánicos o cambian su solubilidad en agua para ser eliminados del cuerpo.
Fase I del metabolismo. Las enzimas más importantes de la fase I son las citocromo P-450 monooxigenasas (P-450s) las cuales son una superfamilia de biocatalizadores que introducen un átomo de oxígeno en un amplio rango de moléculas (entre ellas moléculas contaminantes) para producir un epóxido.
En la Figura 2 se presenta su ciclo catalítico. Para el transporte de electrones interviene la enzima citocromo P-450 reductasa y su cofactor NADPH. Algunos epóxidos son inestables y a menudo sufren reacciones de reordenamiento a fenoles o sufren una hidratación enzimática vía otra enzima de la fase I, la epóxido hidrolasa generando trans-dihidrodioles (Jerina, 1983).
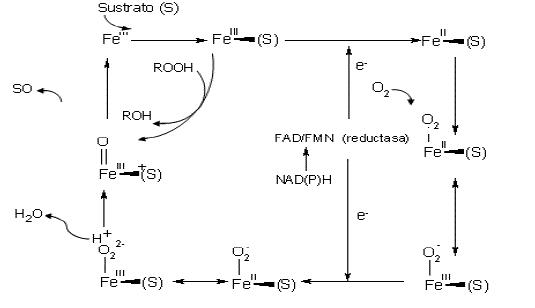
Figura 2. Ciclo catalítico de las enzimas citocromo P-450 monooxigenasas (Jerina,1983).
P-450s son un nombre colectivo dado para todas las hemo-proteinas que forman un complejo Fe(II)-CO con un espectro de absorción cercano a los 450 nm. Se sabe que estas enzimas oxidan más de 200.000 diferentes sustancias químicas exógenas y endógenas (Lewis, 2001; Urlacher y Eiben, 2006). Son altamente específicas en procariotes e inespecíficas en eucariotes (Sariaslani, 1991). A diferencia de las peroxidasas, que son oxidoreductasas con el peróxido de hidrógeno actuando como aceptor de electrones, las citocromos P-450 monooxigenasas, rompen el enlace covalente de la molécula de oxígeno (O2) y un átomo es incorporado a la molécula de sustrato mientras que el otro es reducido a agua (Wood, 1992). Las reacciones globales de los dos sistemas enzimáticos son:
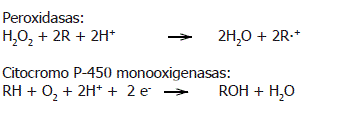
En algunos trabajos recientes relacionados con la secuenciación parcial del genoma del hongo de la pudrición blanca Phanerochaete chrysosporium, se indica que existen en este hongo al menos 97 genes para las P-450s (Ichinose et al., 2002a). Con esto se ha eliminando la idea previa de que estos hongos poseían una pobre diversidad genética para P-450s y por el contrario ha conducido a que se incremente el interés en su identificación y caracterización. En los últimos años, se ha identificado la presencia de enzimas de la fase I P-450s en algunos hongos de pudrición blanca como Phanerochaete chrysosporium, Bjerkandera adusta y Coriolus versicolor (Kullman y Matsumura, 1997; Eilers et al. 1999; Ichinose et al. 2002a; Ichinose et al., 2002b; Harshavardhan et al., 2005) y P-450s y epóxido hidrolasa en Pleurotus ostreatus (Bezalel et al. 1997). Cuando los tratamientos de biorremediación se llevan a cabo con los hongos de la pudrición blanca de la madera en condiciones no ligninolíticas, muchos investigadores han involucrado a las P-450s en la degradación de diferentes compuestos xenobióticos. Como ejemplos de ello se tienen: 2,4,6-TNT (Eilers et al. 1999), atrazina, benzo(a)pireno (Masaphy et al., 1996a; Masaphy et al., 1996b), lindano (Mougin et al., 1996), endosulfan (Kullman y Matsumura, 1996), fenantreno (Bezalel et al. 1997), difenil éter (Hiratsuka et al., 2001).
Fase II del metabolismo. Debido a que los hongos de pudrición blanca son generalmente reconocidos por su habilidad para oxidar contaminantes, el descubrimiento de que también causan deshalogenación reductiva de sustratos clorados fue una gran sorpresa. La primera observación de este fenómeno se hizo durante estudios para revelar las vías usadas por el Phanerochaete chrysosporium para degradar 2,4,6-triclorofenol y pentaclorofenol (Reddy y Gold, 1999; Reddy y Gold, 2001). Este mecanismo corresponde a la fase II del metabolismo.
La fase I del metabolismo puede actuar como intermediaria para que se lleven a cabo las reacciones de la fase II de carácter reductivo, las cuales son conocidas típicamente como vías de conjugación. Estas vías son mediadas por la glutatióntransferasa, sulfotransferasa y glucosyltransferasa entre otras; enzimas que adicionan glutatión, sulfato o residuos de azúcar a los xenobióticos, haciéndolos más solubles en agua, menos tóxicos y más fácilmente eliminables (Casillas et al. 1996; Field y Thurman, 1996). Posteriormente estos conjugados pueden ser reducidos por la enzimas conjugado reductasas generando compuestos más degradados que los xenobiócos originales (Reddy y Gold, 1999; Reddy y Gold, 2001).
Al igual que con las enzimas de la fase I, en los últimos años se han identificado, aislado y purificado las enzimas pertenecientes a la fase II Glutathion S-transferasa y glutatión conjugado reductasa de Phanerochaete chrysosporium involucradas en la dehalogenación reductiva del 2,4,6-triclorofenol, tetracloro-1,4-hydroquinona (Dowd et al., 1997; Reddy et al., 1998; Reddy y Gold, 1999; Reddy y Gold, 2001) y glutatión, sulfato y glucoronosyl transferasas de Pleurotus ostreatus (Bezalel et al., 1997).
Los mecanismos descritos muestran que los hongos de la pudrición blanca de la madera presentan diversidad genética para la degradación de contaminantes ambientales; sin embargo, desde el punto de vista práctico, el mecanismo ligninolítico es el más eficiente por ser extracelular, altamente inespecífico y fácilmente inducible. De otro lado, la presencia de mecanismos intracelulares de degradación de contaminantes vía reductiva y oxidativa les pueden permitir tolerar mayores concentraciones y por tanto hacerlos más aptos para procesos de biorremediación en donde los procesos oxidativos extracelulares con las enzimas ligninolíticas sean la característica usada para alcanzar la descontaminación.
Biodegradación de plaguicidas
Pentaclorofenol (PCP). El pentaclorofenol (PCP) fue producido inicialmente como una sustancia para la conservación de la madera. Posteriormente también se ha usado como insecticida contra una amplia variedad de plagas. Debido a su toxicidad la EPA-USA lo definió como contaminante prioritario y actualmente está restringido su uso. La biodegradación de PCP por hongos de la pudrición blanca, se ha estudiado principalmente con P. chrysosporium y P. sordida tanto en medio líquido como en suelo, obteniéndose entre el 78 y 90% de degradación (Mileski et al., 1988; Lamar y Dietrich, 1990; Udayasoorian et al., 2007). También se han realizado estudios con Trametres versicolor (Tuomela et al., 1998; Walker et al., 2005), Pleurotus ostreatus, Bjerkandera adusta, Irpex lacteus y Antracophyllum discolor (Ruttimann y Lamar, 1997; Cea et al., 2010) (Tabla 2). El PCP es uno de los pocos plaguicidas en los que se ha demostrado que su degradación inicial (declorinación parcial) por hongos de pudrición blanca, se debe al mecanismo oxidativo ligninolítico. En su declorinación total intervienen algunas etapas de deshalogenación reductiva de la fase II del metabolismo (Reddy y Gold, 2000). En la Figura 3, se observa la vía metabólica propuesta para la declorinación total de PCP con P. chrysosporium.
Tabla 2. Capacidad de degradación y mineralización de plaguicidas por hongos de la pudrición blanca de la madera.
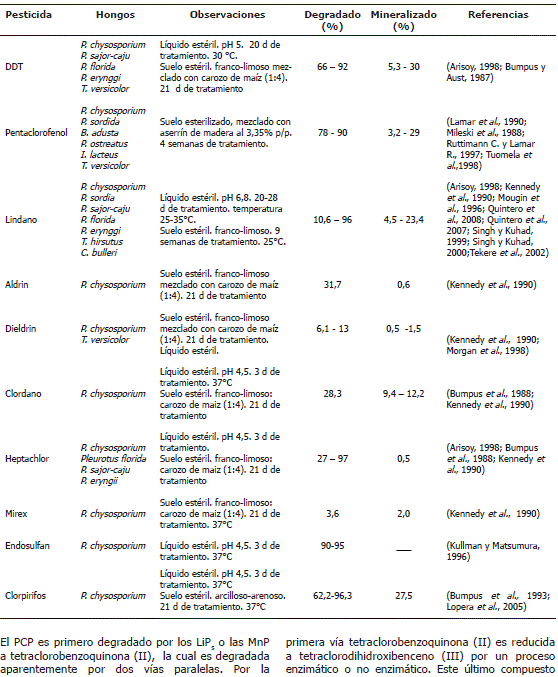
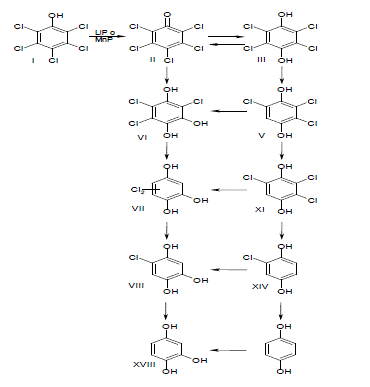
Figura 3. Propuesta de vía de biodegradación de PCP por Phanerochae chrysosporium (Reddy y Gold, M.H., 2000).
El PCP es primero degradado por los LiPs o las MnP a tetraclorobenzoquinona (II), la cual es degradada aparentemente por dos vías paralelas. Por la primera vía tetraclorobenzoquinona (II) es reducida a tetraclorodihidroxibenceno (III) por un proceso enzimático o no enzimático. Este último compuesto sufre cuatro declorinaciones sucesivas pasando por los compuestos (V, XI y XIV) hasta 1,4 dihidroxibenceno que es hidroxilado para formar trihidroxibenceno (XVII). Por otro lado, la tetraclorobenzoquinona (II) es convertida a triclorotrihidroxibenceno (VI) que posteriormente sufre tres declorinaciones sucesivas hasta formar trihidroxibenceno (XVII) (Reddy y Gold, 2000).
Lindano. El lindano es un insecticida cuya formulación inicial contenía una mezcla de isómeros del hexaclorociclohexano (α-, β-, γ- y δ-HCH), de los cuales el lindano es el isómero γ-HCH. Este compuesto ha sido utilizado en agricultura y en salud humana y animal porque es un insecticida de amplio espectro. Su toxicidad ha sido comprobada, y ha sido prohibido en más de 52 países. Varios trabajos han mostrado que es degradado y mineralizado por muchos hongos de pudrición blanca de la madera (Tabla 2). En cultivos en suelo, P. chrysosporium ha mineralizado lindano hasta en un 22,3% inoculando el hongo inmovilizado sobre carozo de maíz, en una relación 4:1 g de sustrato: g de suelo (Kennedy et al., 1990), mientras que con Bjerkandera adusta se ha alcanzado hasta un 17% de degradación inoculado con el mismo sistema de tratamiento (Quintero et al., 2008). P. chrysosporium no ha mostrado capacidad de degradación ni mineralización cuando se inocula sobre suelo de forma no inmovilizada (Mougin et al., 1997; Quintero et al., 2008). Algunos autores han indicado que la degradación de lindano se incrementa con la presencia de hongos sobre suelos contaminados no estériles alcanzándose una relación de sinergismo con la microflora indígena del suelo, a una concentración de 2 mg de biomasa fúngica/g de suelo (Mougin et al., 1997), sin embargo, otros trabajos muestran que la acción de los hongos se ve inhibida a concentraciones de 0,6 mg de biomasa fúngica/g de suelo (Quintero et al. 2008). Es posible que esta diferencia se deba a las diferentes relaciones de inóculo en cada tratamiento. Lo anterior permite señalar la necesidad de optimizar la masa de inóculo fúngico e incluir un sustrato selectivo que les permita mantenerse y proliferar adecuadamente. Estas condiciones se cumplen con la inoculación de los hongos inmovilizados sobre residuos lignocelulósicos; sin embargo, aún con esta estrategia de inoculación, los porcentajes de degradación son relativamente modestos. Por esto se han evaluado sistemas de tratamiento de suelo en fase suspensión “slurry” con B. adusta alcanzando degradaciones de lindano hasta del 94% (Quintero et al., 2007), que son altamente superiores a los alcanzados sobre suelo en fase sólida, lo que muestra que en sistemas en fase sólida existen importantes limitaciones a la transferencia de masa (Rijnaarts et al., 1990).
Por otro lado, se ha comprobado que las enzimas ligninolíticas no son responsables de la degradación del lindano. Además, se han encontrado algunos metabolitos intermediarios de la degradación fúngica como tetraclorocliclohexeno (TCCH), tetraclorociclohexeno-epóxido (TCCH-epóxido) y tetraclorociclohexenol (TCCOL), que no son característicos de ataques oxidativos (Mougin et al., 1996), estos resultados han sido corroborados por nuevas investigaciones (Singh y Kuhad, 1999; 2000).
Con base en la reducción de la degradación de lindano encontrada al aplicar algunos inhibidores de las enzimas P-450s, se planteó que estas enzimas podrían estar involucradas en su degradación (Mougin et al., 1996). También se ha planteado otra hipótesis en la cual las etapas iniciales en la degradación del lindano corresponden a una deshalogenación reductiva de la fase II del metabolismo (Singh y Kuhad, 1999). Hasta el presente solo se ha planteado una propuesta de las etapas iniciales de biodegradación del lindano con hongos de la pudrición blanca de la madera (Figura 4) (Singh y Kuhad, 2000). Es posible que durante la transformación del TCCH en TCCOL se forme el TCCH epóxido como un intermediario, puestos que también ha sido detectado en algunos trabajos ya descritos.
Figura 4. Etapas de reacción de la degradación de lindano por hongos de la pudrición blanca (Singh y Kuhad, 2000).
DDT. El insecticida 1,1,1-tricloro-2,2-bis(p-clorofenil)etano (DDT) es un contaminante ambiental persistente, degradado y mineralizado por cultivos de diferentes hongos de la pudrición blanca (Tabla 2). En algunos trabajos se indica que la biodegradación ha alcanzado valores entre 30 y 90%, mientras que su grado de mineralización ha sido reportado entre 5,3 y 30%, en cultivos líquidos y del 4% en tratamientos sobre suelo contaminado (Zhao y Yi, 2010; Siripong et al., 2009, Arisoy, 1998; Bumpus y Aust, 1987). Varios trabajos han concluido que la mineralización del DDT depende de la acción de las enzimas ligninolíticas (Bumpus et al., 1985; Bumpus y Aust, 1987), pero en contraste con la mineralización, la biodegradación no depende de ellas (Kohler et al., 1988). El DDT es deshalogenado reductivamente a 1,1-dicloro-2,2-bis(4-clorofenil) etano (DDD) o hidroxilado a 2,2,2-tricloro-1,1-bis (4-clorofenil)etanol (Dicofol) (Hammel, 1992). El 1,1-dicloro-2,2-bis(4-clorofenil)etileno (DDE) se ha observado en cultivos autoclavados lo que indica que se puede formar de manera abiótica (Fernando et al., 1989). Estos metabolitos intermediarios se forman y después desaparecen lo cual indica que estos productos tienden a la mineralización (Figura. 5) (Field, 2004).
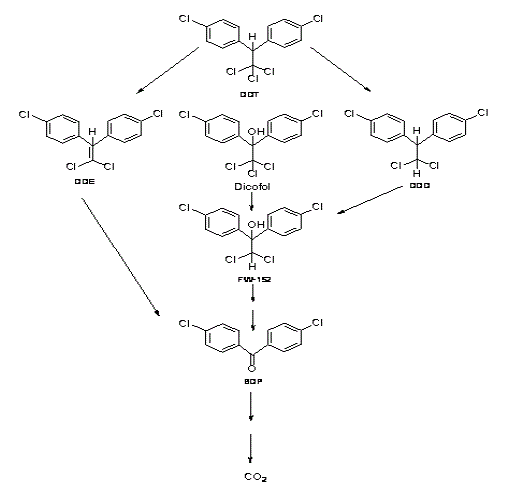
Figura 5. Etapas iniciales en la biodegradación de DDT por hongos de la pudrición blanca (Field, 2004).
Aldrín, Dieldrín, Mirex, Heptaclor, Clordano y Endosulfán. Estos insecticidas del grupo de los ciclodienos han sido extensamente empleados debido a su eficiencia en el control de plagas agrícolas en cultivos de café, algodón, cereales y para el control de vectores de enfermedades como la malaria. A pesar de su elevada capacidad insecticida, su uso se ha minimizado o eliminado debido a su persistencia en la naturaleza, bioacumulación y toxicidad en animales superiores, exceptuando al Endosulfán, los demás pertenecen a la llamada “docena sucia”.
Existen evidencias de la capacidad de los hongos de pudrición blanca de la madera para realizar la biodegradación de estos compuestos y se han hecho estudios tanto en matrices acuosas como en suelo (Tabla 2); sin embargo, solo se ha observado una mineralización significativa de clordano. La bioconversión de Aldrín por P. chrysosporium produce Dieldrín, como resultado de una reacción de epoxidación en la cual no hay declorinación (Kennedy et al., 1990). Para Heptaclor se han encontrado porcentajes de biodegradación entre 71 y 97% con diferentes hongos de la pudrición blanca, siendo las reacciones de hidrólisis e hidroxilación las etapas de la vía de degradación hasta ahora dilucidadas (Xiao et al., 2011; Arisoy, 1998).
El insecticida Endosulfán es altamente degradado por P. chrysosporium; sin embargo, en las etapas iniciales de la degradación, no están involucradas las enzimas ligninolíticas y se ha propuesto que las enzimas P-450s son las responsables de la degradación inicial de este insecticida (Kullman y Matsumura, 1996). Se ha demostrado que este hongo tiene la capacidad de oxidar y también hidrolizar directamente este pesticida, cada una de estas dos vías se activa dependiendo si el metabolismo tiene lugar bajo condiciones ligninolíticas o no ligninolíticas. Estos productos de transformación incluyen endosulfán sulfato, endosulfán diol, endosulfán hidroxieter y endosulfán dialdehido. Kullman y Matsumura (1996) han propuesto una vía de biodegradación con P. chrysosporium (Figura 6).
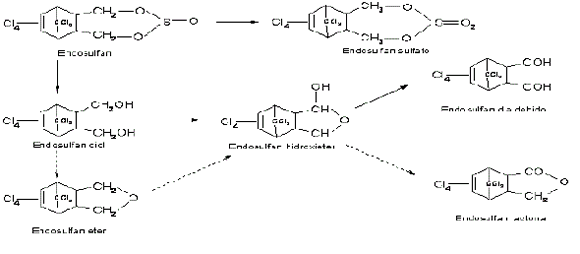
Figura 6. Vía metabólica propuesta para la biodegradación del endosulfán por Phanerochaete chrysosporium. Las líneas sólidas indican el metabolismo principal, las líneas punteadas son vías metabólicas menos probables (Kullman y Matsumura, 1996).
Clorpirifos. Estos compuestos organofosforados poseen actividad insecticida de amplio rango, son ampliamente usados en la agricultura pero también son altamente tóxicos para los mamíferos. Con este insecticida se han realizado pocos trabajos para estudiar su biodegradadación. En un estudio de biodegradación con P. chrysosporium, la mineralización alcanzó el 27,5% (Tabla 2) y se observó la ruptura del anillo de pirimidina (Bumpus et al., 1993). Clorpirifos inicialmente es hidrolizado para producir 3,5,6 tricloropiimidol (TCP) (Figura 7). Los hongos Hypholoma fasciculare y Coriolus versicolor han mostrado capacidad para degradar clorpirifos en un 29% y 36% en 42 d respectivamente, durante tratamientos de suelos mezclados con paja de trigo. En este trabajo no se observó relación entre la degradación y la actividad ligninolítica (Bending et al., 2002). Otro trabajo llevado a cabo con P. chysosporium mostró altas tasas de degradación de Clorpirifos del 96%, 82% y 62% en 3 semanas de tratamiento sobre suelo mezclado con carozo de maíz a concentraciones de 1, 5 y 9 mg/kg respectivamente (Lopera et al., 2005). Estudios con P. chysosporium, Trametes versicolor y Pleurotus osreatus, han mostrado que las enzimas ligninolíticas no están asociadas con la degradación de clorpirifos (Karas et al., 2011), sin embargo, en ninguno de estos trabajos se discute sobre los mecanismos empleados para su degradación. Esos resultados coinciden con observaciones hechas sobre la degradación de otros organofosforados como parathion y tribufos con este tipo de hongos, que indican su origen intracelular y que las enzimas ligninolíticas no juegan un papel importante en este proceso (Jauregui et al., 2003).
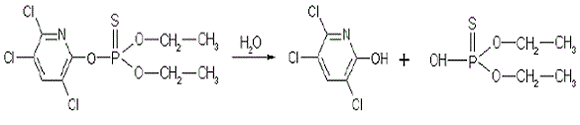
Figura 7. Primera etapa de la vía metabólica de degradación de Clorpirifos por hongos de la pudrición blanca de la madera (Bumpus et al. 1993).
La degradación de plaguicidas por hongos ligninolíticos muestra que el mecanismo ligninolítico es el menos empleado y los procesos reductivos, seguido de la oxidación intracelular por las citocromo P-450 monooxigenasas son los mecanismos involucrados en las biotransformaciones de los plaguicidas. Esto se explica por el elevado potencial de ionización que tienen los plaguicidas como por ejemplo el lindano con 11,4 eV, DDT con 9,6 eV, entre otros (Tanabe y Matsumoto, 2002), lo que también ha sido sugerido por otros autores (Dávila et al., 2005). Se requiere orientar esfuerzos hacia el estudio de los mecanismos de control de los sistemas degradativos intracelulares con el fin de incrementar su actividad y ampliar el espectro de aplicación de los hongos de la pudrición blanca de la madera en biorremediación. Se observa que este tipo de hongos muestra gran diversidad enzimática que los mantendrán como una herramienta de gran valor en biotecnología para la recuperación de ambientes contaminados.
CONCLUSIONES
La mayoría de los tratamientos de biorremediación se han realizado con bacterias, debido a que son fácilmente cultivables, crecen rápidamente y son capaces de usar los contaminantes orgánicos como fuentes de carbono y energía. Sin embargo, en los últimos años, el empleo de los hongos de la pudrición blanca, ha demostrado el gran potencial que poseen para la biodegradación de un amplio espectro de xenobióticos como los insecticidas químicos. Estos hongos exhiben tres principales sistemas enzimáticos empleados para la degradación de contaminantes ambientales. Siendo el mecanismo ligninolítico el más ampliamente estudiado y usado en biorremediación, pues se ha demostrado que este sistema de degradación de la lignina, interviene en la oxidación de un número importante de xenobióticos como hidrocarburos aromáticos policíclicos, tintes sintéticos y naturales y algunos plaguicidas como el pentaclorofenol. Sin embargo, su capacidad oxidativa se limita a compuestos cuyos potenciales de oxidación (PI) sean inferiores a 8,0 eV.
Por el contrario, muchos plaguicidas presentan valores significativamente superiores de PI, indicando que el mecanismo ligninolítico no es viable para degradar estos compuestos. De acuerdo con un gran número de trabajos, los mecanismos de degradación de plaguicidas organoclorados y organofosforados son intracelulares principalmente de origen reductivo y en menor proporción oxidativo. La explotación de los mecanismos del citocromo p-450 y de las enzimas dependientes del glutatión, el conocimiento de sus mecanismos de expresión a nivel molecular, su caracterización cinética en ambientes similares a los encontrados en suelos y aguas contaminadas, permitirá disponer de una nueva herramienta que brindan estos maravillosos hongos de la madera, para reducir los efectos antropogénicos de las diferentes actividades agrícolas, industriales y urbanas.
BIBLIOGRAFÍA
Arisoy, M. 1998. Biodegradation of chlorinated organic compounds by white-rot fungi. B. Bulletin of Environmental Contamination and Toxicology 60(6): 872-876. [ Links ]
Atlas, R.M. and R. Unterman. 1998. Bioremediation. pp. 666-681. In: Demain, A.L. and J.E. Davies (ed.). Manual of Industrial Microbiology and Biotechnology. Second edition. SM Press, Washington D.C. [ Links ]
Aust, D. and J. Benson. 1993. The fungus among us: Use of white rot fungi to biodegrade environmental pollutants. Environmental Health Perspectives 101(3): 232-233. [ Links ]
Barr, D.P. and S.D. Aust. 1994. Mechanisms white rot fungi use to degrade pollutants. Environmental Science and Technology 28(2): 78A-87A. [ Links ]
Bending, G.D., M. Friloux and A. Walker. 2002. Degradation of contrasting pesticides by white rot fungi and its relationship with ligninolytic potential. FEMS Microbiology Letters 212(1): 59-63. [ Links ]
Bezalel, L., Y. Hadar and C. Cerniglia. 1997. Enzymatic mechanisms involved in phenanthrene degradation by the white rot fungus Pleurotus ostreatus. Applied and Environmental Microbiology 63(7): 2495-2501. [ Links ]
Bogan, B.W. and R.T. Lamar. 1996. Polycyclic aromatic hydrocarbon-degrading capabilities of Phanerochaete laevis HHB-1625 and its extracellular ligninolytic enzymes. Applied and Environmental Microbiology 62(5): 1597-1603. [ Links ]
Bogan, B.W. and R.T. Lamar. 1995. One-electron oxidation in the degradation of creosote polycyclic aromatic hydrocarbons by Phanerochaete chrysosporium. Applied and Environmental Microbiology 61(7): 2631-2635. [ Links ]
Bucke, C. 1998. Biochemistry of bioremediation of fungi. Journal of Chemical Technology and Biotechnology 71(4): 356-357. [ Links ]
Bumpus, J.A., Kakar S.N. and R.D. Coleman. 1993. Fungal degradation of organophosphorus insecticides. Applied Biochemistry and Biotechnology 39-40: 715-726. [ Links ]
Bumpus, J.A. and D. Aust. 1987. Biodegradation of DDT [1,1,1-trichloro-2,2-bis4-chlorophenylethane] by the white rot fungus Phanerochaete chrysosporium. Applied and Environmental Microbiology 53(9): 2001-2008. [ Links ]
Bumpus, J.A., M. Tien, D. Wright and S.D. Aust. 1985. Oxidation of persistent environmental pollutants by a white rot fungus. Science 228(4706): 1434-1436. [ Links ]
Bumpus, J.A., F. Tudor, M.A. Jurek and S.D. Aust. 1988. Factors affecting biodegradation of xenobiotics by the white rot fungus Panerochaete chrysosporium. pp. 363-381. In: Lewandowski, G. (ed.). Biotechnology applications in hazardous waste treatment. Engineering Foundation, New York, USA. 423 p. [ Links ]
Cameron, M.D., S. Timofeevski and S.D. Aust. 2000. Enzymology of Phanerochaete chrysosporium with respect to the degradation of recalcitrant compounds and xenobiotics. Applied Microbiology and Biotechnology 54(6): 751-758. [ Links ]
Caplan, J.A. 1993. The worldwide bioremediation industry: prospects for profit. Trends Biotechnology 11(8): 320-323. [ Links ]
Casillas, R.P., S.A. Crow, T.M. Heinze, J. Deck and C.E. Cerniglia 1996. Initial oxidative and subsequent conjugative metabolites produced during the metabolism of phenanthrene by fungi. Journal of Industrial Microbiology and Biotechnology 16(4): 205-215. [ Links ]
Cea, M., M. Jorquera, O. Rubilar, H. Langer, G. Tortella and M.C. Dieza. 2010. Bioremediation of soil contaminated with pentachlorophenol by Anthracophyllum discolor and its effect on soil microbial community. Journal of Hazardous Materials 181(1-3): 315–323.
Cerniglia C.E., J.P. Freeman and R.K. Mitchum. 1982. Glucuronide and sulfate conjugation in the fungal metabolism of aromatic hydrocarbons. Applied and Environmental Microbiology 43(5): 1070-1075. [ Links ]
Christian V., R. Shrivastava, D. Shukla, H.A. Modi and B.R. Vyas. 2005. Degradation of xenobiotic compounds by lignin-degrading white-rot fungi: Enzymology and mechanisms involved. Indian Journal of Experimental Biology 43(4): 301-312. [ Links ]
Collins P.J., M.J. Kotterman, J.A. Field and A.D. Dobson. 1996. Oxidation of anthracene and benzo[a]pyrene by laccases from trametes versicolor. Applied and Environmental Microbiology 62(12): 4563-4567. [ Links ]
Davila, G., R. Tinoco, M. Pickard and R. Vazquez. 2005. Transformation of halogenated pesticides by versatile peroxidase from Bjerkandera adusta. Enzyme and Microbial Technology 36(2-3): 223-231. [ Links ]
Dowd, A.C., C.M. Buckley. and D. Sheehan. 1997. Glutathione S-transferases from the white-rot fungus, Phanerochaete chrysosporium. Biochemical Journal 324: 243-248. [ Links ]
Dua, M., A. Singh, N. Sethunathan and A.K. Johri. 2002. Biotechnology and bioremediation: successes and limitations. Applied Microbiolology and Biotechnology 59(2-3): 143-152. [ Links ]
Eilers, A., E. Rüngeling, U.M. Stündl and G. Gottschalk 1999. Metabolism of 2,4,6-trinitrotoluene by the white-rot fungus Bjerkandera adusta DSM 3375 depends on cytochrome P-450. Applied Microbiology and Biotechnology 53(1): 75-80. [ Links ]
Fernando, T., Aust S.D. and J.A. Bumpus. 1989. Effects of culture parameters on DDT 1,1,1-trichloro-2,2-bis4-chlorophenylethane biodegradation by Phanerochaete chrysosporium. Chemosphere 19(8-9): 1387-1398. [ Links ]
Ferreira, V., D. Magalhaes, S. Kling, J. da Silva and E. Bon. 2000. N-demethylation of methylene blue by lignin peroxidase from Phanerochaete chrysosporium stoichiometric relation for H2O2 consumption. Applied Biochemistry and Biotechnology 84(86): 255-265. [ Links ]
Field, J.A., H. Baten, F. Boelsma and H. Wim. 1996. Biological elimination of polycyclic aromatic hydrocarbons in solvent extracts of polluted soil by the white rot fungus, Bjerkandera sp strain BOS55. Environmental Technology 17(3): 317-323. [ Links ]
Field, J.A. and E.M. Thurman. 1996. Glutathione conjugation and contaminant transformation. Environmental Science Technology 30: 1413-1418. [ Links ]
Field, J.A. 2004. Biodegradation of chlorinated compounds by white rot fungi. pp. 159-204. In: Häggblom M. and I.D. Bossert (eds.). Dehalogenation: Microbial processes and environmental applications. Springer, New York. 481 p. [ Links ]
Furukawa, K. 2003. ‘Super bugs’ for bioremediation. Trends Biotechnology 21(5): 187-190.
Hammel, K.E. 1992. Oxidation of aromatic pollutants by lignin-degrading fungi and their extracellular peroxidases. pp. 41-60. In: H. Sigel and A. Sigel (eds.). Degradation of environmental pollutants by microorganisms and their metalloenzymes. Institute of Inorganic Chemistry, University of Basel, Switzerland. 557 p. [ Links ]
Hammel, K.E., B. Kalyanaraman and T.K. Kirk. 1986. Oxidation of polycyclic aromatic hydrocarbons and dibenzo[p]-dioxins by Phanerochaete chrysosporium ligninase. Journal of Biological Chemistry 261: 16948-16952. [ Links ]
Harshavardhan, D., C. Ranajit and Y. Jaqjit. 2005. Genome-wide structural and evolutionary analysis of the P450 monooxygenase genes P450ome in the white rot fungus Phanerochaete chrysosporium: evidence for gene duplications and extensive gene clustering. BMC Genomics 6: 1-24. [ Links ]
Hiratsuka, N., H. Wariishi and H. Tanaka. 2001. Degradation of diphenyl ether herbicides by the lignin-degrading basidiomycete Coriolus versicolor. Applied Microbiology and Biotechnology 57(4): 563-571. [ Links ]
Hofrichter, M. 2002. Review: lignin conversion by manganese peroxidase MnP. Enzyme and Microbial Technology 30(4): 454-466. [ Links ]
Ichinose, H., H. Wariishi and H. Tanaka H. 2002a. Identification and characterization of novel cytochrome P450 genes from the white-rot basidiomycete, Coriolus versicolor. Applied Microbiology and Biotechnology 58(1): 97-105. [ Links ]
Ichinose, H., H. Wariishi and H. Tanaka. 2002b. Identification and heterologous expression of the cytochrome P450 oxidoreductase from the white-rot basidiomycete Coriolus versicolor. Applied Microbiology and Biotechnology 59(6): 658-664. [ Links ]
Jauregui, J., B. Valderrama, A. Albores and R. Vazquez. 2003. Microsomal transformation of organophosphorus pesticides by white rot fungi. Biodegradation 14(6): 397-406. [ Links ]
Jerina D.M. 1983. Metabolism of aromatic hydrocarbons by the cytochrome P-450 system and epoxide hydrolase. Drug Metabolism and Disposition 11: 1-4. [ Links ]
Karas P., C. Perruchon, K. Exarhou, C. Ehaliotis and D. Karpouzas. 2011. Potential for bioremediation of agro-industrial effluents with high loads of pesticides by selected fungi. Biodegradation 22(1):215-28. [ Links ]
Kearney P.C. and T. Roberts. 1998. Pesticide Remediation in soil and water. John Wiley and Sons, Baffins Lane, Chichester, England. 382 p. [ Links ]
Kennedy, D., S. Aust and A. Bumpus. 1990. Comparative biodegradation of alkyl halide insecticides by the white rot fungus, Phanerochaete chrysosporium BKM-F-1767. Applied Environmental Microbiology 56(8): 2347-2353. [ Links ]
Khindaria, A., D.P. Barr and S.D. Aust. 1995. Lignin peroxidases can also oxidize manganese. Biochemistry 34(23): 7773-7779. [ Links ]
Kirk, T.K. and R. L. Farrell. 1987. Enzymatic “combustion”: The microbial degradation of lignin. Annual Review of Microbiology 41: 465-505.
Kohler, A., A. Jager, H. Willerhausen and H. Graf. 1988. Extracellular ligninase of Phanerochaete chrysosporium Burdsall has no role in the degradation of DDT. Applied Microbiology and Biotechnology 29(6): 618-620. [ Links ]
Kullman, S.W. and Matsumura F. 1997. Identification of a novel cytochrome P-450 gene from the white-rot fungus Phanerochaete chrysosporium. Applied and Environmental Microbiology 63: 2741-2746. [ Links ]
Kullman, S.W. and F. Matsumura. 1996. Metabolic pathways utilized by Phanerochaete chrysosporium for degradation of the cyclodiene pesticide endosulfan. Applied and Environmental Microbiology 62(2): 593-600. [ Links ]
LaGrega, M.D., P.L. Buckingham and J.C. Evans. 1996. Gestión de residuos tóxicos. Tratamiento, eliminación y recuperación de suelos. McGraw-Hill, Madrid. 1316 p. [ Links ]
Lamar, R.T. and D.M. Dietrich. 1990. In situ depletion of pentachlorophenol from contaminated soil by Phanerochaete spp. Applied and Environmental Microbiology 56(10): 3093-3100. [ Links ]
Lamar, R.T., Larsen M.J. and Kirk T.K. 1990. Sensitivity to and degradation of pentachlorophenol PCP by Phanerochaete chrysosporium. Applied and Environmental Microbiology 56(11): 3519-3526. [ Links ]
Lewis, D.F. 2001. COMPACT: a structural approach to the modelling of cytochromes P450 and their interactions with xenobiotics. Journal of Chemical Technology and Biotechnology. 76(3): 237-244. [ Links ]
Li, Y.F. 1999. Global technical hexachlorocyclohexane usage and its contamination consequences in the environment: from 1948 to 1997. The Science of the Total Environment 232(3): 121-158. [ Links ]
Liu, S. and J.M. Suflita. 1993. Ecology and evolution of microbial populations for bioremediation. Trends Biotechnology 11(8): 344-352. [ Links ]
Lopera, M.M., G. Peñuela, M. Damínguez y G. Mejía. 2005. Evaluación de la degradación del plaguicida clorpirifos en muestras de suelo utilizando el hongo Phanerochaete chrysosporium. Revista Facultad de Ingeniería, Universidad de Antioquia 33: 58-69. [ Links ]
Masaphy, S., Henis Y. and D. Levanon. 1996a. Manganese-enhanced biotransformation of atrazine by the white rot fungus Pleurotus pulmonarius and its correlation with oxidation activity. Applied and Environmental Microbiology 62(10): 3587-3593. [ Links ]
Masaphy, S., D. Levanon, Y. Henis, K. Venkateswarlu and S. Kelly. 1996b. Evidence for cytochrome P-450 and P-450-mediated benzo(a)pyrene hydroxylation in the white rot fungus Phanerochaete chrysosporium. FEMS Microbiology Letters 135(1): 51-55. [ Links ]
Mileski, G.J., J.A. Bumpus, M.A. Jurek and S. Aust. 1988. Biodegradation of pentachlorophenol by the white rot fungus Phanerochaete chrysosporium. Applied and Environmental Microbiology 54(12): 2885-2889. [ Links ]
Moreira, M.T., G. Feijoo and J.M. Lema. 2000. Manganese peroxidase production by Bjerkandera sp. BOS55. 1. Regulation of enzymatic production. Bioprocess and Biosystems Engineering 23(6): 657-661. [ Links ]
Moreira, M.T., C. Palma, I. Mielgo, G. Feijoo and J.M. Lema. 2001. In vitro degradation of a polymeric dye Poly R-478 by manganese peroxidase. Biotechnology and Bioengineering 75(3): 362-368. [ Links ]
Morgan, P., T. Stephen and R.J. Watkinson. 1991. Comparison of abilities of white-rot fungi to mineralize selected xenobiotic compounds. Applied Microbiology and Biotechnology 34(5): 693-696. [ Links ]
Mougin, C., C. Pericaud, J. Dubroca and M. Asther. 1997. Enhanced mineralization of lindane in soils supplemented with the white rot basidiomycete Phanerochaete chrysosporium. Soil Biology and Biochemistry 29(9-10): 1321-1324. [ Links ]
Mougin, C., C. Pericaud, C. Malosse, C. Laugero and M. Asther. 1996. Biotransformation of the insecticide lindane by the white rot basidiomycete Phanerochaete chrysosporium. Pest Management Science 47(1): 51-59. [ Links ]
Nyholm, N., B.N. Jacobsen, B.M. Pedersen, O. Poulsen, A. Damborg and B. Schultz. 1992. Removal of organic micropollutants at Ppb levels in laboratory activated sludge reactors under various operating-conditions biodegradation. Water Research 26(3): 339-353. [ Links ]
Paoletti, M.G. and D. Pimentel. 2000. Environmental risks of pesticides versus genetic engineering for agricultural pest control. Journal of Agricultural and Environmental Ethics 12(3): 279-303. [ Links ]
Quintero, J.C., G. Feijoo and J.M. Lema. 2006. Producción de enzimas ligninolíticas con hongos basidiomicetos cultivados sobre materiales lignocelulósicos. Vitae 13: 61-67. [ Links ]
Quintero, J.C., T.L. Lú-Chau, M.T. Moreira, G. Feijoo and J.M. Lema. 2007. Bioremediation of HCH present in soil by the white-rot fungus Bjerkandera adusta in a slurry batch bioreactor. International Biodeterioration and Biodegradation 60(4): 319-326. [ Links ]
Quintero, J.C., M.T. Moreira, G. Feijoo and J.M. Lema. 2008. Screening of white rot fungal species for their capacity to degrade lindane and other isomers of hexachlorocyclohexane HCH. Ciencia e Investigación Agraria 35(2): 159-167. [ Links ]
Reddy, G.V. and M.H. Gold. 1999. A two-component tetrachlorohydroquinone reductive dehalogenase system from the lignin-degrading basidiomycete Phanerochaete chrysosporium. Biochemical and Biophysical Research Communications 257(3): 901-905. [ Links ]
Reddy, G.V. and M.H. Gold. 2001. Purification and characterization of glutathione conjugate reductase: a component of the tetrachlorohydroquinone reductive dehalogenase system from Phanerochaete chrysosporium. Archives of Biochemistry and Biophysics 391(2): 271-277. [ Links ]
Reddy, G.V., M.D. Gelpke and M.H. Gold. 1998. Degradation of 2,4,6-trichlorophenol by Phanerochaete chrysosporium: Involvement of reductive dechlorination. Journal of Bacteriology 180(19): 5159-5164. [ Links ]
Reddy, G.V. and M.H. Gold. 2000. Degradation of pentachlorophenol by Phanerochaete chrysosporium: intermediates and reactions involved. Microbiology 146: 405-413. [ Links ]
Riahi, S., P. Norouzi, M. Bayandori, M. Ganjali. G. Karimipour and H. Sharghi. 2007. Theoretical and experimental report on the determination of oxidation potentials of dihydroxyanthracene and thioxanthens derivatives. Chemical Physics 337(1-3): 33-38. [ Links ]
Rijnaarts, H., A. Bachmann, J. Jumelet and A- Zehnder. 1990. Effect of desorption and intraparticule mass transfer on the aerobic biomineralization of alfa-hexachlorocyclohexane in a contaminated calcareous soil. Environmental Science and Technology 24(9): 1349-54. [ Links ]
Ruttimann, C. and R. Lamar. 1997. Binding of substances in pentachlorophenol to humic soil by the action of white rot fungi. Soil Biology and Biochemistry 29(7): 1143-1148. [ Links ]
Sack, U., M. Hofrichter and W. Fritsche. 1997. Degradation of polycyclic aromatic hydrocarbons by manganese peroxidase of Nematoloma frowardii. FEMS Microbiology Letters 152(2): 227-234. [ Links ]
Sariaslani, F.S. 1991. Microbial cytochromes P-450 and xenobiotic metabolism. Advances in Applied Microbiology 36: 133-178. [ Links ]
Semple, K.T., A. Morriss and G. Paton. 2003. Bioavailability of hydrophobic organic contaminants in soils: fundamental concepts and techniques for analysis. European Journal of Soil Science 54(4): 809-818. [ Links ]
Singh, B.K. and R.C. Kuhad. 1999. Biodegradation of lindane gamma-hexachlorocyclohexane by the white-rot fungus Trametes hirsutus. Letters in Applied Microbiology 28(3): 238-241. [ Links ]
Singh, B.K. and R.C. Kuhad. 2000. Degradation of insecticide lindane gamma-HCH by white-rot fungi Cyathus bulleri and Phanerochaete sordida. Pest Management Science 56(2): 142-146. [ Links ]
Siripong, P., B. Oraphin, T. Sanro and P. Duanporn. 2009. Screening of fungi from natural sources in Thailand for degradation of polychlorinated hydrocarbons. American Eurasian Journal Agricultural and Environmental Science 5(4): 466-472. [ Links ]
Summersgill, I.M. and D.W. Scott. 2005. Remediation technology costs in the UK and Europe. Report. VHE Technology. Barnsley, UK. 8 p. [ Links ]
Sutherland, J.B. 1992. Detoxification of polycyclic aromatic hydrocarbons by fungi. Journal of Industrial Microbiology and Biotechnology 9(1): 53-61. [ Links ]
Tanabe, K. and T. Matsumoto. 2002. Prediction of carcinogenicity of chlorine-containing organic compounds by neuronal network. Journal of Computer Chemistry, Japan 1(1): 23-28. [ Links ]
Tekere, M., I. Ncube, J.S. Read and R. Zvauya. 2002. Biodegradation of the organochlorine pesticide, lindane by a sub-tropical white rot fungus in batch and packed bed bioreactor systems. Environmental Technology 23(2): 199-206. [ Links ]
Tuomela, M., M. Lyytikainen, P. Oivanen and A. Hatakka. 1998. Mineralization and conversion of pentachlorophenol PCP in soil inoculated with the white-rot fungus Trametes versicolor. Soil Biology and Biochemistry 31(1): 65-74. [ Links ]
Udayasoorian, C., P.C. Prabu and G. Balasubramanian. 2007. Degradation of pentachlorophenol by white rot fungus (Phanerochaete chrysosporium-TL 1) grown in ammonium lignosulphonate media. Biotechnology 6(1): 76-80. [ Links ]
Ullah, M.A., H. Kadhim, R.A. Rastall and C.S. Evans. 2000. Evaluation of solid substrates for enzyme production by Coriolus versicolor, for use in bioremediation of chlorophenols in aqueous effluents. Applied Microbiology and Biotechnology 54(6): 832-837. [ Links ]
Urlacher, V.B. and S. Eiben. 2006. Cytochrome P450 monooxygenases: perspectives for synthetic application. Trends in Biotechnology 24(7): 324-330. [ Links ]
Valli, K., H. Wariisshi and M.H. Gold. 1992. Degradation of 2,7-dichlorodibenzo-p-dioxin by the lignin-degrading basidiomycete Phanerochaete chrysosporium. Journal of Bacteriology 174(7): 2131-2137. [ Links ]
Vázquez, D.R., D.W. Westlake and P.M. Fedorak. 1994. Lignin peroxidase oxidation of aromatic compounds in systems containing organic solvents. Applied and Environmental Microbiology 60(2): 459-466. [ Links ]
Walter, M., K. Boyd, L. Boul, C. Ford, D. McFadden, B. Chong and J. Pinfold. 2005. Field-scale bioremediation of pentachlorophenol by Trametes versicolor. International Biodeterioration and Biodegradation 56(1): 51-57. [ Links ]
Wong, M., A. Leung, J. Chan and M. Choi. 2005. A review on the usage of POP pesticides in China, with emphasis on DDT loadings in human milk. Chemosphere 60(6): 740-752. [ Links ]
Wood, P.M. 1992. A comparation of peroxidase and cytochrome P-450. Biochemical Society Transactions 20(2): 349-352. [ Links ]
Xiao, P., T. Mori, I. Kamei and R. Kondo. 2011. Metabolismoforganochlorine pesticide heptachlorand its metabolite heptachlor epoxide by white rot fungi, belonging to genus Phlebia. FEMS Microbiology Letters 314(2): 140–146.
Zhao, Y. and X. Yi 2010. Effects of soil oxygen conditions and soil pH on remediation of DDT-contaminated soil by Laccase from white rot fungi. International Journal Environmental Research and Public Health 7(4): 1612-1621. [ Links ]

