










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.64 no.1 Medellín Jan./June 2011
COLECCIÓN Y CARACTERIZACIÓN MORFOAGRONÓMICA DEL SUBGÉNERO TACSONIA EN LA ZONA ANDINA DEL DEPARTAMENTO DE NARIÑO, COLOMBIA
COLLECTION AND MORPHOAGRONOMIC CHARACTERIZATION OF TACSONIA SUBGENDER IN THE ANDEAN ZONE OF THE DEPARTMENT OF NARIÑO, COLOMBIA
Óscar Checa Coral1; Edwin Rosero Álvarez2 e Iván Eraso Cultid3
1 Profesor Asociado. Universidad de Nariño. Facultad de Ciencias Agrícolas. Panamericana - Vipri, San Juan de Pasto, Colombia. <cicagrarias@hotmail.com>
2 Grupo de Investigación Cultivos Andinos. Universidad de Nariño. Facultad de Ciencias Agrícolas. Panamericana- Vipri, San Juan de Pasto, Colombia. <cicagrarias@hotmail.com>
3 Grupo de Investigación Cultivos Andinos. Universidad de Nariño. Facultad de Ciencias Agrícolas. Panamericana- Vipri, San Juan de Pasto, Colombia. <cicagrarias@hotmail.com>
Recibido: Marzo 3 de 2010; aceptado: Abril 18 de 2011
Resumen. En la zona andina de Nariño - Colombia, se colectaron 99 materiales, 91 del subgénero Tacsonia y 8 del subgénero Manicata. Las semillas se sembraron en una colección de trabajo, con el fin de preservarlas y realizarles la caracterización morfoagronómica “in vivo". Las muestras corresponden a las especies: Passiflora tarminiana Coppens y Barney, Passiflora tripartita var. mollissima (Kunt) Bailey, Passiflora tripartita (Juss) Poir, Passiflora mixta L. F., Passiflora cumbalensis (Karst) Harás, Passiflora pinnatistipula Cav, Passiflora x rosea Karst., Passiflora manicata (Juss) Pers y las accesiones BGP 63 y BGP 76. El 64,8% de los genotipos se ubican en la zona sur, destacándose el municipio de Aldana con mayor diversidad. El análisis de componentes principales (ACP) y el análisis de correspondencia múltiple (ACM), permitieron caracterizar las muestras y formar grupos mediante el método de caracterización jerárquica. En el ACP se conformaron 5 factores que explican el 66,30% de la variación total evaluada, la clasificación jerárquica permitió agrupar 4 clases, las características con mayor aporte a la variabilidad fueron: peso del fruto y longitud del pedúnculo, destacándose el grupo uno. El ACM expresó el 50,60% de la variabilidad total en los 5 primeros componentes, el análisis de clasificación permitió discriminar 5 grupos de especies, los caracteres que más aportaron a la variabilidad fueron: forma del fruto y presencia de antracnosis Colletotrichum gloeosporioides (Penzing) Penzing y Saccardo, los grupos de P. tarminiana y P. tripartita, se caracterizaron por ausencia y presencia de la enfermedad respectivamente.
Palabras clave: Manicata,genotipo, diversidad, pasiflora, frutas tropicales.
Abstract. In the Andean region of Nariño-Colombia, 99 materials were collected, 91 of the subgenus Tacsonia and 8 of Manicata. Seeds were sowing in a working collection, in order to preserve and make the morphoagronomic characterization “in vivo". The samples correspond to species: Passiflora tarminiana Coppens and Barney, Passiflora tripartita var. mollissima (Kunt) Bailey, Passiflora tripartita (Juss) Poir, Passiflora mixta L.F., Passiflora cumbalensis (Karst) Harás, Passiflora pinnatistipula Cav, Passiflora x rosea Karst., Passiflora manicata (Juss) Pers and accessions BGP BGP 63 and 76. The 64.8% of the genotypes are located in the South, highlighting the municipality of Aldana with more diversity. The principal component analysis (PCA) and multiple correspondence analysis (MCA), allowed to characterize the samples and form groups by hierarchical characterization method. ACP was formed in the 5 factors that explain 66.30% of the total variation assessed the hierarchical groupings of 4 classes, the features with the greatest contribution to variability were: weight fruit, length peduncle, the first group is standing out. The ACM showed 50.60% of the total variability in the first 5 components, the classification analysis to discriminate 5 groups of species, the characters that most contributed to the variability were: fruit shape and presence of anthracnose Colletotrichum gloeosporioides (Penzing) Penzing and Saccardo, P. Tarminiana and P. Tripartite groups were characterized by the absence and presence of the disease respectively.
Key words: Manicata, genotype, diversity, passiflora, tropical fruits.
Tacsonia, subgénero al que pertenecen las curubas, es uno de los más representativos dentro de la familia Passiflorácea; comprende aproximadamente 50 especies de alto-andino (Coppens, 1993). De éstas, 21 se encuentran en Colombia según la lista publicada por Hernández y Bernal (2000). Se localizan principalmente en la cordillera oriental extendiéndose ampliamente en los departamentos de Boyacá, Cundinamarca, Valle, Nariño, Cauca, Tolima, Antioquia, Caldas y Santander (Hansen, 1987).
La reciente clasificación, de Feuillet y Mac Dougal (2007), considera a Tacsonia una supersección, que agrupa las siguientes especies y subespecies: Passiflora tripartita var. mollissima (Kunth.) (Holm et al., 1988), conocida como curuba de Castilla, originaria de la zona andina de suramérica en donde se ha encontrado en condiciones silvestres (Campos, 1992), es la especie más conocida y cultivada; Passiflora tarminiana Coppens y Barney, conocida en Colombia como curuba india, descrita recientemente como una nueva especie (Coppens et al., 2001). Passiflora pinnatistipula, un bejuco ornamental de grandes flores rosado-violáceas y una fruta comestible (Uribe, 1940); Passiflora tripartita Juss Poir. Entre las subespecies que también son de valor comercial, pero con frutos de menor sabor y aroma, se tienen: Passiflora cumbalensis Karst que difiere de P. mollissima en la forma de la hoja, las flores azules o púrpuras y el fruto de color rojo; Passiflora mixta (curubito indio) tiene gran importancia en la búsqueda de resistencia a enfermedades, y también en cruzamientos interespecíficos; Passiflora antioquensis Karst y Passiflora bracteosa Planch. y Lind. ex Tr. y Planch., consideradas fuera del subgénero en las clasificaciones anteriores, se encuentran actualmente incluidas. Además Passiflora manicata (Juss.) Pers. (subgénero Manicata) aparece relacionada con Tacsonia.
Dentro del mejoramiento de las Passifloras, Escobar (1991), afirma que el entrecruzamiento constituye un método económicamente viable en la supersección Tacsonia para mejorar las especies, debido a que los representantes de este subgénero son autocompatibles mostrando una gran capacidad de cruces interespecíficos, con gran éxito y por tanto se puede incorporar en ellas características deseadas en las cultivadas, una más amplia tolerancia a las condiciones ambientales y mayor resistencia a enfermedades. De esta manera se han realizado estudios en el mejoramiento de curuba efectuado cruzamientos entre P. mollissima con especies de Passiflora, pero sin previo conocimiento sobre la variabilidad intraespecífica y las afinidades entre especies (Schoëniger, 1986; Escobar, 1985).
El estudio de la diversidad genética se apoya en la caracterización por medio de descriptores morfológicos, isoenzimáticos o marcadores moleculares. La caracterización morfológica pretende mostrar atributos que diferencian o relacionan las plantas evaluadas. Se basa en variables cualitativas de alta heredabilidad y características cuantitativas, generalmente de media a baja heredabilidad. Es importante tener en cuenta que la mayoría de los genes útiles para mejoramiento se encuentran en las especies silvestres, puesto que la variabilidad genética es una estrategia de supervivencia en condiciones naturales (Chaparro, 1993). La conservación de estos genes y la información obtenida de sus características, serán de gran importancia para los fítomejoradores, quienes podrán elegir progenitores con un conocimiento más amplio de las bondades de los caracteres que poseen. Además, las distancias genéticas ayudarán a los mejoradores a visualizar los genotipos con los cuales se pueden realizar cruzamientos que permitan obtener una favorable trasgresión genética.
Desde el punto de vista morfológico se han hecho publicaciones acerca de la diversidad genética del subgénero Tacsonia. Restrepo y Aristizábal (1998), evaluaron 12 accesiones provenientes de colectas e introducciones del Departamento de Nariño en donde se pudieron distinguir claramente cuatro materiales que corresponden a los subgéneros Tacsonia y Manicata, esta caracterización permitió identificar la alta variabilidad genética que se puede tener dentro del género Passiflora, especialmente entre estos subgéneros donde se puede considerar la posibilidad de realizar mejoramiento genético. Por su parte Primot et al. (2005) estudiaron la variación morfológica de ocho accesiones de las tres principales especies de curubas cultivadas y silvestres del Valle del Cauca, P. mollissima, P. tarminiana y P. mixta y 11 de sus híbridos, este estudio les permitió separar claramente las tres especies. Dentro de P. mixta, encontraron que se distinguen un tipo representativo de las plantas silvestres más comunes, y un tipo con importante variación morfológica y rasgos de curuba de Castilla, indicando flujos de genes entre P. mollissima y P. mixta. Los estudios más generales de Fajardo et al. (1998) y Sánchez et al. (1999), con marcadores RAPDs y RFLP, permitieron separar P. tarminiana de P. mollissima, mostrando también una fuerte afinidad, pero no incluían accesiones de P. mixta. En la investigación de Maya y Mipaz (2007), acerca de la caracterización morfológica in situ de Passifloráceas en el municipio de Pasto, encontraron que el subgénero Tacsonia presentó poca divergencia entre sí; pero tuvo alta variabilidad en caracteres referentes al fruto como: forma, sabor, color y dureza o firmeza de la cáscara.
No obstante en los trabajos reportados dentro de las Passifloras del subgénero Tacsonia, no se ha hecho una exploración amplia en el Departamento de Nariño que permita establecer su diversidad con base en un representativo número de accesiones procedentes de los diferentes municipios de clima frío y frío moderado. Además sin un programa de conservación específico algunos de ellos corren el riesgo de desaparecer y con ellos los genes que poseen, los cuales pueden ser requeridos para resolver problemas actuales y futuros en la explotación de este tipo de fruta.
El objetivo del presente trabajo fue realizar la colección y caracterización morfoagronómica de las Passifloras del subgénero Tacsonia y especies afines encontradas en la zona andina del Departamento de Nariño. La información recopilada en la caracterización de las variables cuantitativas y cualitativas, permitirá explorar su potencial genético e implementar un programa de mejoramiento de la curuba en Nariño.
MATERIALES Y MÉTODOS
La colecta se realizó entre los meses de abril hasta agosto del 2006, en los diferentes municipios de la zona andina del departamento de Nariño, ubicados entre los 1.800 a 3.200 msnm, dividiendo la región andina en cinco zonas. Los genotipos se colectaron principalmente a orillas de camino, bosques, linderos, patios caseros y pequeñas siembras comerciales, tomando de cadaplanta un fruto maduro. Para cada genotipo colectado se llevó un registro en forma individual con los datos de pasaporte. La ubicación del lugar donde fueron encontradas las muestras se determinó mediante GPS (Global Position System).
Establecimiento de la colección de trabajo y de su conservación in vivo. La colección se sembró en la granja experimental de Botana perteneciente a la Universidad de Nariño, Municipio de Pasto, Colombia, a una altitud de 2.820 msnm, temperatura promedio de 13°C, y una humedad relativa de 85%.
Se establecieron dos bloques de 53 surcos cada uno; con 12,5m de largo de tal forma que a cada accesión le correspondió un surco,en cada surco se ubicaron cinco plantas a una distancia de 2,5m y se utilizó una distancia entre surcos de 2m. La caracterización morfoagronómica se realizó cuando las plantas alcanzaron la primera floración, registrando las características cuantitativas y cualitativas, con base en los descriptores propuestos por el IPGRI, para Passifloras del subgénero Tacsonia; los datos cuantitativos, se tomaron del promedio de las 5 plantas de cada surco y los datos cualitativos correspondieron a la moda.
Análisis estadístico. La información cuantitativa recolectada para cada uno de los materiales se trabajó con análisis de componentes principales (ACP) y las características cualitativas con análisis de correspondencia múltiple (ACM). Antes de someter los datos de las variables cuantitativas, al ACP, se eliminaron todas aquellas variables que mostraron bajos índices de variabilidad, tomando como criterio el coeficiente de variación (<30%) por ser un indicativo de baja variabilidad. Además, mediante la matriz de correlación de Pearson, se seleccionaron las variables que mostraron independencia, de tal manera que cuando dos variables presentaron correlación alta y significativa, solo una de ellas se incluyó en el análisis. Los datos fueron procesados mediante la utilización del software Spad 3.5. (CISIA - CERESTA, 1998).
RESULTADOS Y DISCUSIÓN
Mediante el estudio taxonómico de las accesiones colectadas que serealizó en el herbario de la Universidad de Nariño, se identificaron las siguientes especies: Passiflora cumbalensis (Karst.) Harás, Passiflora mixta L.F, Passiflora tripartita var. mollissima (Kunth) Bailey, Passiflora tarminiana Coppens y Barney, Passiflora tripartita var. tripartita, Passiflora manicata (Juss) Pers., Passiflora pinnatistipula Cav. y Passiflora x rosea. Además las accesiones BGP 63 y BGP 76, que están en proceso de identificación.
Distribución geográfica. Se colectaron un total de 99 genotipos, ubicados entre los 2.000 msnm y 3.232 msnm, de éstos el 64,08% se encontraron en la zona sur, correspondiendo el mayor número de colectas a los municipios de Ipiales, Pupiales y Aldana con 7,77%, 6,80% y 6,80% respectivamente, este último municipio mostró alta diversidad presentándose cuatro de las 10 especies encontradas: P. tarminiana, P. tripartita var. mollissima, P. pinnatistipula y P. x rosea.
Descripción morfológica de las plantas colectadas del subgénero Tacsonia. (Figura 1)

Figura 1. Especies de Tacsonia colectadas en la zona andina del departamento de Nariño, Colombia: A. Passiflora tarminiana Coppens y Barney, B. Passiflora tripartita var. mollissima (Kunth) Bailey, C. Passiflora tripartita var. tripartita, D. Passiflora mixta L.F, E. Passiflora cumbalensis (Karst.) Harás, F. Passiflora pinnatistipula Cav., G. Passiflora x rosea, H. Passiflora manicata (Juss) Pers., I. BGP 63 y J. BGP 76.
Passiflora tarminiana Coppens y Barney. Es la especie más difundida en la zona andina del Departamento de Nariño, debido a que se encontró en la mayoría de los municipios visitados, destacándose Córdoba y el Tambo, lugares en donde se cultiva comercialmente. Sus hojas son glabras y lustrosas en el envés y las flores se abren ampliamente de forma reflexa (hacia atrás). Además el fruto es alargado y fino (Imagen A).
Passiflora tripartita var. mollissima (Kunth) Bailey. Subespecie de Passiflora tripartita (Juss) Poir, se localizó en 11 municipios de las zonas 1, 2 y 3. Se caracteriza por ser una liana vigorosa, muy pubescente. La flor es péndula, con una corola campanulada de color rosado carmesí, que da un fruto oblongo de color amarillo al madurar, y un arilo suculento. Uno de los principales problemas fitosanitarios es la antracnosis, enfermedad que deprecia los frutos y puede afectar el follaje (Imagen B).
Passiflora tripartita var. tripartita (Juss) Poir. Se localizó en 9 municipios de las zonas 1, 2 y 3, de la región andina del departamento de Nariño. Posee hojas glabras en la parte superior de la superficie, el fruto es más fusiforme y de un amarillo más profundo, a veces tintado de rosado, con una pulpa un poco menos aromática, pero menos astringente que la de P. mollissima (Imagen C).
Passiflora mixta L.F. Se encuentra distribuida en el municipio de Pasto de forma silvestre de acuerdo a lo expuesto por Maya y Mipaz (2007); son plantas con tallos glabros o pubescentes, sus pedúnculos son más rígidos que mantienen generalmente las flores en posición parcialmente erecta y los frutos son de coloración amarillo verdoso al madurar (Imagen D).
Passiflora cumbalensis (Karst.) Harás. Al igual que P. mixta está distribuida a lo largo de la zona rural del municipio de Pasto, presenta tallos glabros, hojas pequeñas y coriáceas; posee flores de color violáceo azulado; el hipanto de color azul morado caracterizado por ser más largo y angosto que en las otras variedades; los frutos son pequeños ovoides de sabor dulce insípido y contienen aproximadamente tres veces más semillas que los frutos de P. mollissima (Imagen E).
Passiflora pinnatistipula Cav. Se ubica en la zona sur, especialmente en los municipios de Aldana, Sapuyes e Iles. Es una planta trepadora leñosa de flores rosadas, lilas o rojizas, con el envés de los pétalos tintado de blanco. Sus hojas son trilobadas y sus estípulas pinnadas y divididas en hilillos de ahí su nombre; el fruto es redondo o subglobuloso, con un pericarpio delgado y quebradizo de color verde grisáceo o amarillo. La pulpa es grisácea hasta amarillenta, dulce o levemente ácida, muy perfumada (Imagen F).
Passiflora x rosea Vaz. Es un híbrido natural interespecifico de P. pinnatistipula con P. tripartita var. mollissima. Se forma muy fácilmente cuando las dos especies están cerca en una misma área, se encontró en el municipio de Aldana. Es muy parecida a P. pinnatistipula, de la cual se distingue por la longitud de las anteras y del ginóforo, equivalente a la del androginóforo, las estípulas más foliáceas y las brácteas coalescentes en su base. Los frutos son un poco más alargados y su arilo es amarillo-grisáceo (Imagen G).
Passiflora manicata (Juss) Pers. Arbusto muy variable perteneciente al subgénero Manicata. Se localizó en los municipios de Funes e Imues, en estado silvestre. De especial interés para mejoramiento genético. Posee flores de copa floral reflexa, con orientación erecta, de color rojo; el fruto es ovoide con un pericarpio coriáceo, verde cuando madura y el arilo es grisáceo y poco suculento. Difiere de las especies del subgénero Tacsonia por un tubo floral más corto, una corona filamentosa compleja y su adaptación a sitios menos elevados (Imagen H).
Accesión BGP 63. Este ejemplar se encontró en el municipio de Carlosama, vereda de Macas y actualmente está en proceso de identificación. Es una enredadera que se caracteriza porque presenta polimorfismo foliar y heteroblastia, sus hojas alcanzan los 15 cm de longitud y su color en el haz es verde oscuro brillante; el tallo es glabro; la flor es similar a la de P. tripartita var. mollissima, aunque en el fruto presenta características de P. tarminiana, en la forma, sabor y resistencia a antracnosis Colletotrichum gloeosporioides (Penzig) Penzigy Saccardo (Imagen I).
Accesión BGP 76. Se localizó en el municipio de Iles, vereda San Javier. Es una planta enredadera, posee tallos gruesos y pubescentes; las hojas son trilobuladas poco pubescentes de color verde claro; la flor se asemeja a la de P. mixta pero difiere al presentar tubérculos que oscilan entre 2 y 7 mm; el fruto es la característica más destacable debido a que tiene forma de gota, es resistente a Colletotrichum gloeosporioides, mide no más de 8 cm de largo, es de color amarillo-rojizo cuando maduro y con un arilo muy suculento y perfumado. Al igual que la anterior también está en proceso de identificación (Imagen J).
Análisis de componentes principales (ACP) para variables cuantitativas. A partir del descarte de variables con base en el coeficiente de variación (<30 %) se obtuvo un total de 32 variables y mediante la realización de la matriz de correlación de Pearson se determinaron asociaciones altas (= 0,70) entre las variables (longitud de la primera hasta la segunda bifurcación en el pedúnculo) LPSB y (longitud de la primera base del pedúnculo hasta la segunda bifurcación en el pedúnculo) LBPEPB; LPSB y (número de dientes en el margen foliar) NDMF. Además la variable (número de bifurcaciones en el pedúnculo) NBP, se correlacionó con (longitud del pedúnculo) LPEDU. Estas correlaciones están explicadas dada la morfología exhibida en el pedúnculo y el pedicelo por las plantas del subgénero Tacsonia, las cuales no presentan bifurcaciones en el pedúnculo; por lo tanto, las dimensiones evaluadas hacen referencia a la misma parte del pedúnculo en cuestión. Con este análisis se seleccionaron las variables que mostraron independencia, LPSB y LPEDU, las cuales se incluyeron en el ACP. Para este se dispuso de 29 variables, 19 de ellas activas.
El ACP permitió establecer cinco factores que explican el 66,30% de la variación total evaluada. Esto indica que puede reducirse la dimensión del problema a cinco componentes con baja perdida de información (33,7%).
En la Tabla 2 se muestran las estimaciones de los valores propios y la proporción de la variación total explicada por cada uno de los componentes principales elaboradas con base en las 19 características seleccionadas.
Tabla 2. Valores propios y varianza explicada por los componentes resultantes del ACP realizado a materiales del subgénero Tacsonia para las variables cuantitativas, en el departamento de Nariño, Colombia.
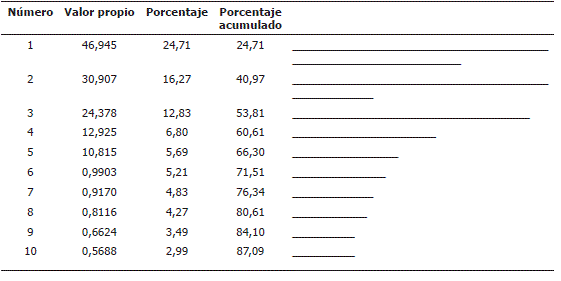
El primer componente explica el 24,71% de la variabilidad y como se observa en la Tabla 3, está correlacionado principalmente por variables referentes a tallo, estípulas, flor y fruto estos son: NSFC (0,81), PSFF (- 0,78), LEN (- 0,62), AHE (0,59) y LE (longitud entrenudos). El factor dos, con una explicación del 16,27% de la variabilidad observada, está conformado básicamente por variables relacionadas el pedúnculo y pedicelo: LPEDU (0,77), LPE (-0,71), NNP (-0,68) y DIPSLCE (- 0,63). Las variables que más aportaron al tercer factor el cual explicó el 12,83% de la variación total presentada por las accesiones está compuesto principalmente por caracteres del pecíolo y flor estos son: LL (- 0,83), LFTL (-0,81) y LBPPN (0,58). El cuarto factor mostró 6,80% de explicación del total de la variabilidad y está conformado fundamentalmente por caracteres de la flor: NSFTH (0,42) y OFG (-0,34). Finalmente el quinto componente principal que explica un 5,69% de la variación lo integran variables que aluden a hojas, estípulas y flor: DBFINS (0,54), NSFTH (0,39) y AHE (-0,38).
Tabla 3. Contribución de 19 variables cuantitativas a la conformación de los primeros cinco factores de la colección de plantas del subgénero Tacsonia, en el departamento de Nariño, Colombia.
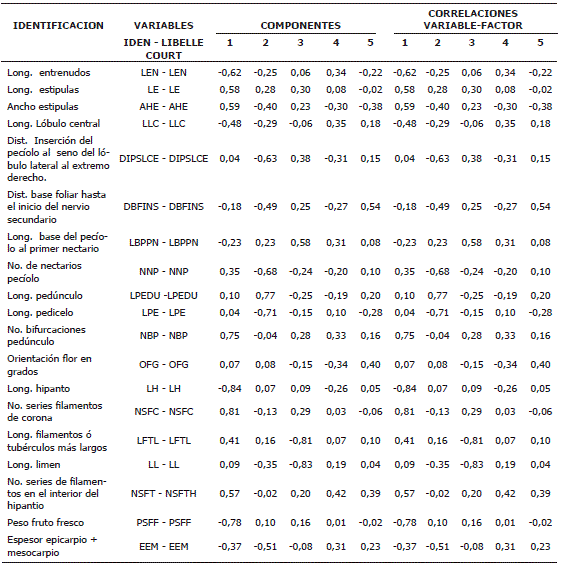
Con lo anterior se evidencia que los cinco componentes comprenden características o variables tanto de partes vegetativas como tallo, hojas, pedúnculo, pedicelo, brácteas, y variables de partes como la flor y el fruto.
Análisis de clasificación para las variables cuantitativas. El análisis de clasificación, basado en las características cuantitativas, se realizó con la conformación de cuatro clases en un dendograma (Figura 2), con el fin de establecer en el subgénero Tacsonia, diferencias en los niveles de divergencias morfológicas entre las especies y las secciones, evidentes por la formación de grupos con alto grado de similitud entre las accesiones que lo componen.
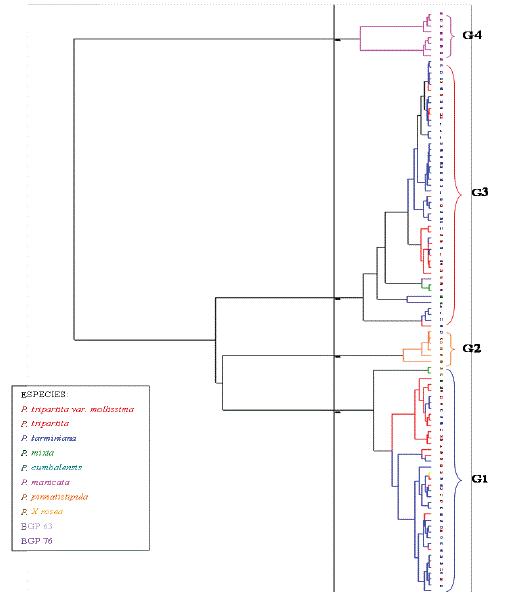
Figura 2. Clasificación jerárquica para variables cuantitativas de la colección de Tacsonias de la zona andina del departamento de Nariño, Colombia.
El primer grupo fue conformado por 38 accesiones que representan el 38,38% de la colección y corresponden las siguientes especies: P. tarminiana, P. mollissima, P. tripartita, la accesión BGP 76, P. pinnatistipula, P. cumbalensis y P. mixta (21, 12, 2, 1, 1 y 1 individuos respectivamente). Se destacan por presentar un promedio superior en la longitud del pedúnculo (35,02 mm) comparado con el promedio general del total de las accesiones que fue de 20,94 mm, dentro de este ítem las accesiones de P. mollissima de Funes, P. manicata de Imues y P. Cumbalensis de Pasto fueron las que presentaron los pecíolos más largos. Por otra parte mostraron características agronómicas sobresalientes como el peso superior de sus frutos (PSFF= 84,54 g) en contraste con el promedio general (74,65 g), destacándose dentro del conjunto las accesiones P. tarminiana de Túquerres y P. tripartita de Ipiales; así mismo las accesiones de esta clase mostraron mayores dimensiones en las variables referentes a la flor y al pecíolo como son la longitud del hipantio (LH = 71,32 mm), y la longitud de la base del pecíolo al primer nectario (LBPPN = 12,30 mm).
Desde el punto de vista agronómico este grupo posee como característica favorable para la producción y comercialización, el mayor peso del fruto. Restrepo y Aristizábal (1998) afirman que son de calidad comercial los frutos con peso igual o superior a 60 g y en consecuencia las accesiones incluidas en este agrupamiento se pueden tener en cuenta en un programa de mejoramiento encaminado a incrementar el tamaño del fruto.
Por otra parte en esta clase existe una marcada proximidad entre individuos de P. tarminiana, P. mollissima, P. tripartita, P. cumbalensis y P. mixta, constituyendo grandes subconjuntos dentro de Tacsonia, al parecer por un constante flujo de genes entre especies dada por la cercanía regional. La proximidad de P. mollissima y P. tarminiana, apoya la idea de que éstas son dos subespecies o variedades botánicas de un mismo taxón o que constituyen una divergencia en proceso de especiación, en este sentido, Fajardo et al. (1998); Sánchez et al. (1998), indicaron que las dos especies se agrupaban estrechamente a nivel molecular. Igualmente la aparición de híbridos espontáneos y reportados regularmente, tanto en forma silvestre como cultivada, sugiere barreras interespecíficas débiles, por lo que su existencia aumenta el riesgo de confusión relacionado con la multiplicidad de las especies dentro del subgénero. No obstante a veces los rasgos morfológicos son susceptibles de modificaciones debidas al ambiente (Segura, 2003), por lo que otro tipo de descriptores como los moleculares podrían indicar la presencia o ausencia de estos fenómenos.
El grupo dos está compuesto por seis genotipos que representan el 6,06% del total de la colección, cinco de los individuos de esta clase corresponden a P. pinnatistipula y uno a P. x rosea. Este híbrido tiene una flor más pequeña que P. mollissima, estípulas y hojas más grandes que P. pinnatistipula, pero en el dendrograma Passiflora x rosea se agrupa con las accesiones de P. pinnatistipula debido a su marcada similitud morfológica global. En el dendograma se sitúan de forma intermedia entre los grupos 1 y 3, con más afinidad hacia el primero, se caracterizan por presentar la longitud de los filamentos o tubérculos en la flor (LFTL = 18,52 mm) superiores a los del promedio general que fue de 3,32 mm; además presentan las longitudes del limen y pedúnculo (LPEDU = 53,85 mm; LL = 5,04 mm) más amplias que las medias generales de 20,94 mm y 5,04 mm respectivamente. También presentaron una longitud del hipanto (LH = 41,48 mm) y un peso de fruto (PSFF =43,65 g) inferiores a los promedios de toda la colección; 63,74 mm y 74,65 g respectivamente. Los caracteres como el tubo floral intermedio y la presencia de una corona sencilla pero filamentosa la situaron en posición casi intermedia entre los subgéneros Passiflora y Tacsonia (Killip, 1938).
El tercer grupo es el más numeroso, está compuesto por 47 genotipos (44,44%) y al igual que en el primer grupo existe una diversidad de especies: 30 de P. tarminiana, 6 de P. mollissima, 9 de P. tripartita, el hibrido BGP 63, 1 de P. mixta y 1 de P. cumbalensis. Se distinguen por presentar una longitud del pedicelo (LPE = 34,40 mm) y un espesor del epicarpio más el mesocarpio (EMM = 4,68 mm) más amplios que los promedios generales de la colección que en su orden son: 23,94 mm y 3,84 mm, siendo la clase con la cáscara más gruesa. Otra característica sobresaliente es que el grupo tiene las flores más largas (LH = 68,99 mm) que el resto de la colección.
Por último el grupo 4 se conformó por 8 accesiones (8,88%), todas pertenecientes a P. manicata, los cuales se agrupan por poseer un mayor número de series de filamentos de corona (NSFC = 2,27 mm) en contraste con la media (1,15 mm). También presentan más bifurcaciones en el pedúnculo (NBP = 1,63) en comparación con el promedio de la colección (1,05). Otra característica importante de esta clase es que presentó un mayor número de dientes en las hojas, el cual alcanzó los 10,55 dientes en 1 cm, muy superior a lo presentado por las otras accesiones que fue de 5,55. En cuanto a flores estas accesiones presentaron valores inferiores en la longitud del hipanto, siendo éste el grupo con la flor más pequeña. En el dendograma este grupo se presentó distante de las otras especies aunque P. manicata tiene cierta afinidad para cruzamientos con las especies del subgénero Tacsonia. En este sentido Escobar (1985) reportó hibridaciones de P. mollissima con P. manicata. El interés de esta especie proviene de su potencial para el mejoramiento genético de las curubas, por su cercanía con ellas y su rusticidad debido a que es resistente a los nemátodos y a las enfermedades fúngicas (antracnosis, oidio y Alternaria) (Campos, 2001).
Análisis de correspondencias múltiples (ACM). El ACM permitió seleccionar 5 componentes que explican el 50,60% de la variabilidad total; sobresale el primer factor que representa el 19,71% de la variabilidad, seguido del factor dos con 12,40%. Los factores 3, 4 y 5 explican el 8,55%, 5,64% y 4,36% respectivamente (Tabla 4). Observando las variables más sobresalientes, predominaron los genotipos con pubescencia de la copa floral glabra PubCopF (92 accesiones) al igual que el hipanto PubHip (V=15) (91 accesiones); de tipo de crecimiento de enredadera TipCrec (91 accesiones) y con una superficie de semillas reticulada TipSuS (90 accesiones).
Tabla 4. Valores propios y distribución de la varianza explicada por los componentes resultantes del ACM realizado para variables cualitativas en la colección de Tacsonias en el departamento de Nariño, Colombia.
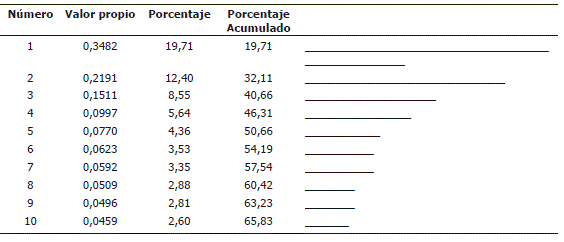
El análisis de contribuciones de las variables cualitativas muestra la conformación de cada uno de los ejes factoriales (Tabla 5). Las variables que aportaron a la conformación del factor uno son: color de los filamentos estaminales (CoFiEs = 7,8%), color de pétalos (ColDPet = 6,9%), color del androginóforo (ColAnd = 6,9%), tipo de crecimiento (TipCrec = 6,5%) y forma del fruto maduro (ForFruM = 6,3%).
Tabla 5. Contribución de las variables cualitativas evaluadas en la colección de Tacsonias en el departamento de Nariño, Colombia, a la conformación de los primeros 5 ejes factoriales.
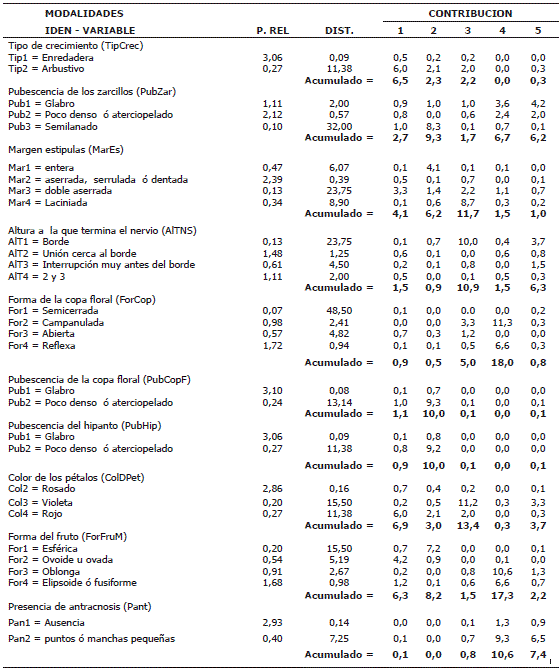
El segundo eje factorial está compuesto por las densidades de pubescencia tanto de la copa floral (PubCopF = 10%), del hipanto (PubHip = 10%) como de los zarcillos (PubZar = 9,3%). Las especies P. pinnatistipula y P. x rosea son las que presentan más alta densidad de pubescencia.
El eje tres está conformado principalmente por las variables: color de los pétalos (ColDPet = 13,5%), en donde se encuentran un total de 84 accesiones con flores de color rosa, 6 de color violeta, 6 de color rojo y 1 de flor color púrpura; margen de las estípulas (MarEs = 11,8%) con 71 individuos con estípulas de margen aserrada, 14 individuos con margen entera, 10 con margen dentada y 4 con estípulas de margen serrulada y AlTNS (10,9%) donde se destacan 44 genotipos en donde el nervio secundario termina en unión cerca al borde (V9=2).
La forma de la copa floral (ForCopF = 17,4%) es la variable que más aporta al cuarto eje, dado que se presentan 51 individuos con copa floral reflexa, 29 con copa floral campanulada, 17 con copa floral abierta y dos con copa semicerrada. También a este eje contribuyó la variable ForFruM (16,4%) en donde 50 de las accesiones presentaron fruto en forma elipsoide. Otra variable que aportó a la conformación del eje fue el color de los sépalos con un 11,0%, la cual presentó 66 individuos con sépalos de color rosado. El quinto eje se conformó principalmente por la variable tipo de superficie de la semilla (TipSupS) con 18,8% de aporte, seguido por la orientación de la flor (OrFlo = 9,8%).
Análisis de clasificación para las variables cualitativas. La categorización cuantificó cinco clases (Figura 3), donde fue posible diferenciar cinco grupos de especies. En este caso a diferencia del ACP realizado de este estudio, con las variables cualitativas si fue posible separar gran parte de las accesiones de P. tarminiana, el grupo de las subespecies de P. tripartita, y el de P. mixta con P. cumbalensis. En este sentido Villacis et al. (1998) observaron la misma separación interespecífica con descriptores cualitativos, caracterizando germoplasma ecuatoriano, pero no pudieron conseguir resultados coherentes con los descriptores cuantitativos, lo que atribuyeron a la composición de la muestra, el balance entre caracteres vegetativos y florales y el posible efecto de la selección artificial sobre caracteres cuantitativos.
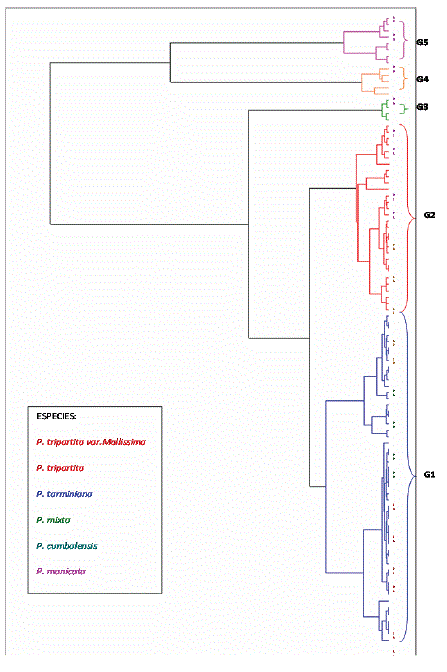
Figura 3. Clasificación jerárquica de las variables cualitativas de la colección de Tacsonias de la zona andina del departamento de Nariño, Colombia.
El primer grupo se conformó por 53 accesiones que representaron el 53,54% del total de la colección. En esta clase el 96% (48 individuos) son accesiones de P. tarminiana, 2 de P. mollissima, 2 de P. tripartita y 1 de P. pinnatistipula. Se caracterizan porque el 98% de las accesiones presentan frutos en forma elipsoide o fusiforme, el 86,27% de los genotipos tienen una forma reflexa de la copa floral y el 75,76% tiene el color de los sépalos rosado. También el 62,20% de los individuos de este grupo mostraron frutos de color amarillo y el 58,82% de los individuos no presentaron antracnosis Colletotrichum spp. Las características de fruto y resistencia observadas en este grupo las hacen clasificar como las mejores Tacsonias dentro de la colección, destacando de manera importante la presencia de P. tarminiana la cual es particularmente interesante por su rusticidad y resistencia a la antracnosis Colletotrichum gloeosporioides (Penzig) Penzig y Saccardo (Campos, 2001). La antracnosis del fruto es considerada una de las enfermedades más importantes de la curuba en los departamentos de Antioquia, Boyacá, Caldas, Cundinamarca, Nariño, Santander, Tolima y Valle del Cauca. Es causada por el hongo Colletotrichum gloesporioides y produce pérdidas superiores al 50% de la fruta cosechada (Tamayo, 2005). Accesiones de P. tarminiana, pueden ser de utilidad para el mejoramiento de curuba por resistencia a antracnosis.
El grupo dos está conformado por 29 genotipos que representan el 29,29% de la población total, hacen parte de esta clase 16 accesiones de P. mollissima, 9 de P. tripartita, 2 de P. tarminiana y las dos accesiones BGP 63 y BGP 76. El 85,19 % de plantas que componen esta clase tienen una forma de fruto oblonga, la forma de la copa floral del 79,31% de los individuos es campanulada (V12=2). Se observó la alta susceptibilidad de P. mollissima a la antracnosis Colletotrichum spp, con el 76,92% de los individuos de este grupo afectados por esta enfermedad, siendo este uno de los principales problemas fitosanitarios que deprecia los frutos y puede afectar el follaje. Lo anterior confirma lo manifestado por Campos (2001), el cual afirma que P. mollissima se ve afectada por nemátodos del género Meloidogyne. Las accesiones de P. tripartita y las accesiones BGP 63 y BGP 76 mostraron resistencia a dicha enfermedad. En el dendograma se ubican entre los grupos de P. tarminiana y el de P. mixta y P. cumbalensis, con mas cercanía hacia el primero, asegurando la compatibilidad entre las especies y la existencia de un flujo natural de genes lo cual abre interesantes perspectivas para el mejoramiento de la curuba de Castilla.
El tercer grupo compuesto por cuatro accesiones que representan el 4,04% del total de la colección, se caracterizan porque el 75% de los individuos que lo conforman presentan el nervio secundario terminado en el borde de la hoja, además el 66,67% de los individuos son de pétalos color violeta. Conforman este grupo accesiones de P. mixta y P. cumbalensis, estas especies tienen un fuerte parecido morfológico, aunque son de dos secciones diferentes, P. mixta de la sección Tacsonia y P. cumbalensis de la sección Bracteogama. En el dendograma se agrupan de forma independiente con cierta afinidad hacia la clase formada por P. tripartita, esto se explica debido a que P. mixta y P. cumbalensis han manifestado también un flujo de genes en especial hacia P. tripartita (Sañudo y Zúñiga, 1991). Además este grupo pudo distinguirse del resto de especies con base en las variables cualitativas y constituye un recurso genético para considerar en el mejoramiento de P. mollissima, por sus evidentes afinidades morfológicas con esta especie.P. cumbalensis es resistente a las enfermedades fúngicas (oidio, antracnosis, Alternaria), y P. mixta, también posee características de resistencia. En este sentido Escobar (1981) y Schoëniger (1986) obtuvieron híbridos fértiles al cruzar P. mixta y P. mollissima; Primot, Rioux y Coppens et al., (2001), concluyeron que existe una gran facilidad en los cruzamientos entre P. mollissima, P. tarminiana y P. mixta.
El cuarto grupo se compone por cinco accesiones que representan el 5,05% del total de la colección, cuatro de P. pinnatistipula y 1 de P. x rosea similar a lo presentado en el ACP. El 100% de los individuos del grupo mostraron zarcillos de pubescencia aterciopelada, de igual forma sucede con la pubescencia del tallo.La antocianina de las estípulas está presente en el 80% de las accesiones de este grupo. En cuanto a fruto el 66,67% de los individuos de esta clase muestran frutos redondos y con cáscara quebradiza. Las accesiones del grupo se ubican en una posición intermedia en el dendograma en donde se evidencia una proximidad hacia P. manicata por sus similitudes en la flor. Por lo anterior algunos autores colocan a P. pinnatistipula como fuera del subgénero Tacsonia, aunque cabe resaltar que en las clasificaciones actuales (Feuillet y MacDougal, 2007), esta especie hace parte del subgénero estudiado. Debido a sus características de rusticidad del endocarpio P. pinnatistipula es resistente a los hongos patógenos (Sañudo y Jurado, 1990), con lo cual se podría contribuir al objetivo de diversificación. Además, puede aportar a mejorar el rendimiento de pulpa, en las especies comerciales. Por su parte P. x rosea así como P. pinnatistipula, es resistente a Alternaria passiflorae (Sañudo y Jurado, 1990), pero su producción de frutos es generalmente muy limitada, aparentemente por la relativa incompatibilidad de los genomas parentales.
El grupo cinco se constituyó por ocho accesiones (8,08%) todas de la especie P. manicata; estos individuos se destacaron por ser de tipo de crecimiento arbustivo, de color de pétalos rojos, con margen en las estípulas doble aserrada y de color púrpura. También el 77,78% de los genotipos de este grupo tuvieron una orientación de la flor erecta y una dureza de la cáscara resistente a la presión. Las similitudes de la flor de P. manicata y P. pinnatistipula se reflejan por su proximidad, siendo ligeramente más cercanas al grupo de especies de Tacsonia silvestres, similar a lo obtenido en el análisis de componentes principales y al estudio realizado por Segura et al. (2003).
En general, los grupos obedecen a la clasificación taxonómica de los materiales evaluados, lo que indica la efectividad de las variables para distinguir las especies del subgénero Tacsonia. Además la caracterización morfológica muestra claras diferencias interespecíficas en la distribución de los caracteres cuantitativos y cualitativos con especial importancia de estos últimos, reflejándose en las clasificaciones obtenidas con los dos tipos de variables, donde se separan claramente las especies. Esta variabilidad genética presente en las especies estudiadas sugiere buenas posibilidades para mejorar y diversificar las curubas comercializadas actualmente. Donde se pueden explotar mejor los caracteres de resistencia existentes en P. tripartita, P. manicata, P. mixta y los tipos de P. cumbalensis, para beneficio de P. mollissima y P. tarminiana.
CONCLUSIONES
La mayor diversidad del subgénero Tacsonia en Nariño está en el municipio de Aldana.
Las variables cuantitativas que hicieron la mayor contribución a la variabilidad de las Passifloras del grupo Tacsonia colectadas en Nariño fueron: longitud de los filamentos o tubérculos más largos en la flor, longitud del pedúnculo, peso del fruto fresco, ancho y longitud de las estípulas y longitud del hipanto.
Las variables cualitativas que más aportaron a la variabilidad de las Passifloras del grupo Tacsonia colectadas en Nariño fueron: pubescencia de la copa floral y del hipanto, tipo de crecimiento, forma de la copa floral, color de pétalos, forma de fruto maduro y presencia de antracnosis.
Las accesiones de las especies P. mollissima, P. tarminiana, P. cumbalensis, P. mixta en los análisis de clasificación jerárquica para variables cuantitativas se integraron en los mismos grupos sugiriendo cercanía genética y flujo de genes entre ellas.
El análisis de clasificación jerárquica para variables cualitativas permitió hacer una separación más clara entre especies, sugiriendo patrones evolutivos diferentes de las variables cuantitativas respecto a las variables cualitativas.
BIBLIOGRAFÍA
Campos, T.J. 1992. El cultivo de la curuba Passiflora mollissima (H.B.K) Bailey en Colombia. Acta de Horticultura 310: 215-229. [ Links ]
Campos, T.J. 2001. La curuba. Cultivo. Instituto Interamericano de Cooperación para la Agricultura IICA, Bogotá. 87 p. [ Links ]
Coopens, G. 1993. Promesa de las passifloras. Boulevard de la Lironde, TA50/PS4, Montpellier, Francia, http://www.cefe.cnrs.fr/ibc/pdf/coppens/24_pasifloras%20Medellin2_Geo.htm; consulta: diciembre 2009. [ Links ]
Coppens, G., V. Barney, P. Moller and J. MacDougal. 2001. Passiflora tarminiana, a new cultivated species of Passiflora subgenus Tacsonia. Journal for Botanical Nomenclature 11(1): 8-15. [ Links ]
Chaparro, G.A. 1993. Biotecnología biodiversidad y patentes en la agricultura. Agricultura Tropical 30(1): 39-44. [ Links ]
CISIA – CERESTA. 1998. SPAD. Version 3.5. Manual de Prise en Main – Mande, France.
Escobar, L.A. 1981. Experimentos preliminares en la hibridación de especies comestibles de Passiflora. Actualidades Biológicas 10(38): 103-111. [ Links ]
Escobar, L.A. 1985. Biología reproductiva de Passiflora manicata e hibridación con la curuba, Passiflora mollisima. Actualidades Biológicas 14(54):111-121. [ Links ]
Escobar, L.K. 1991. La sistemática y evolución de las passifloras. pp. 51–54. En: Memorias. 1er Simposium Internacional de Passifloras. Palmira, Colombia.
Fajardo, D., F. Ángel, M. Grum, J. Tohme, M. Lobo, W. Roca and I. Sánchez. 1998. Genetic variation analysis of the genus Passiflora L. Using RAPDS Markers. Euphytica 101:341-347. [ Links ]
Feuillet, C. and J. Mac Dougal. 2007. Passifloraceae. pp. 270-281. In: Kubitzki, K. (ed.). The families and genera of vascular plants. Springer, Berlin. v. 9. [ Links ]
Hansen, H. 1987. Aspectos relevantes del manejo precosecha, cosecha y pos-cosecha en el desarrollo de las exportaciones de frutas tropicales. pp. 49-56 En: Cuellar S. (comp.). Producción, manejo y exportación de frutas tropicales de América Latina. Federación Nacional de Cafeteros de Colombia. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Ed. Presencia Ltda, Bogotá. 80 p. [ Links ]
Hernández, A. y R. Bernal. 2000. Lista de especies de Passifloraceae de Colombia. Biota Colombianam 1(3): 320-335. [ Links ]
Holm, L., P. Jorgensen and J. Lawesson. 1988. Passifloraceae. 31 p. En: Harling, G. y Andersson, L. (eds.). Flora of Ecuador. University of Göteborg, Copenague. 130 p. [ Links ]
IPGRI. 1998. Diversidad, Conservación y uso sostenible de los recursos genéticos de frutales nativos de América Tropical. Informe Final. Cooperación Técnica IPGRI-BID No.ATN/SF-4356-RG. [ Links ]
Killip, E.P. 1938. The american species of Passifloraceae. Publications of the Field Museum of Natural History, Chicago. Botany Series 19(1,2): 1-163. [ Links ]
Maya, J. y O. Mipaz. 2007. Caracterización morfológica de plantas del género Passiflora en el municipio de Pasto, departamento de Nariño. Trabajo de grado de Ingeniería Agrónomica. Facultad de Ciencias Agrícolas. Universidad de Nariño, Pasto, Colombia. 73 p. [ Links ]
Primot, S., V. Rioux, D. Coppens, F. Garcín y J. Ocampo. 2005. Variación genética de tres especies de curubas (Passiflora tripartita var. mollisima, P. tarminiana y P. mixta) y sus híbridos en el Valle del Cauca, http://garcin.francois.free.fr/images/articulo6-12.PDF.11 p.; consulta: noviembre 2009. [ Links ]
Restrepo, J. y J. Aristizábal. 1998. Descripción de germoplasma e identificación de accesiones promisorias para el mejoramiento genético en Passifloras. Agronomía. Facultad de Ciencias Agropecuarias. Universidad de Caldas 8(1): 29-35. [ Links ]
Sánchez, I., F. Ángel, M. Grum, M. Duque, M. Lobo, J. Tohme y W. Roca. 1999. Variability of chloroplast DNA in the genus Passiflora L. Euphytica 106(1): 15-26. [ Links ]
Sañudo, S. y D. Jurado. 1990. Búsqueda de fuentes de resistencia a enfermedades fungosas de la curuba en Nariño. ASCOLFI Informa 16:3. [ Links ]
Sañudo, S. y R. Zuñiga. 1991. Híbridos interespecíficos de curuba resistentes a la antracnosis Colletotrichum gloeosporioides (Penz.) Sacc. en el departamento de Nariño. ASCOLFI Informa 17:9-10. [ Links ]
Schoëniger, G. 1986. La curuba. Técnicas para el mejoramiento de su cultivo. Editora Guadalupe Ltda., Bogotá. 255 p. [ Links ]
Segura, S., G. Coppens, C.H. Ocampo, and P. Ollitrault. 2003. Isozyme variation in Passiflora subgenera Tacsonia and Manicata. Relationships between cultivated and wild species. Genetic Resources and Crop Evolution 50(4): 417-427. [ Links ]
Tamayo, P. 2005. Tecnología para el cultivo de la curuba. Manual técnico No 6. pp. 101-131. Corporación Colombiana de Investigación Agropecuaria. CORPOICA, Rionegro, Antioquia. [ Links ]
Uribe, U.L. 1940. Flora de la real expedición botánica del nuevo reino de granada; Pasifloráceas y Begoniáceas. Colcultura Hispánica, Madrid. Tomo 17. [ Links ]
Villacis, L., J. Vega, M. Grum and G. Coppens. 1998. Caracterización morfológica de passifloras andinas del Ecuador. Plant Genetic Resources Newsletter 115: 51-55. [ Links ]

