










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Facultad Nacional de Agronomía Medellín
Print version ISSN 0304-2847
Rev. Fac. Nac. Agron. Medellín vol.64 no.2 Medellín July/Dec. 2011
Relaciones de Competencia entre el Fríjol Trepador (Phaseolus vulgaris L.) y el Maíz (Zea mays L.) Sembrados en Asocio
Study Interactions of Competition between Climbing Common Bean (Phaseolus vulgaris L.) and Corn (Zea mays L.) Plants Sowed Associate
León Darío Vélez Vargas1; Andrea Moya Muñoz2 y León Jairo Clavijo Porras3
1 Profesor Asistente. Universidad Nacional de Colombia - Sede Medellín - Facultad de Ciencias Agropecuarias. A.A. 1779, Medellín, Colombia. <ldvelez@unal.edu.co>
2 Ingeniera Agrónoma. Coordinadora Parque Temático Chaquen. Hospital Nazareth I nivel E.S.E. Calle 48 No. 73-55, Bogotá, Colombia. <andrea.moya@esenazareth.gov.co>
3 Profesor Asociado. Universidad Nacional de Colombia - Sede Bogotá - Facultad de Agronomía. A.A. 14490, Bogotá, Colombia. <jfclavijop@unal.edu.co>
Recibido: Agosto 2 de 2010; aceptado: Octubre 12 de 2011.
Resumen. Una forma de cultivar fríjol trepador es en asocio con maíz. Su principal característica es el mayor rendimiento por unidad de área con respecto a los unicultivos de cada especie, aunque la competencia que se presenta entre ellas reduce los rendimientos del fríjol en más del 40% y los de maíz en 20%. La investigación sobre los procesos fisiológicos afectados por la competencia y su dinámica ha sido escasa. El propósito de esta investigación fue evaluar los efectos de la competencia sobre el comportamiento fisiológico del fríjol. En un diseño de bloques completos al azar con cuatro repeticiones se evaluaron los siguientes tratamientos: fríjol sin competencia, fríjol y maíz competiendo por luz, por recursos del suelo y simultáneamente por luz y recursos del suelo. Las variables medidas fueron materia seca acumulada y su tasa de acumulación, área foliar, longitud del tallo y componentes del rendimiento del fríjol. Los datos se trataron con un análisis de varianza, comparación de medias y correlaciones simples. Los tipos de competencia redujeron significativamente la materia seca acumulada, las tasas de acumulación, el rendimiento y los componentes del rendimiento. Entre los tipos de competencia las diferencias no fueron significativas en los rendimientos en semilla y en materia seca acumulada en la planta. Si bien, hubo diferencias significativas entre los tratamientos durante su ontogenia, se establecieron procesos de retroalimentación dentro de cada tipo de competencia, que no permitieron diferenciar sus efectos en el último estadío de la planta, lo cual es una manifestación de que la planta de fríjol, es una unidad integrada a través de procesos fisiológicos.
Palabras clave: Cultivos asociados, fisiología de cultivos, ecofisiología, componentes de rendimiento.
Abstract. Associated corn and bean is one of several systems to produce climbing bean. The main characteristic of the associated is the greater yield by area than monoculture of two species, although the competence between them reduces the yield bean in 40% and 20% in corn. Research about physiological processes affected by competence between these species is still scarce. The objective of this research was to evaluate the effects of competence on the physiological behavior of bean. In a randomized block design with 4 replications were established the following treatments: bean without competition, competition between bean and maíze by light, competition between bean and maíze by soil resources and simultaneous competition between bean and maíze by both resources light and soil. The variables measured were accumulated dry matter and its accumulation rate, leaf area, stem longitude, and the yield components of bean. The variance analysis, comparison of means and simple correlations were used. The types of competition significantly reduced the accumulated dry matter, the rates of accumulation, the yield and the yield components. The types of competence did not show significant differences on seed weigh and accumulated total dry matter of plants. Although during the plants ontogeny there were significant differences between them, feedback processes were established within each type of competition. Therefore, the effects of competence in the final stage were not distinguished. It suggests that bean plant is an integrated unit through the physiological processes.
Key words: Intercropping, physiology of crops, ecophysiology, yield components.
Una forma de cultivar fríjol trepador es en asocio con maíz, la cual ha constituido un componente fundamental de la economía y viabilidad de los pequeños productores de las regiones andina y centroamericana. Las características de este arreglo productivo permiten al agricultor disminuir su vulnerabilidad a factores abióticos, bióticos, económicos y de mercado; mejorar su soberanía y seguridad alimentaria, ser altamente eficiente en el uso de recursos escasos como tierra, mano de obra y monetarios, obtener mayores rendimientos por unidad de tierra en comparación con los unicultivos de maíz o fríjol, contribuir a la protección y mejoramiento de la agrobiodiversidad in situ; disminuir la presión sobre el bosque por no requerir soporte para el fríjol, ya que el maíz cumple esta función; y disminuir la contaminación por agroquímicos debido a que permite reducir las aplicaciones producto del menor ataque de enfermedades, plagas y arvenses (Bazan et al., 1975; Altieri et al., 1977; Francis, 1978; Mead y Willey, 1980; Camarena y Cerrate, 1980; Sánchez, 1981; Beets, 1982; Tobón, 1990; Fukai, 1993; Trenbath, 1993; Cortez y Trujillo, 1994; Thrupp, 1998; Brush, 1999; Peter y Wanjau, 1999; Vélez, 1999; Morales y Manrique, 2000; Cannolly et al., 2001; Smale y Jarvis, 2002; Van Dusen, 2002; Birol, 2002; Broughton et al., 2003; Bavec et al., 2005; Abera et al., 2005; Songa et al., 2006; Gebeyehu et al., 2006; Kachigamba y Maulawo, 2007; Roldan et al., 2007; Worku, 2008; Yilmaz et al., 2008, Zeyaur et al., 2009, Santos et al., 2009; Mucheru-Muna et al., 2010).
Además, las condiciones de relieve y clima de las regiones andina y centroamericana, la cultura existente para su cultivo y las características de adaptabilidad de la planta de fríjol trepador, no han permitido el aprovechamiento suficiente del potencial del asocio fríjol - maíz. Las investigaciones que vienen realizando instituciones como el CIAT y CORPOICA en Colombia, INIAP en Ecuador, INIA y la Universidad Nacional Agraria La Molina en Perú, la Universidad Nacional de Colombia y la Universidad de Nariño en Colombia, hacen prever un importante potencial para su expansión en climas medios y cálidos en las regiones tropicales y subtropicales de Latinoamérica y África (CIAT, 1993; Checa y Blair, 2003).
El asocio fríjol - maíz consiste en la siembra de estas dos especies en el mismo sitio y al mismo tiempo. Su principal característica es la mayor producción por unidad de área a pesar de la competencia entre las dos especies que reduce los rendimientos del fríjol en más del 40% y del maíz hasta un 22%. La mayor producción por unidad de área índica mayor eficiencia en el uso de la tierra, en la captación y uso de la energía lumínica, del agua y nutrientes (Crookston et al., 1975; Sanabria, 1975; Allen et al., 1976; Henao y Mejía, 1981; Casadiego y García, 1982; Murcia y González, 1988; Firbank y Watkinson, 1990; Rajcan y Swanton, 2001; Tsubo et al., 2001), este es el principio de la producción competitiva (Vandermeer, 1989).
La competencia por recursos puede ser analizada como un tipo de interacción indirecta entre organismos, en que los recursos competidos constituyen el intermediario. El efecto inicial es de los competidores sobre la abundancia de recursos, y luego la respuesta de los competidores a los cambios en la abundancia de los recursos. Estos efectos permiten establecer dos mecanismos fisiológicos que posibilitan ser buenos competidores, uno es apropiándose y agotando rápidamente el recurso, es el caso del maíz, y el otro es continuar el crecimiento en niveles disminuidos de los recursos haciendo una conversión más eficiente de ellos, es el caso del fríjol, que aún así alcanza producciones económicas significativas. Estos mecanismos están relacionados con características fisiológicas y morfológicas de las especies, y con la posición, actividad y tamaño de los órganos que capturan recursos, lo cual está en función de la distribución de fotoasimilados. La mayor asignación de fotoasimilados a las hojas o al crecimiento del tallo, puede maximizar la interceptación de luz, mientras que si es a las raíces puede contribuir a mejorar la captura de recursos del suelo (Donald, 1963; Grime, 1977; Louwerse y Zweerde, 1977; Grime, 1979; Givnish, 1982; Tilman, 1982, 1985, 1987; Davis y García 1983a, 1983b; Wilson y Keddy, 1986; Chapín et al., 1987; Caldwell et al., 1987; Keddy y Shipley, 1989; Connell, 1990; Grace, 1990; Golberg, 1990; Keddy, 1990; Fukai y Trenbath, 1993; Trenbath, 1993; Keating y Carberry, 1993).
La mayor apropiación de recursos por parte del maíz se debe a características; a. Morfológicas, como su crecimiento erecto, posibilitado por la mayor asignación de fotoasimilados al tallo en comparación con el fríjol; b. De arquitectura, al concentrar sus hojas en los tercios medio y superior mientras que el fríjol las concentra en su tercio inferior y medio; y c. Fisiológicas, ya que el maíz es una poacea con mecanismo fotosintético C-4, en cambio el fríjol es una leguminosa C-3 tolerante a la sombra gracias a su plasticidad para modificar características como tamaño y grosor de hojas, concentración de clorofila y densidad de grana apilada. Estas características le permiten al fríjol mayor eficiencia fotosintética, disminuir su punto de compensación (Boardman, 1977; Tsubo et al., 2001; Tsubo y Walker, 2004), explotación de microambientes y crecer en niveles disminuidos de recursos (Golberg, 1990).
Las relaciones de competencia entre plantas, aún están poco entendidas, ya que muchos de los experimentos de campo se diseñan para demostrar la existencia de la competencia, más no para entender los mecanismos (Tilman, 1987). En otros casos, la competencia se asume como un postulado de causalidad. También se presentan situaciones en que no es fácil diferenciar entre los efectos de la competencia por diferentes recursos. Así en la competencia por luz, la planta sombreada podría reducir la disponibilidad de fotoasimilados para el crecimiento de la raíz, afectando negativamente la toma de nutrientes. Consecuentemente, lo que inicialmente era competencia por luz se traduce en competencia por nutrientes (Caldwell et al., 1987; Keddy, 1990). Esto demuestra, primero, que la parte aérea de la planta y las raíces son partes de una unidad integrada fisiológicamente que no deben ser tratadas como entidades independientes, y, segundo, que las interacciones entre recursos del suelo y de la superficie del suelo, son inevitables por lo que no es real particionar, por ejemplo, el nitrógeno y la radiación incidente (Harper, 1977; Vandermeer, 1989).
Otro aspecto que se debe considerar es el análisis integrado de los procesos que ocurren durante la ontogenia de las especies en competencia. Una adecuada interpretación de los resultados debe incluir el periodo de tiempo de evaluación, los días a inicio de captura de recursos, la eficiencia en la captura y uso de recursos en los diferentes estados de crecimiento (Newman, 1983). Evidencias experimentales en el cultivo asociado fríjol - maíz (Gardiner y Craker, 1981; Díaz et al., 1995,) indican que la competencia por los diferentes recursos es dinámica y diferencial durante la ontogenia de las especies, lo mismo que los mecanismos y respuestas de las plantas. En los primeros estados, la competencia es de poca significancia, pero es progresiva y cambia en cuanto al tipo de recursos por los que compiten y al tiempo de agotamiento de los mismos.
La investigación y manejo de cultivos asociados, particularmente de fríjol - maíz, se ha orientado a identificar y evaluar los efectos de la competencia sobre el rendimiento y sus componentes, y sobre factores y labores agronómicas que permiten cuantificar, atenuar y manejar dichos efectos, como poblaciones relativas de cada especie en el asocio, arreglos espaciales, épocas de siembra, fertilización, selección de genotipos apropiados y efectos sobre población de artrópodos. Por el contrario, pocas investigaciones han estudiado aspectos fisiológicos que expliquen la reducción en el rendimiento de las leguminosas en el asocio y los mecanismos de la competencia (Edje y Laing, 1982). Para Colombia Vélez et al. (2007) encontraron que de 21 investigaciones que evalúan algún aspecto fisiológico del cultivo asociado fríjol - maíz, solo dos evalúan características fisiológicas.
Resultados similares se hallaron en una revisión de estudios experimentales reportados entre 2004 a 2010 en las bases de datos Academia Search Complete, Annual Reviews, Science Direct, CAB Direct y EBSCO relacionados con el cultivo asociado fríjol trepador (Phaseolus vulgaris L.) -; maíz (Zea mays L.), incluyendo como criterio de búsqueda las palabras claves ''Intercropping'' ''Intercropping maíze and bean'' ''Growth curves'', ''Corn ecophysiology'' y ''Bean ecophysiology''. La búsqueda arrojó un total de 242.464 publicaciones, de las cuales se seleccionaron aquellas que cumplían con los siguientes criterios: arreglos cronológicos en siembra y cosecha escalonada y cultivos en asociación de acuerdo con la terminología de Hart (1984), que se tuviera acceso al artículo completo debido a la necesidad de identificar que el fríjol fuera trepador (Betancur et al., 2010). Solo 17 publicaciones, que representan el 0,007% de las publicaciones, cumplieron con estos criterios (Bavec et al., 2005; Maciel et al., 2004; Tsubo y Walker, 2004; Castillo y González, 2005; Ogindo y Walker, 2005; Songa et al., 2006; Gebeyehu et al., 2006, Oramas y Vivas, 2007, Roldán et al., 2007; Kachigamba y Maulawo, 2007; Worku, 2008; Yilmaz et al., 2008; Castillo y González, 2008, Zeyaur et al., 2009; Xinhua et al., 2009; Santos et al., 2009; Mucheru-Muna et al., 2010). De éstas, solamente tres estudiaron variables explicativas relacionadas con procesos fisiológicos como tasas fotosintéticas, materia seca, madurez fisiológica e índice de área foliar (IAF) (Bavec et al., 2005; Tsubo y Walker, 2004; Zeyaur et al., 2009). Estas investigaciones mencionan un mayor IAF en asocio maíz - fríjol (5,0) que en unicultivo de maíz (3,7) y una correlación positiva del mayor IAF del asocio con el rendimiento en grano por unidad de área (Bavec et al., 2005) y con una mayor eficiencia en el control de arvenses (Zeyaur, 2009). El tiempo de la emergencia, la floración y la madurez, determinados por el tiempo térmico (Grados día (DD): 8 °C d), estuvo fuertemente ligado a la disponibilidad de agua del suelo y la temperatura (Tsubo et al., 2004).
Estos resultados evidencian el estado incipiente del conocimiento sobre factores, mecanismos y procesos ecofisológicos determinantes del comportamiento y productividad del fríjol en el cultivo asociado, a pesar de tratarse de especies de importancia económica y de seguridad y soberanía alimentaria en Colombia, la región Andina, Centroamérica y recientemente países africanos. Un mejor conocimiento de estos aspectos, permitirá establecer relaciones de causa y avanzar en el mejoramiento de las estrategias de manejo del asocio.
Con base en los anteriores análisis, ésta investigación se planteó las siguientes preguntas: ¿las relaciones de competencia son diferenciales y dinámicas durante la ontogenia del cultivo asociado fríjol -; maíz? y ¿durante la competencia se establecen procesos de retroalimentación entre la explotación de los recursos del suelo, la explotación de recursos de la superficie del suelo y el crecimiento de las plantas?. El objetivo de ésta investigación fue evaluar los efectos de la competencia por luz, por recursos del suelo y de la competencia simultánea por luz y recursos del suelo, sobre cambios morfológicos y acumulación de materia seca, su dinámica y distribución durante la ontogenia de las plantas.
MATERIALES Y MÉTODOS
El experimento se realizó en los invernaderos de la Facultad de Agronomía de la Universidad Nacional de Colombia, Sede Bogotá (4º 38' 21,1'' latitud norte y 74º 05' 29,3'' longitud oeste), ubicado en la zona de vida bosque seco Montano Bajo (bh-MB) (Espinal, 1977), a una altitud de 2.547 msnm. En el año 2003 la temperatura media fue 13,7 ºC, la precipitación fue 800 mm, la humedad relativa fue 80% y el número de horas de brillo solar fueron 1.559, que corresponden con los promedios históricos para la zona (Instituto de Hidrología, Meteorología y Estudios Ambientales 2008). Se utilizaron cultivares regionales de maíz sogamoseño y de fríjol trepador bola roja de hábito de crecimiento indeterminado y alta capacidad trepadora, tipo IVb.
Se establecieron los siguientes cuatro tratamientos: fríjol sin competencia (Fo), fríjol y maíz en competencia simultánea por luz y recursos del suelo (FxM), fríjol y maíz en competencia sólo por luz (F*M), fríjol y maíz en competencia sólo por recursos del suelo (F_M). Cada tratamiento consistió de 24 bolsas plásticas. En Fo se utilizaron bolsas de 20 kg y se sembró una planta de fríjol por bolsa, en los tratamientos FxM, F*M y F_M se utilizaron bolsas de 40 kg y se sembró una planta de fríjol y una planta de maíz por bolsa. En Fo y F_M en cada bolsa se colocó como tutor para sostener el fríjol una vara de madera de dos metros de largo. En F_M, al tutor se le ató una tela plástica negra separando la planta de maíz y la del fríjol, para evitar la competencia por luz, durante el día la bolsa se rotaba de tal manera que ambas plantas recibieran igual cantidad de luz. En F*M, se colocó una tabla en medio de la bolsa de triplex inmunizada separando las raíces del fríjol y del maíz para evitar la competencia por recursos del suelo.
La fertilización se hizo según resultados de los análisis de suelo del Laboratorio de Suelos de la Facultad de Agronomía de la Universidad Nacional de Colombia, Sede Bogotá y en los requerimientos de la planta según las recomendaciones de la quinta aproximación (ICA, 1992). En el caso de los tratamientos sin competencia por recursos del suelo (Fo y F*M), se tuvieron en cuenta los requerimientos de cada planta. En el caso de los tratamientos con competencia por recursos del suelo (FxM y F_M), se asumieron los requerimientos del maíz, que es la especie con mayor demanda de nutrientes y es como lo hacen los agricultores. La humedad del suelo se mantuvo a capacidad de campo evaluada por el método propuesto por Laffite (1994).
El diseño experimental usado fue de bloques completos al azar, con tres repeticiones, tomando como unidad experimental la planta. Se realizaron muestreos cada 45 días de acuerdo con la escala de estadios de las plantas mono y dicotiledóneas (Meier, 2001). En cada muestreo se tomaron dos bolsas por tratamiento, el suelo se lavó con manguera provista de regadera para no dañar las raicillas, y se separaron las raíces. Las variables medidas en los tres primeros muestreos fueron materia seca de raíz, tallo y hojas, para lo cual se colocaron en bolsas de papel, se marcaron y secaron a 70 °C durante 72 h en una estufa WTC Binder® Fd 115. Posteriormente, se registró el peso seco de cada órgano de acuerdo con el estadio de desarrollo. Se midió longitud de raíz, tallo, y el diámetro del tallo. El área foliar se estimó con base en 30 foliolos para cada planta mediante la aplicación de la fórmula yt = (AF de 30 foliolos/30 foliolos)*número total de foliolos (Marín et al., 1998), donde yt es el área foliar (AF) de la planta. Los 30 foliolos son el producto de tomar 10 foliolos de cada tercio del perfil de la planta (tercio superior, medio e inferior), con base en la longitud de la guía principal del fríjol. En el último muestreo, se tomó el peso seco del total de granos y de un grano, y se contaron el número de vainas/planta y el número de granos/vaina.
Se realizó el análisis de varianza para cada muestreo, órgano e interacción. Para las variables con diferencias estadísticamente significativas se hizo comparación de medias y análisis de correlación simple. Los datos fueron procesados y analizados en los entornos estadísticos SAS versión 9.0 (Statistical Análisis System) y MINITAB, en el Departamento de Estadística de la Universidad Nacional de Colombia, Sede Bogotá.
RESULTADOS
Efectos de la competencia sobre la materia seca en la planta. Los tres tipos de competencia afectaron significativamente la materia seca acumulada en la planta durante la ontogenia del fríjol. La materia seca del fríjol sin competencia (Fo) fue significativamente mayor que la materia seca del fríjol compitiendo por luz (F*M) a los 90, 135 y 180 días después de siembra (dds), a la del fríjol compitiendo por recursos del suelo (F_M) a los 90 y 180 dds, y a la del fríjol compitiendo simultáneamente por luz y recursos del suelo (FxM) a los 180 dds (Tablas 1 y 2). El fríjol compitiendo por luz presentó la menor acumulación de materia seca durante el periodo evaluado. Entre los tipos de competencias, las diferencias fueron significativas solo a los 90 dds.
Tabla 1. Efectos en el crecimiento de los tipos de competencia del fríjol con maíz.
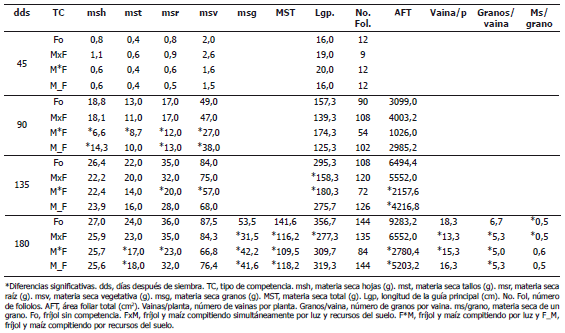
Tabla 2. Diferencias en la materia seca acumulada entre órganos de fríjol y maíz en los tipos de competencia.
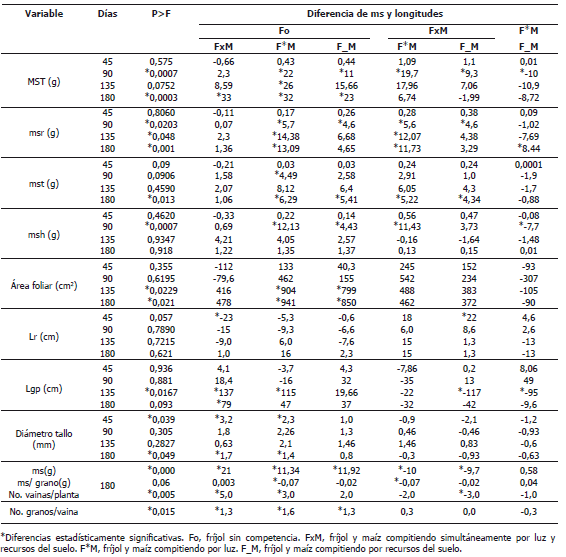
Efectos de la competencia sobre la materia seca en los diferentes órganos de las plantas. El Fo fue el que mayor materia seca acumuló en raíz, tallo y hojas durante toda su ontogenia, con diferencias significativas con el F*M a los 90 dds en todos los órganos, a los 135 dds en raíz y a los 180 dds en raíz y tallo. Con el F_M, las diferencias significativas fueron a los 90 dds en raíz y hojas y a los 180 dds en tallo. En los componentes del rendimiento (número de vainas por planta y número de granos por vaina) y en grano, Fo superó de manera significativa a todos los tipos de competencia (Tablas 1 y 2).
Se esperaba que la competencia simultánea por luz y recursos del suelo fuera la de mayor efecto en la acumulación de materia seca tanto en la parte vegetativa como en el rendimiento y en los componentes del rendimiento. Sin embargo, sus efectos sobre la parte vegetativa no fueron significativos con respecto al Fo, pero sí lo fue en el rendimiento y en los componentes del rendimiento, que fueron significativamente menores no sólo con respecto al Fo, sino también con relación a los otros tipos de competencia, especialmente con el F*M. Es decir, la planta de fríjol compitiendo simultáneamente por luz y recursos del suelo, dio prioridad al crecimiento de órganos captadores de recursos, caso contrario a lo sucedido con los otros tipos de competencia.
El fríjol compitiendo por luz acumuló la menor materia seca en todos sus órganos durante toda la ontogenia del fríjol, con diferencias significativas con el FxM en raíz a partir de los 90 dds, en tallo a los 180 y en hojas a los 90 dds. También presentó diferencias significativas con el F_M en hojas a los 90 dds y en raíz a los 180 dds. De acuerdo con estos resultados, los efectos de la competencia por luz, al compararlo con Fo y FxM, fueron principalmente sobre la raíz lo que podría limitar la toma de recursos del suelo y con ello la disponibilidad de recursos para la síntesis de rubisco y clorofila. Si se compara con F_M y FxM, el efecto principal de la competencia por luz, fue sobre la materia seca acumulada en las hojas, lo cual limitaría la fijación de CO2 y la producción de fotoasimilados, restringiendo el crecimiento de raíces.
El tallo fue el órgano con menor materia seca en todos los muestreos, lo cual se entiende si se considera que el fríjol voluble trepador no tiene un tallo vigoroso y requiere de tutor. El órgano que mayor materia seca acumuló fue el grano.
Efectos de la competencia sobre la dinámica de acumulación de materia seca. Las tasas de acumulación de materia seca y su distribución entre los diferentes órganos de la planta se presentan en la Tabla 3 y constituyen indicadores de la importancia relativa de cada órgano para la planta durante su ontogenia. En los órganos vegetativos, las mayores tasas se registraron entre los 90 y 135 dds, periodo que corresponde al crecimiento de vainas. El tallo fue el órgano con menores tasas y porcentajes de materia seca, durante todo el periodo evaluado en todos los tipos de competencia, excepto en el F*M a los 90 dds cuando superó a las hojas. En el caso del Fo, durante el periodo inicial de crecimiento hasta los 90 dds, las tasas de acumulación y porcentajes de materia seca asignados a las hojas y raíces fueron los mayores. A partir de los 135 dds, durante el llenado de vainas, las tasas y el porcentaje de materia seca asignada a las hojas, cayeron de manera considerable, en tanto que en la raíz continuaron altas. Es decir, el Fo dio prioridad, durante toda su ontogenia, a la raíz. Donde hubo competencia por recursos del suelo (F_M y FxM), en los primeros periodos de crecimiento, la prioridad fueron las hojas al presentar las mayores tasas y porcentajes de materia seca asignada. A partir de los 135 dds, la prioridad fueron las raíces. En el F*M, durante su periodo inicial de crecimiento la prioridad fueron las raíces y el tallo al presentar las mayores tasas y asignarles mayor porcentaje de materia seca. A partir de los 135 dds, la asignación de materia seca fue similar para hojas y raíces.
Tabla 3. Distribución y tasas de acumulación de materia seca en la planta de fríjol trepador en los diferentes tipos de competencia con maíz.
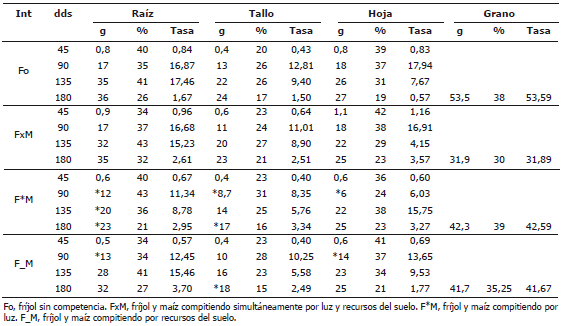
En todos los tipos de competencia, durante el periodo de llenado de vainas, 135 -; 180 dds, las tasas de acumulación de materia seca de los órganos vegetativos cayeron drásticamente, mientras que las tasas en grano fueron las mayores de todo el periodo evaluado en todos los tipos de competencia. Esto indica una mayor tasa fotosintética durante el llenado de grano, dando máxima prioridad en la asignación de fotoasimilados, al grano. El Fo presentó la mayor tasa de acumulación de materia seca en grano (53,6 g) y la menor tasa fue para el FxM (32 g). El F*M y el F_M presentaron tasas alrededor de 42 g. En cuanto a la distribución de la materia seca, el Fo y el F*M destinaron los mayores porcentajes de materia seca al grano, alrededor de 38%, y menos a los órganos vegetativos, 62%. El F_M destinó el 35% de la materia seca al grano. En el FxM, la situación fue diferente, ya que éste fue el que menor porcentaje de su materia seca destinó al grano (27,4%) y un mayor porcentaje a sus órganos vegetativos (más del 70%), y fue el único caso en que la planta destinó una proporción similar de materia seca a un órgano vegetativo, la raíz (32%), y al grano (Figura 1).
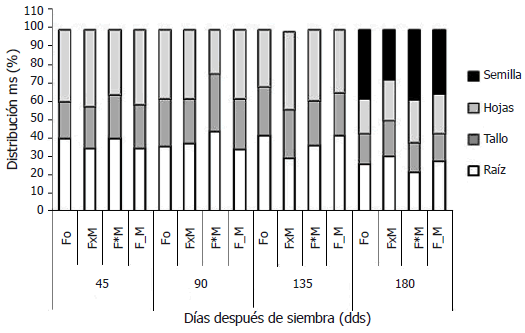
Figura 1. Distribución de la materia seca en la planta de fríjol (%). Fo, fríjol sin competencia. FxM, fríjol y maíz compitiendo simultáneamente por luz y recursos del suelo. F*M, fríjol y maíz compitiendo por luz. F_M, fríjol y maíz compitiendo por recursos del suelo.
Efectos de la competencia sobre el área foliar y la longitud de la guía principal. En los estados iniciales de la ontogenia, hasta los 90 dds, las diferencias en las áreas foliares del Fo y todos los tipos de competencia no fueron significativos, aunque la materia seca en hojas del F*M y del F_M fueron significativamente menores (Tablas 1 y 2). Durante el llenado de vainas (135 y 180 dds), las áreas foliares del de FxM, F*M y de F_M fueron significativamente menores al área foliar de Fo, aunque en materia seca las diferencias fueron significativas solo entre el F*M y el Fo. La menor área foliar se traduce en menor área de captación de luz solar, mientras que hojas más delgadas posiblemente afecta la fijación de CO2 y se traduce en menor contenido de proteínas, entre ellas rubisco. El fríjol compitiendo por luz presentó, durante toda la ontogenia, las menores áreas foliares y las hojas más delgadas.
En el periodo de crecimiento de vainas, 90-135 dds, la longitud de la guía principal fue significativamente menor en las plantas de fríjol en las cuales la competencia por luz estuvo presente (F*M y FxM). En cambio, las diferencias en materia seca no fueron significativas. Estas guías más cortas limitan el acceso de la planta a la luz. El F*M, hasta los 90 dds, fue el de mayor tasa de crecimiento en la longitud de la guía principal (154,3 cm), a los 135 dds prácticamente no creció, su tasa de crecimiento fue de 6 cm. El FxM, cuyo crecimiento a los 90 dds (120,3 cm) superaba la del F_M, y presentó una tasa muy baja de 19,07 cm con respecto al Fo y al F_M, cuyas tasas sobrepasaron los 130 cm (Tablas 1 y 2). A los 180 dds, las plantas de fríjol compitiendo por luz (F*M y FxM) nuevamente fueron las que presentaron las mayores tasas de crecimiento, 119 y 130 cm respectivamente.
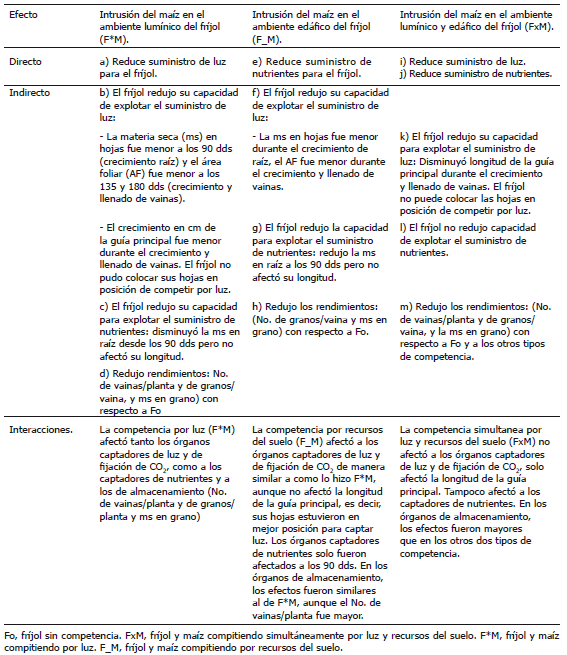
En la Tabla 4 se presentan los efectos de la competencia por los diferentes recursos sobre el aprovechamiento de otro recurso que no estaba en competencia.
Tabla 4. Interacciones entre la competencia por recursos del suelo y la competencia por recursos de la superficie del suelo.
DISCUSIÓN
El fríjol compitiendo por luz presentó la menor acumulación de materia seca durante el periodo evaluado, 180 dds, sin embargo, las diferencias fueron significativas solo con el fríjol sin competencia. La competencia por luz afectó significativamente la materia seca en órganos, especialmente en raíz y en menor proporción en tallo, no en hojas, tampoco en la longitud de la raíz ni de la guía principal. La competencia por luz también afectó significativamente el área foliar, fue la que menos afectó la acumulación de materia seca en el grano y fue donde el mayor porcentaje de materia seca, se destinó al grano, superando inclusive al fríjol sin competencia.
Los efectos de la competencia por recursos del suelo fueron similares a los efectos de la competencia por luz, excepto en la raíz, cuyas diferencias en la ms acumulada no fueron significativas con Fo ni con FxM. Tampoco en el número de vainas/planta mostraron diferencias significativas.
La competencia simultánea por luz y recursos del suelo, no afectó la acumulación de materia seca en los órganos vegetativos ni la distribución de los fotoasimilados entre ellos, ni el área foliar, ni la longitud de la raíz. Es decir, los órganos captadores de recursos, hojas y raíz, estuvieron en condiciones similares a las del fríjol sin competencia. Sin embargo, esta competencia simultánea fue la de mayor efecto negativo en la acumulación de materia seca en grano y en los componentes del rendimiento, con diferencias significativas respecto a los otros tipos de competencia. Esto podría indicar que el fríjol compitiendo simultáneamente por luz y recursos del suelo dio prioridad a los órganos vegetativos. La respuesta del fríjol compitiendo por luz y del fríjol compitiendo por recursos del suelo fue diferente: las tasas y porcentajes de asignación de materia seca a grano y a vainas llegaron a superar a las del fríjol sin competencia. En este caso, la prioridad en la distribución de fotoasimilados fue para los componentes del rendimiento. No obstante, el tamaño de sus órganos captadores de recursos, medido en materia seca en raíz y hojas y en área foliar, fue significativamente menor con respecto a los otros tratamientos.
En general, en todos los tratamientos se dio prioridad a la acumulación de materia seca en raíz, excepto a los 45 dds en el fríjol compitiendo por recursos del suelo y en el fríjol compitiendo simultáneamente por luz y recursos del suelo.
Los resultados muestran un efecto similar de la competencia por luz y de la competencia por recursos del suelo y pueden indicar que ambos efectos son de igual importancia, lo cual constituye una diferencia importante con la mayoría de las publicaciones que identifican la competencia por luz, como la principal causa de reducción de materia seca en el fríjol cuando se asocia con maíz. Varios autores han centrado sus investigaciones en esta perspectiva, Así, por ejemplo, argumentan que la altura es una de las características que más confiere habilidad competitiva al permitir a las plantas mejor acceso a la luz (Davis y García, 1983a; Angulo, 1986; Caldwell et al., 1987; Keddy y Shipley, 1989; Keddy, 1990; Kohashi, 1990; Tsubo y Walker, 2004). Vélez et al. (2007), muestran que alrededor del 52% de las investigaciones realizadas en Colombia han sido sobre altura y longitud de entrenudos de las plantas de maíz y fríjol en asocio. Marín et al. (1998), sustentan el efecto determinante de la competencia por luz, por medio de los valores que obtuvo para el potencial hídrico foliar, los cuales fueron más bajos en el asocio, lo que sugiere mayor extracción de agua en correspondencia con el mayor índice de área foliar del asocio. Solo Sanabria (1975), encontró que el fríjol en asocio con maíz (FxM), en hidroponía, produjo tanta materia seca como el fríjol sin competencia. Sin embargo, cuando el asocio se sembró en suelo los resultados obtenidos fueron similares a los reportados por la literatura. Con estos resultados se concluye que los efectos depresivos de la asociación sobre el fríjol, se deben a la competencia por recursos del suelo y no a la competencia por luz.
Los resultados de esta investigación apoyan la hipótesis sobre la existencia de procesos de retroalimentación entre la explotación de los recursos del suelo, la explotación de recursos de la superficie del suelo y el crecimiento de las plantas (Tabla 4). El hecho de que cambios en el comportamiento de las hojas (materia seca y área foliar), tengan efectos sobre el sistema de raíces y el comportamiento de las raíces a su vez, incida sobre el desarrollo de la parte aérea, hace que las interacciones entre los recursos del suelo y los recursos de la superficie del suelo, sean inevitables y debe considerarse la planta como una unidad integrada fisiológicamente (Harper, 1977; Vandermeer, 1989).
CONCLUSIONES
Los resultados obtenidos muestran que las relaciones de competencia son diferenciales y dinámicas durante la ontogenia del asocio y que entre las posibles respuestas a la competencia están los procesos de retroalimentación (Tabla 4) y la participación de asimilados, en este caso, entre la parte vegetativa y el rendimiento y sus componentes.
Aunque durante la ontogenia del asocio los efectos de la competencia fueron diferentes, al final del ciclo el efecto total fue similar, al igual que los efectos de los diferentes tipos de competencia. Esta es una manifestación de que la planta de fríjol, es una unidad que responde como un todo debido a procesos fisiológicos. Es decir, aquello que inicialmente fue una competencia por un recurso del suelo, en etapas posteriores puede manifestarse como resultado de una competencia por luz. Por tanto, el conocimiento de la dinámica de la competencia, es importante para establecer los recursos por los cuales se compite en un momento dado y las respuestas específicas de las plantas.
Durante el periodo de crecimiento inicial, hasta los 45 dds, los tipos de competencia no tuvieron efectos significativos en la materia seca acumulada en los órganos, ni en el área foliar. Los efectos fueron significativos solamente donde hubo competencia por luz (F*M y FxM), afectando la longitud de la raíz. En este periodo, las raíces y las hojas aún no se sobreponen en la demanda por asimilados porque dependen, en gran medida, de las reservas nutricionales de la semilla.
Estos resultados constituyen un referente para la investigación y el diseño de estrategias de manejo, dirigidas, de manera especifica, a las diferentes etapas de crecimiento y desarrollo de los cultivos asociados, de acuerdo con los recursos por los cuales se esté compitiendo y de acuerdo con las características morfológicas, fisiológicas y bioquímicas de las plantas. De igual manera, las investigaciones en fisiología y mejoramiento genético, deben diseñarse de acuerdo con estos criterios para tratar de aliviar las causas y mejorar las respuestas de la plantas.
AGRADECIMIENTOS
Esta investigación fue posible gracias a la financiación del proyecto 20301003485 otorgada por la División de Investigación de la Universidad Nacional de Colombia, Sede Bogotá.
BIBLIOGRAFÍA
Abera, T., T. Tana and L. Pant. 2005. Grain yield and LER maize-climbing beam intercropping affected by inorganic, organic fertilizers and population density in western Oromiya, Etiopia. Asian Journal of Plant Science 4(5): 458-465. [ Links ]
Allen, L., T. Sinclair and E. Lemon. 1976. Radiation and microclimate relationship in multiple cropping systems. pp. 171-200. In: Matthias, S. (ed.). Multiple cropping. American Society of Agronomy, Wisconsin, United States. 278 p. [ Links ]
Altieri, M., A. Schoonhoven and J. Doll. 1977. The ecological role of weeds in insect pest management systems: a review illustrated by bean (Phaseolus vulgaris L.) cropping systems. Tropical Pest Management 23(2): 195-205. [ Links ]
Angulo, N. 1986. Comportamiento agronómico de selecciones avanzadas de fríjol voluble con maíces en asocio y en unicultivo. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad Nacional de Colombia, Bogotá. 89 p. [ Links ]
Bavec, F., U. Živec, S. Mlakar and L. Radics. 2005. Competitive ability of maize in mixture with climbing bean in organic farming, http://orgprints.org/4214/4/4214-Bavec_etal_4p_revised-ed.pdf. 4 p.; consulta: febrero 2009. [ Links ]
Bazan, R., A. Pinchinat, G. Páez, N. Mateo, R. Moreno, J. Fargas and W. Forsythe. 1975. Investigación sobre sistemas de producción agrícola para el pequeño agricultor del trópico. Turrialba 25(3): 283-293. [ Links ]
Beets, W. 1982. Multiple cropping and tropical farming systems. Westview Press. Colorado, USA. 156 p. [ Links ]
Betancur, M., M. Mejía y L. Vélez. 2010. Estudio de la ecofisiología del cultivo asociado. Informe del Programa Especial de Trabajo Académico. Facultad de Ciencias Agropecuarias, Universidad Nacional de Colombia, Sede Medellín. 21 p. [ Links ]
Birol, E. 2002. Applying environmental valuation methods to support the conservation of agricultural biodiversity. pp. 19-29. In: Smale, M., I. Már and D.I. Jarvis. (eds.). The economics of conserving agricultural biodiversity on farm. International Plant Genetic Resources Institute (IPGRI), Rome, Italy. 73 p. [ Links ]
Boardman, N. 1977. Comparative photosynthesis of sun and shade plants. Annual Review of Plant Physiology 28: 355-377. [ Links ]
Broughton J., G. Hernández, M. Blair, S. Beebe, P. Gepts and J. Vanderleyden. 2003. Beans (Phaseolus spp.)- model food legumes. Plant and Soil 252(1): 55-128. [ Links ]
Brush, S.B. 1999. Genes in the field: on farm conservation of crop diversity. International Plant Genetic Resources institute (IPGRI), Rome, Italy. 288 p. [ Links ]
Caldwell M., J. Richards, J. Manwaring and D. Eissenstat. 1987. Rapid shift in phosphate acquisition show direct competition between neighboring plants. Nature 327: 615-616. [ Links ]
Camarena, P y A. Cerrate. 1980. Comparación de sistemas en monocultivo y asociado y época oportuna de siembra de fríjol en asociación con maíz en El Callejón de Huaylas. Universidad Nacional Agraria La Molina. Anales Científicos 18(1-4): 191-197. [ Links ]
Cannolly, J., H. Goma and K. Rahim. 2001. The information content of indicators in intercropping research. Agriculture, Ecosystems and Environment 87(2): 191-207. [ Links ]
Casadiego, E. y I. García. 1982. Evaluación agronómica de veinticinco variedades de maíz (Zea mays L.) de clima frio bajo el sistema de asociación con fríjol voluble. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad Nacional de Colombia, Bogotá. 164 p. [ Links ]
Castillo, N. y C. González. 2005. Efecto del color y la altura de las trampas sobre la captura de cicadélidos en la asociación fríjol-maíz. Revista Protección Vegetal 20(2): 128-131. [ Links ]
Castillo, N. y C. González. 2008. Comportamiento poblacional de insectos fitófagos en el unicultivo de fríjol (Phaseolus vulgaris L.) y en la asociación con maíz (Zea mays L.). Revista Protección Vegetal 23(3): 154-159. [ Links ]
Chapin, III F., A. Bloom, C. Field and R. Waring. 1987. Plant responses to multiple environmental factors. BioScience 37(1): 49-57. [ Links ]
Checa, O. and M. Blair. 2003. Trait correlations in climbing beans. Annual Report of the Bean Improvement Cooperative (BIC) 46(46): 15-16. [ Links ]
CIAT. 1993a. Adaptation of Andean climbing beans to warm temperatures. In: Bean Program, annual report. Working document 161: 85-87. [ Links ]
Connell, J. 1990. Apparent versus real competition in plants. In: Grace, J.B. y D. Tilman (eds). Perspective on plant competition. Academic Press, San Diego, California. 484 p. [ Links ]
Cortez, M. y A. Trujillo. 1994. Incidencia del gusano cogollero y sus enemigos naturales en tres agrosistemas de maíz. Turrialba 44(1): 1-9. [ Links ]
Crookston, R., K. Treharne, P. Ludford and J. Ozbun. 1975. Response of bean to shading. Crop Science 15(3): 412-416. [ Links ]
Davis, J. and S. García. 1983b. Competitive ability and growth habit of indeterminate beans and maize for intercropping. Field Crops Research 6: 59-75. [ Links ]
Davis, J. y S. García. 1983a. Interacción de genotipos por sistema de cultivo en fríjol - maíz. En: CIAT, Programa de Fríjol. Curso de Fríjol. Cali, Colombia. 8 p. [ Links ]
Díaz, C., J. Rivera y J. Quirós. 1995. Crecimiento y desarrollo del maíz y fríjol trepador en monocultivo y asociación. Actualidades CORPOICA 9(9): 5-10. [ Links ]
Donald, C. 1963. Competition among crop and pasture plants. pp. 1-118. In: Norman, A.G. (ed.). Advances in Agronomy, volume 15. Academic Press, New York. 409 p. [ Links ]
Edje, O. and D. Laing. 1982. Physiological aspects of maize and bean in monoculture and in association summary. p. 69. In: Keswani, C.L. and B.J. Ndunguru (eds.). Intercropping: Proceedings of the Second Symposium on Intercropping in Semi Arid Areas, held at Morogoro, Tanzania. International Development Research Centre, Otawa. 168 p. [ Links ]
Espinal, S. 1977. Zonas de vida o formaciones vegetales de Colombia: Memoria explicativa sobre el mapa ecológico. Instituto Geográfico Agustín Codazzi, Bogotá. 238 p. [ Links ]
Firbank, L. and A. Watkinson. 1990. On the effects of competition: from monocultures to mixture. In: Grace, J.B. and D. Tilman (eds.). Perspective on plant competition. Academic Press, San Diego, California. 484 p. [ Links ]
Francis, C. 1978. Multiple cropping potentials of beans and maize. HortScience 13(1): 12-17. [ Links ]
Fukai, S. 1993. Intercropping, basis of productivity. Field Crop Research 34(3,4): 239-245. [ Links ]
Fukai, S. and B. Trenbath. 1993. Processes determining intercrop productivity and yields of components crops. Fields Crops Research 34(3,4): 247-271. [ Links ]
Gardiner, T. and L. Craker. 1981. Bean growth and light interception in a bean-maize intercrop. Field Crops Research 4: 313-320. [ Links ]
Gebeyehu, S., B. Simane and R. Kirkby. 2006. Genotype x cropping system interaction in climbing beans (Phaseolus vulgaris L.) grown as sole crop and in association with maize (Zea mays L.). European Journal of Agronomy 24(4): 396-403. [ Links ]
Givnish, T. 1982. On the adaptive significance of leaf height in forest herbs. The American Naturalist 120(3): 353-381. [ Links ]
Golberg, D. 1990. Components of resource competition in plant communities. In: Grace, J.B. and D. Tilman. (eds.). Perspective on plant competition. Academic Press, San Diego, California. 484 p. [ Links ]
Grace, J. 1990. On the relationship plant traits and competitive ability. In: Grace, J.B. and D. Tilman. (eds.). Perspective on plant competition. Academic Press, San Diego, California. 484 p. [ Links ]
Grime, J. 1977. Evidence for the three primary strategies in plants and its relevance ecological and evolution theory. The American Naturalist 111(982): 1169-1194. [ Links ]
Grime, J. 1979. Plant strategies and vegetation processes. Jhon Wiley, New York. 222 p. [ Links ]
Harper, J. 1977. Population biology of plants. Academic Press, New York. 892 p. [ Links ]
Hart, R. 1984. Agroecosistemas, conceptos básicos. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), Turrialba, Costa Rica. 211 p. (Serie Materiales de Enseñanza No. 1). [ Links ]
Henao, G. y F. Mejía. 1981. Épocas críticas de competencia entre las malezas y el cultivo de fríjol (Phaseolus vulgaris L.) en la zona de Santagueda Caldas. Tesis de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad de Caldas. 127 p. [ Links ]
ICA. 1992. Fertilización en diversos cultivos, quinta aproximación. FENALCE -; SENA -; SAC. Manual de Asistencia Técnica 25. 64 p. [ Links ]
Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM). Registros mensuales de parámetros climáticos. Estación 2120579, Fecha de proceso: julio 19 de 2008. [ Links ]
Kachigamba, D.L and A.C. Maulawo. 2007. Bean agronomy program, www.cabi.org.http://www.cabi.org/gara/FullTextPDF/2008/20083327150.pdf. 10 p.; consulta: noviembre 2009. [ Links ]
Keating, B. and P. Carberry. 1993. Resource capture and use intercropping: solar radiation. Field Crop Research 34(3,4): 273-301. [ Links ]
Keddy, P. 1990. Competitive hierarchies and centrifugal organization in plant communities. pp. 265-290 In: Grace, J.B. and D. Tilman. (eds.). Perspective on plant competition. The Blackburn Press, San Diego, California. 484 p. [ Links ]
Keddy, P. and B. Shipley. 1989. Competitive hierarchies in herbaceous plant communities. Oikos 54(2): 234-241. [ Links ]
Kohashi, J. 1990. Aspectos de la morfología y fisiología del fríjol (Phaseolus vulgaris L.) y su relación con el rendimiento. Centro de Botánica. Colegio de Postgraduados. Montecillo, México. 44 p. [ Links ]
Lafitte, H. 1994. Identifying production problems in tropical maize: a field guide. CIMMYT, Mexico D.F. 121 p. [ Links ]
Louwerse, W. and V. Zweerde. 1977. Photosynthesis, transpiration and leaf morphology of Phaseolus vulgaris and Zea mays grown at different irradiances in artificial and sunlight. Photosynthetica 11: 11-21. [ Links ]
Maciel, A.D., O. Arf, M.G. da Silva, M.E. de Sá, S. Buzetti, J.A. Andrade and E.S. Bianchini. 2004 Corn performance in intercropping corn-common bean in no tillage system. Acta Scientiarum Agronomy 26(3): 309-314. [ Links ]
Marín, D., A. Olivar y R. Cavanerio. 1998. Crecimiento y rendimiento en granos en una asociación de maíz (Zea mays L.) y caraota (Phaseolus vulgaris L.), con siembra simultánea. Revista de la Facultad de Agronomía 15(4): 297-311. [ Links ]
Mead, R. and R. Willey. 1980. The concept of a ''Land Equivalent Ratio'' and advantages in yield from intercropping. Experimental Agriculture 16(3): 217-228. [ Links ]
Meier, U. 2001. Estadios de las plantas mono y dicotiledóneas. Segunda edición. Centro Federal de Investigaciones Biológicas para Agricultura y Silvicultura, Berlín. 149 p. [ Links ]
Morales, C. y A. Manrique. 2000. Agrobiodiversidad en la región andina y amazónica. Asociación Gráficas Educativas, Lima. 416 p. [ Links ]
Mucheru-Muna, M., P. Pypers, D. Mugendi, J. Kung'u, J. Mugwe, R. Merckx and B. Vanlauwe. 2010. A staggered maize-;legume intercrop arrangement robustly increases crop yields and economic returns in the highlands of Central Kenya. Field Crops Research 115(2): 132-139. [ Links ]
Murcia, S. y J. González. 1988. Evaluación en Semijaca, Cundinamarca, de seis genotipos de maíz - fríjol voluble en diferentes densidades de población. Trabajo de Grado Ingeniero Agrónomo. Facultad de Agronomía. Universidad Nacional de Colombia, Bogotá. 155 p. [ Links ]
Newman, H. 1983. Interactions between plants. pp. 679-711. In: Lange, O.L., P.S. Nobel, C.B. Osmond and H. Ziegler (eds.). Physiological plant ecology III. Responses to the chemical and biological environment. Encyclopedia of Plant Physiology New Series. Spring-Verlag, New York. 799 p. [ Links ]
Ogindo H.O. and S. Walker. 2005. Comparison of measured changes in seasonal soil water content by rainfed maize-bean intercrop and component cropping systems in a semi-arid region of southern Africa. Physics and Chemistry of the Earth 30(11,16): 799-808. [ Links ]
Oramas, W. y N.J. Vivas. 2007. Evaluación de dos híbridos y una variedad de maíz (Zea mays) en monocultivo y en asociación con fríjol (Phaseolus vulgaris), para ensilaje. Facultad de Ciencias Agropecuarias 5(1): 28-35. [ Links ]
Peter, A. and F. Wanjau. 1999. Competition and productivity in crop mixture: some properties of productive intercrops. Journal of Agriculture Science 132(4): 425-435. [ Links ]
Rajcan, I. and C. Swanton. 2001. Uderstanding maize-weed competition: resource competition, light quality and the whole plant. Field Crop Research 71(2): 139-150. [ Links ]
Roldán, A., J.R. Salinas, M. Alguacil and M.F. Caravaca. 2007. Soil sustainability indicators following conservation tillage practices under subtropical maize and bean crops. Soil Tillage Research 93(2): 273-282. [ Links ]
Sanabria, E. 1975. Producción de biomasa, nutrición mineral y absorción de agua en la asociación maíz -; fríjol cultivada en solución nutritiva. Tesis Magister en Ciencia. Departamento de Cultivos y Suelos Tropicales. Instituto Interamericano de Ciencias Agrícolas de la OEA, Centro Tropical de Enseñanza e Investigación. Turrialba, Costa Rica. 80 p. [ Links ]
Sánchez, P. 1981. Suelos del trópico. Características y manejo. IICA, San José, Costa Rica. 634 p. [ Links ]
Santos, N., M. Tarsitano, O. Arf and G. Mateus. 2009. Economic analysis of maize (for green corn production) and common bean intercropping. Revista Brasileira de Milho e Sorgo 8(1): 1-12. [ Links ]
Smale, M. and D. Jarvis. 2002. The economics of conserving agricultural biodiversity on farm. Definitions and context. pp. 1-3. In: Smale, M., I. Már and D.I. Jarvis. (eds.). The economics of conserving agricultural biodiversity on farm. International Plant Genetic Resources Institute (IPGRI), Rome, Italy. 73 p. [ Links ]
Songa J.M., N. Jiang, F. Schulthess and C. Omwega. 2007. The role of intercropping different cereal species in controlling lepidopteran stemborers on maize in Kenya. Journal Applied Entomology 131(1): 40-49. [ Links ]
Thrupp, L. 1998. Cultivating diversity. agrobiodiversity and food security. World Resources Institute, Washington D.C. 80 p. [ Links ]
Tilman, D. 1987. The importance of the mechanisms of interspecific competition. The American Naturalist 129(5): 769-774. [ Links ]
Tilman, D. 1988. Plant strategies and the dynamics and structure of plant communities. Princeton University Press, New Jersey, USA. 376 p. [ Links ]
Tilman, D. 1982. Resource competition and community structure. Monograph in Population Biology 17. Princeton University Press, New Jersey, USA. 296 p. [ Links ]
Tilman, D. 1985. The resource-ratio hypothesis of plant succession. The American Naturalist 125(6): 827-852. [ Links ]
Tobón, J. 1990. La asociación de leguminosas en Colombia. En: Curso Nacional de Fríjol. Instituto Colombiano Agropecuario (ICA). C.I. La Selva. Rionegro, Antioquia. 12 p. [ Links ]
Trenbath, B. 1993. Intercropping for the management of pests and diseases. Field Crop Research 34(3-4): 381-405. [ Links ]
Tsubo, M. and S. Walker. 2004. Shade effects on Phaseolus vulgaris L. intercropped with Zea mays L. under well-watered conditions. Journal of Agronomy and Crop Science 190(3): 168-176. [ Links ]
Tsubo, M., S. Walker and E. Mukhala. 2001. Comparisons of radiation use efficiency of mono-/inter-cropping systems with different row orientations. Field Crops Research 71(1): 17-29. [ Links ]
Van Dusen, E. 2002. Identifying the determinants of crops biodiversity on farm with econometric applications of the household model. pp. 11-18. In: Smale, M., I. Már and D.I. Jarvis, (eds.). The economics of conserving agricultural biodiversity on farm. International Plant Genetic Resources Institute (IPGRI), Rome, Italy. 73 p. [ Links ]
Vandermeer, J. 1989. The ecology of intercropping. Cambridge University Press, Cambridge. U.K. 237 p. [ Links ]
Vélez, L. 1999. Dinámica y funcionalidad del cultivo de fríjol (Phaseolus vulagaris L.) a escala predial. p. 1-6. En: Memorias. Seminario Desarrollo Tecnológico para el Cultivo de Fríjol. Departamento de Ciencias Agronómicas, Universidad Nacional de Colombia, El Peñol, Antioquia. [ Links ]
Vélez, L., J. Clavijo y G. Ligarreto. 2007. Análisis ecofisiológico del cultivo asociado maíz (Zea mays L.) -; fríjol voluble (Phaselus vulgaris L.). Revista Facultad Nacional de Agronomía Medellín 60(2):3965- 3984. [ Links ]
Wilson, S. and P. Keddy. 1986. Measuring diffuse competition along an environmental gradient: results from a shoreline plant communities. The American Naturalist 127(6): 862-869. [ Links ]
Worku, W. 2008. Evaluation of common bean (Phaseolus vulgaris L.) genotypes of diverse growth habit under sole and intercropping with maize (Zea mays L.) in southern Ethiopia. Journal of Agronomy 7(4): 306-313. [ Links ]
Xinhua, H., X. Minggang, Y.Q. Guo, Z. Jianbin. 2009 Use of 15N stable isotope to quantify nitrogen transfer between mycorrhizal plants. Journal of Plant Ecology 2(3): 107-118. [ Links ]
Yilmaz, S., M. Atak and M. Erayman. 2008. Identification of advantages of maize-legume intercropping over solitary cropping through competition indices in the east mediterranean region. Turkish Journal of Agriculture and Forestry 32(2): 111-119. [ Links ]
Zeyaur, K., C. Midega, J. Wanyama, D. Amudavi, A. Hassanali, J. Pittchar and J. Pickett. 2009. Integration of edible beans (Phaseolus vulgaris L.) into the push-;pull technology developed for stemborer and Striga control in maize-based cropping systems. Crop Protection 28(11): 997-1006. [ Links ]

